斷尾對蘭州大尾羊生長性能、脂肪沉積分布和屠宰性能的影響
2023-02-27 14:27:00宋淑珍劉俊斌朱才業徐紅偉劉立山孔艷龍
畜牧獸醫學報 2023年2期
宋淑珍,劉俊斌,朱才業,徐紅偉,劉立山,孔艷龍
(1.甘肅省農業科學院畜草與綠色農業研究所,蘭州 730070; 2.甘肅農業大學動物科學技術學院,蘭州 730070;3.西北民族大學生命科學與工程學院,蘭州 730030;4.永靖縣瑞霖科技養殖有限公司,永靖 731603)
綿羊是最早被馴化家養的畜種之一[1],為人們提供肉、乳、毛、皮等重要的食物和御寒物品,在人類歷史進程中具有重要的經濟價值。世界上700多個綿羊品種中,將近25%的綿羊屬于脂尾型羊[2]。脂尾是綿羊在長期的選擇和進化過程中形成的特征,在早期,綿羊尾部脂肪不僅是人們重要的脂類來源食物,也是綿羊抵御極端寒冷氣候和營養匱乏等極端生存壓力的能量保證,綿羊在夏秋季牧草豐盛時儲存大量脂肪供冬春季牧草匱乏時動用以維持機體能量需要[3]。但隨著養殖業現代化的推進和人們生活水平的提高,大量脂肪沉積在尾部不僅浪費飼料,降低飼料效率,而且還會降低胴體重量,影響胴體品質[4];此外,人們日常飲食中過量的脂肪攝入會引起一些健康問題,因而對于生產來說綿羊尾部脂肪已經失去了其原來的價值,如何減少尾部脂肪的沉積,提高胴體品質是現代畜牧業的主要目標之一。
蘭州大尾羊屬于長脂尾型綿羊,具有耐粗飼、生長發育快、肉質鮮嫩等優點,其脂尾肥大,方圓平展,自然下垂至飛節上下,被尾中溝平分為左右對稱的兩瓣,尾尖外翻,成年羯羊尾脂占胴體重的13.23%[5],大量的脂肪沉積在尾部造成飼料資源的極大浪費,沉積1 kg脂肪所耗飼草料相當于生產2 kg瘦肉所需的飼草料[3],過多的尾部脂肪沉積降低了養羊業的經濟效益。此外,脂肪沉積與分布是影響胴體品質的主要因素[6],脂肪沉積在肌間有助于提高肉品質,而沉積在尾部、皮下、腹部等部位則成為“無用脂肪”[7-8],降低胴體質量。對1~3日齡的灘羊羔羊斷尾,135日齡左右屠宰時,尾部脂肪顯著降低,肌肉剪切力具有減小的趨勢,每天增加收益0.36元·只-1[9]。斷尾洼地綿羊公羔90日齡屠宰時,尾脂較未斷尾組減少了61.22%[10]。目前,保護畜禽種質資源,挖掘特有畜種生產潛力成為畜牧業的研究重點,如何減少脂尾型綿羊尾部脂肪沉積,并改善胴體脂肪沉積分布,使更多的脂肪沉積在肌間具有重要的研究意義。對于脂尾型綿羊減少尾脂沉積的研究主要集中在雜交改良[11]、早期屠宰[10]等方面,對改變脂肪沉積分布以及脂肪因子等涉及內分泌方面的報道較少。本研究探討斷尾對蘭州大尾羊生長性能、血清脂肪因子、屠宰性能和脂肪沉積分布等的影響,為通過脂肪沉積部位干預提高綿羊生產效率,改善胴體品質提供數據支持,并為脂尾型綿羊的生產利用提供參考。
1 材料與方法
1.1 試驗時間與地點
試驗于2021年3月24日—11月28日在永靖縣瑞霖養殖有限公司進行,年平均氣溫9.0 ℃,年平均日照時數2 517.6 h,海拔約2 150.0 m。
1.2 試驗動物與試驗設計
選擇血緣清楚、體重接近的5日齡健康蘭州大尾羊羔羊18只,隨機分為對照組和試驗組,每組9只羔羊,試驗組羔羊采用橡皮圈結扎法斷尾。試驗期兩組羊日糧相同,均飼喂全價配合飼料(精粗比為70∶30),自由飲水。每日上、下午各飼喂1次,每隔30 d對各組試驗羊只空腹靜脈采血、稱重。試驗期240 d,結束時,每組選擇4只羊屠宰取樣用于后續試驗。
1.3 試驗動物飼養管理
試驗羔羊隨母羊哺乳至2月齡斷奶,哺乳期自由哺乳,并通過羔羊補飼槽補飼羔羊補飼料(臨洮德華飼料有限公司提供),主要成分為玉米、豆粕、棉籽粕、植物油、奶粉、乳清粉、糖蜜、氨基酸、礦物質元素和微量元素等,營養成分為粗蛋白質≥18.0%,粗纖維≤7.0%,粗灰分≤8.0%,鈣0.60%~1.20%,總磷≥0.50%,氯化鈉0.30%~1.50%,賴氨酸≥0.80%,并添加6%的苜蓿和6%的燕麥草。羔羊斷奶后,轉入試驗羊舍;轉入前1周,對羊舍、羊欄、食槽等進行打掃、消毒。單欄飼養,羊舍溫度、光照、通風等飼養環境條件一致。每天上午7:00、下午16:00分2次飼喂,自由飲水。試驗日糧根據美國 NRC(2007)肉用綿羊營養需要配制,試驗期日糧的組成、營養水平見表1。
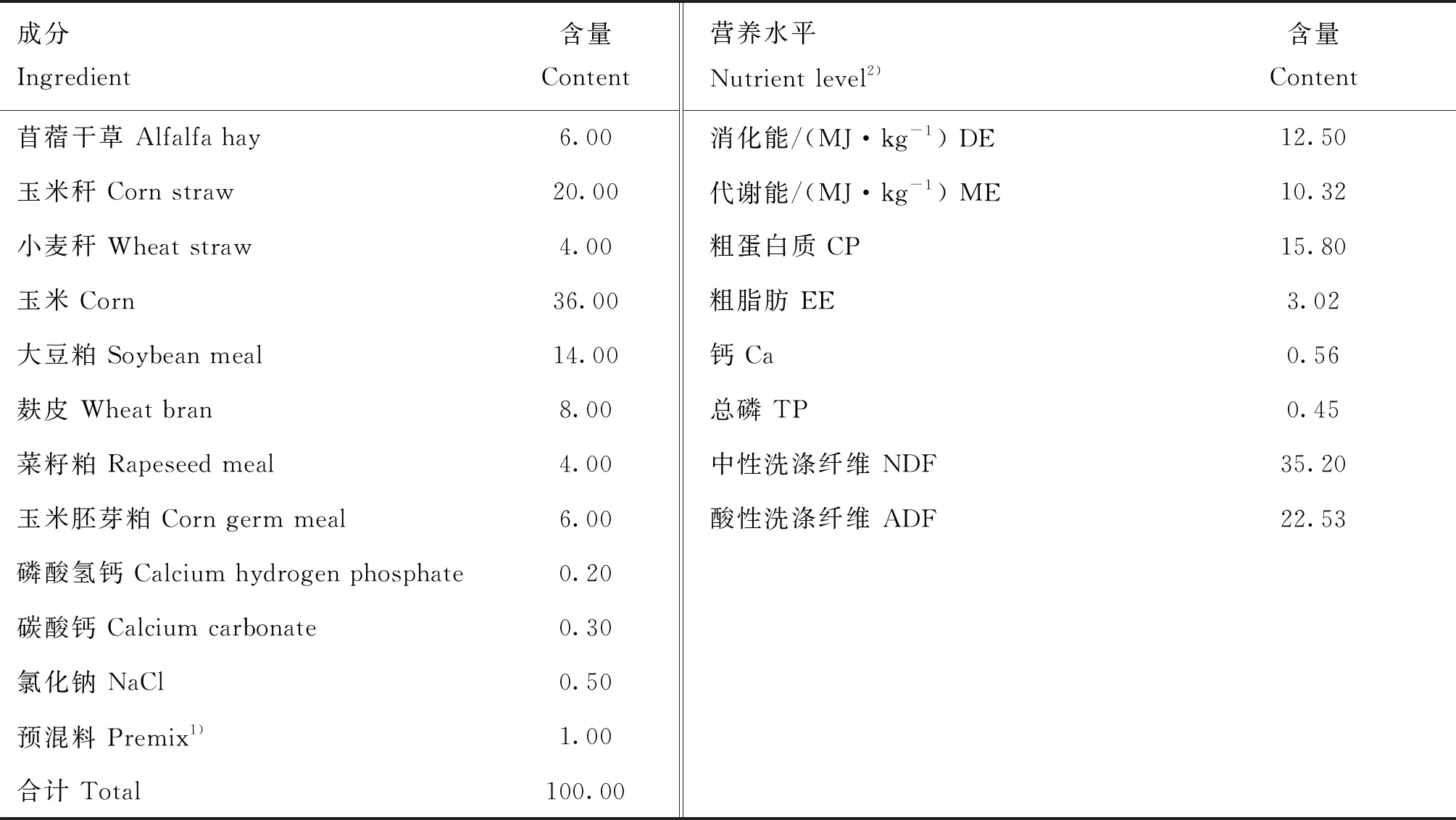
表1 試驗日糧組成及營養水平(干物質基礎)
1.4 測定指標與方法
1.4.1 生長性能 試驗開始前(第0天)、試驗期每隔30 d對試驗羊進行早晨空腹稱重,其中試驗開始前和結束時連續2 d空腹稱重(取平均值),記錄體重數據,并計算每個階段平均日增重(average daily gain,ADG)=(末重-初重)/間隔天數。
試驗期每30 d連續3 d記錄投料量、剩料量,并計算平均日采食量(average daily feeding intake,ADFI)、料重比(F/G)。
1.4.2 血清脂肪因子 試驗期第0、30、90、120、150、180、210、240、270天早晨7:00在試驗羊左靜脈進行空腹采血。采血采用未加抗凝劑的5 mL離心管,常溫靜置30 min后,2 150×g離心10 min,吸取上清液分裝于2個1.5 mL EP管中,置于-80 ℃保存。
血清腫瘤壞死因子α(tumor necrosis factor-α,TNF-α)、白細胞介素6(interleukin- 6,IL-6)、瘦素(leptin,LEP)、脂聯素(adiponectin,ADPN)、抵抗素(resistin,RETN)采用雙抗體一步夾心法酶聯免疫吸附試驗(ELISA)測定,使用由黃石艾恩斯生物科技有限公司提供的羊專用試劑盒,上樣50 μL于酶標儀(上海科華,ST-360),讀取450 nm的OD值,代入回歸方程計算出樣品的濃度;血清游離脂肪酸(nonesterified fatty acid,NEFA)采用銅試劑比色法,使用南京建成生物工程研究所游離脂肪酸測試盒;血清甘油三酯(Triglyceride,TG)、葡萄糖(glucose,GLU)分別采用甘油磷酸氧化法、氧化酶法測定,用全自動生化儀(希森美康,BX-3010)測定,使用上海科華生物工程有限公司試劑盒。
1.4.3 屠宰性能和肉品質 屠宰率:試驗羊宰前12 h禁食,2 h禁水,空腹稱重屠宰后,去頭、毛皮、內臟及前肢膝關節和后肢趾關節以下部分,整個軀體(包括腎及其周圍脂肪)靜置30 min后稱胴體重,計算屠宰率=胴體重/宰前活重×100%。
GR值:第12~13肋骨之間距離背中線11 cm處組織厚度,用游標卡尺測定。
眼肌面積:用直尺測定倒數第1與第2肋骨之間脊椎上眼肌的高和寬,計算眼肌面積=眼肌高度×眼肌寬度×0.7。
背膘厚度:第12與第13胸椎結合處的背部皮下脂肪的厚度,用直尺測定。
剪切力:參考《肉嫩度的測定 剪切力測定法》(NYT 1180—2006),取右側后腿肉一塊(≥6×3×3 cm),除去表面脂肪、筋膜,裝入塑料薄膜袋中包扎好,放在15~16 ℃條件下24 h,然后置于4 ℃熟化24 h,取出在室溫下放置1 h。將熱電耦溫度計插入肌肉中心部,再包扎好肉樣,保持袋口向上,放入功率1 500 W的80 ℃恒溫水浴鍋中,待肉羊中心溫度達到70 ℃為止(約45 min)。取出肉樣,置于4 ℃冷卻。按與肌纖維呈垂直方向切取厚度為2.5 cm,長寬為5 cm的肉片,再用直徑1.27 cm的圓形取樣器順肌纖維方向鉆切肉樣塊,做3個重復,計算平均值。
熟肉率:取右側后腿肉約100 g,剝去表面脂肪稱重,在2 000 W電爐上蒸45 min,室溫懸掛冷卻30~45 min后稱重,3個平行,計算平均值。
采集右側背最長肌樣品,測定pH、肉色、失水率,每個樣品做3個平行,取平均值。
1.4.4 脂肪沉積與器官指數 尾巴大小指數:試驗羊屠宰去毛皮、四肢后,用軟尺量尾長、尾根部寬、尾中部寬(尾巴中間最寬處的寬度)[13]、尾周長(尾巴最寬處一周長度)及尾巴重量,并計算各指標指數。
各部位脂肪沉積:試驗羊屠宰去毛皮、四肢后,剝離皮下、尾部、心組織周圍、腎組織周圍、大網膜、腸系膜、腹部脂肪,用吸水紙吸干水分,進行稱重,并計算內臟脂肪和總脂肪以及各部位脂肪指數。內臟脂肪=心組織周圍脂肪+腎組織周圍脂肪+大網膜脂肪+腸系膜脂肪+腹部脂肪;總脂肪=內臟脂肪+尾部脂肪+皮下脂肪;各部位脂肪指數(%)=該部分脂肪重/宰前活重×100。
1.5 數據處理與統計分析
運用 SPSS 軟件(IBM SPSS Statistics 19)中T檢驗。所有數據以“平均值±標準誤(Mean±SE)”表示,顯著水平為0.05(P<0.05)。
2 結 果
2.1 斷尾對蘭州大尾羊生長性能的影響
由圖1A可知,蘭州大尾羊羔羊斷尾后的30 d內,斷尾組(T組)羔羊日增重135.93 g,顯著低于對照組(C組)的208.15 g(P<0.05);31~60 d內,斷尾組日增重168.52 g,數值上低于對照組的174.44 g,但差異不顯著(P>0.05);61~90 d、91~120 d內,斷尾組日增重分別達到207.03和308.52 g,分別較對照組的180.36和291.85 g提高了14.79%和5.71%,差異不顯著(P>0.05);121~150 d內,斷尾組日增重223.33 g,稍低于對照組的238.89 g,無顯著差異(P>0.05);151~240 d內兩組試驗羊的日增重在223.12~238.89 g,無顯著差異(P>0.05)。由圖1B可知,由于斷尾對羔羊早期(斷尾30 d內)生長影響,斷尾組試驗羊的體重在斷尾后的前3個階段(斷尾90 d內)顯著低于未斷尾組,但隨著斷尾后時間的延長,兩組試驗羊的體重無顯著差異(P>0.05)。

數據點上無字母表示差異不顯著(P>0.05),不同小寫字母表示差異顯著(P<0.05),下同
由表2可知,蘭州大尾羊羔羊斷尾后的料重比在斷尾后的61~90 d內,顯著小于對照組(P<0.05),其余階段差異不顯著(P>0.05)。斷尾組除了0~30 d、121~150 d、181~240 d外,各階段料重比及平均料重比在數值上小于對照組。

表2 斷尾對蘭州大尾羊料重比的影響
2.2 斷尾對蘭州大尾羊血清脂肪因子的影響
斷尾后,短期內(0~60 d)蘭州大尾羊血清TNF-α較對照組顯著升高(P<0.05),61~240 d雖然斷尾組血清中濃度一直高于對照組,但差異不顯著(圖2C,P>0.05)。試驗期內,血清GLU(圖2A)、LEP(圖2E)、RETN(圖2F)、ADPN(圖2H)斷尾組在數值上大于對照組,并且隨著試驗期的增加,斷尾組和對照組之間的差異增大,但差異不顯著(P>0.05)。血清TG斷尾后0~30 d內斷尾組高于對照組,61~210 d內斷尾組低于對照組,211~240 d內斷尾組高于對照組,但差異不顯著(圖2B,P>0.05)。血清IL-6斷尾后0~30 d內斷尾組在數值上高于對照組,但差異不顯著(P>0.05),31~240 d內斷尾組與對照組變化趨勢一致(圖2D)。試驗期內,血清NEFA在兩組間變化趨勢一致,兩組間無顯著差異(圖2I,P>0.05)。
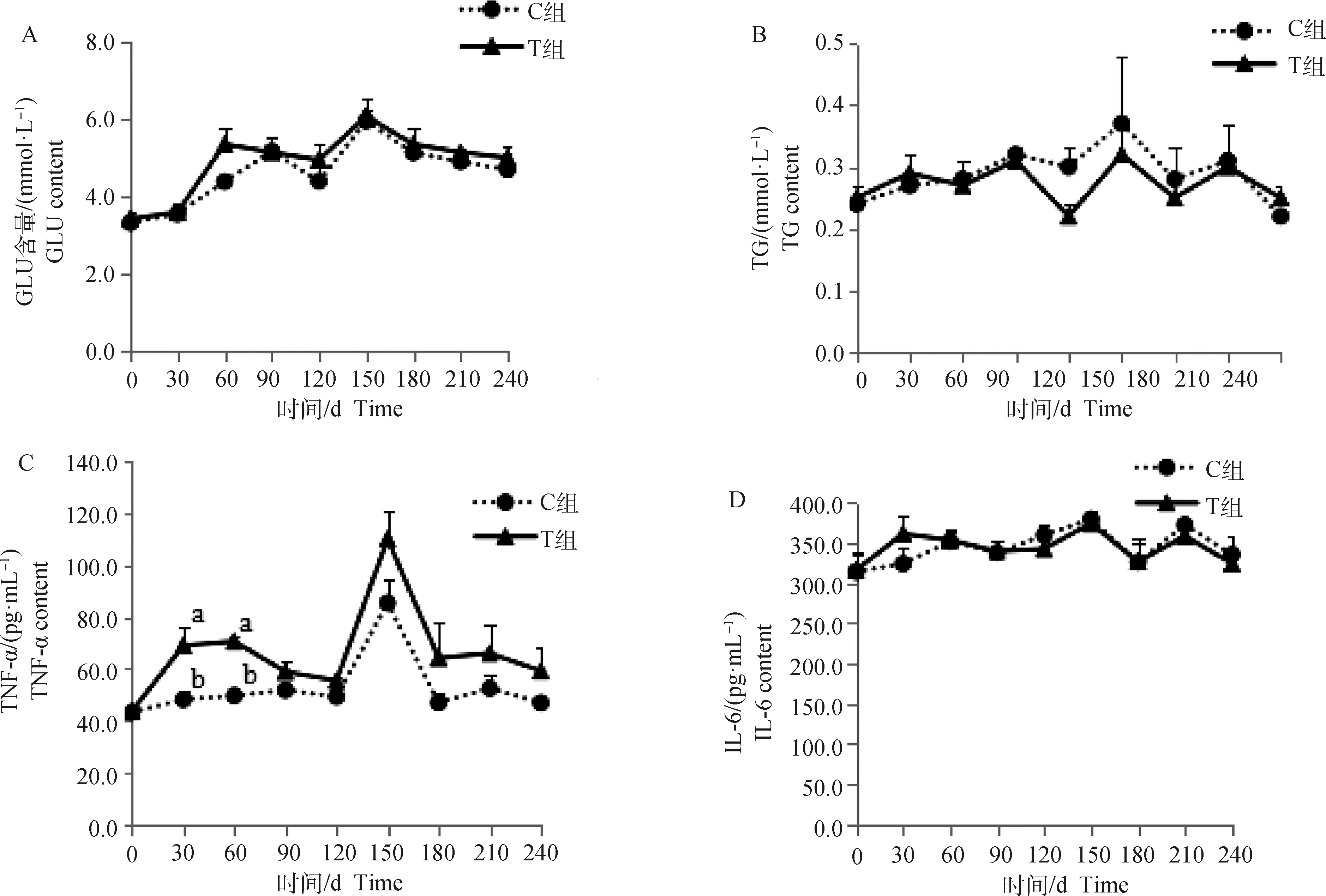
A~I.分別為血清GLU、TG、TNF-α、IL-6、LEP、RETN、ADPN和NEFA含量
2.3 斷尾對蘭州大尾羊脂肪沉積和器官指數的影響
由表3可見,斷尾對蘭州大尾羊各部位脂肪沉積有影響。斷尾后,蘭州大尾羊尾部脂肪指數、總脂肪指數與對照組相比,顯著降低(P<0.05),分別降低了74.31%和34.02%。斷尾組皮下脂肪指數、腎周脂肪指數、腹部脂肪指數和內臟脂肪指數顯著高于對照組(P<0.05),心臟脂肪指數、大網膜脂肪指數和腸系膜脂肪指數兩組間無顯著差異(P>0.05)。
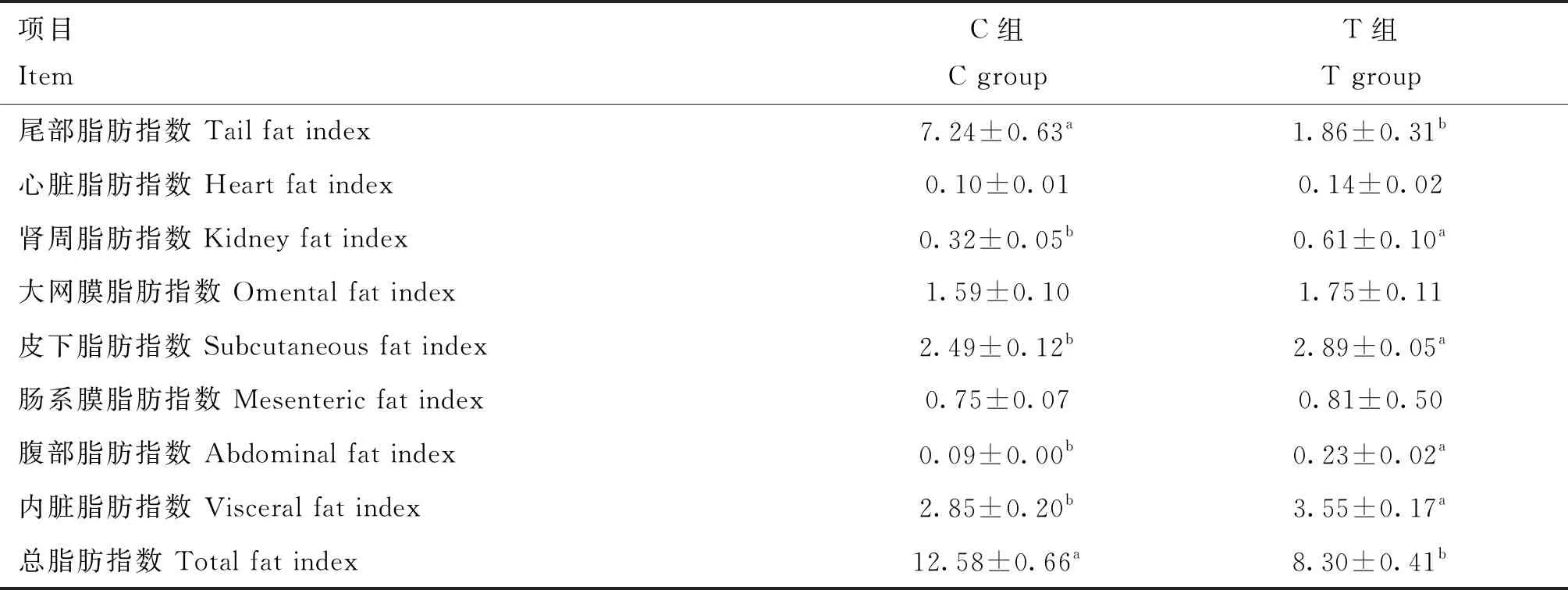
表3 斷尾對蘭州大尾羊脂肪沉積指數的影響
由表4可見,早期斷尾使蘭州大尾羊尾長指數、尾中寬指數、尾周長指數和尾重指數顯著降低(P<0.05),而對尾根寬指數無顯著影響(P>0.05),兩組試驗蘭州大尾羊尾巴大小見圖3。斷尾后,蘭州大尾羊頭重指數顯著升高(P<0.05),蹄重指數、毛皮重指數兩組間無顯著差異(P>0.05)。
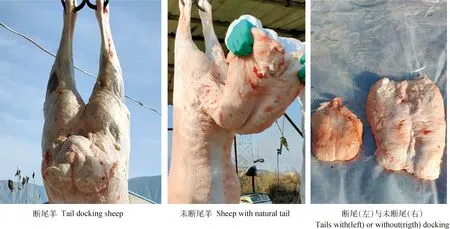
圖3 斷尾與未斷尾的蘭州大尾羊尾巴

表4 斷尾對蘭州大尾羊尾部及器官指數的影響
2.4 斷尾對蘭州大尾羊屠宰性能和肉品質的影響
斷尾組羊的屠宰率顯著大于對照組(P<0.05),GR值、背膘厚、眼肌面積差異不顯著(P>0.05,表5)。斷尾組背最長肌的肉色(亮度、紅度、黃度)和pH與對照組無顯著差異(P>0.05),斷尾組羊背最長肌剪切力顯著小于對照組(P<0.05),失水率和蒸煮損失兩組間無顯著差異(P>0.05,表6)。

表5 斷尾對蘭州大尾羊屠宰性能的影響
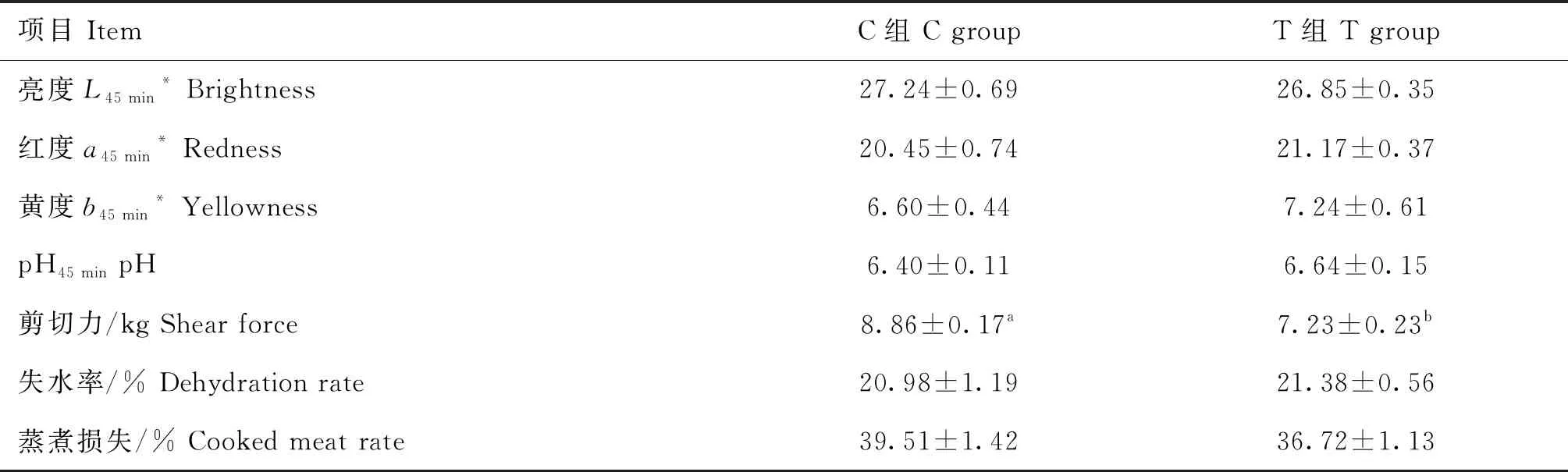
表6 斷尾對蘭州大尾羊肉品質的影響
3 討 論
3.1 斷尾對蘭州大尾羊生長性能、屠宰性能和肉品質的影響
在舍飼條件下,脂尾型綿羊早期斷尾不僅利于交配和避免羊糞污染羊毛,而且可以提高屠宰性能和飼料轉化率(料重比)。日增重和飼料轉化率是衡量家畜生產性能的主要指標,屠宰性能則直接反映家畜產肉能力和肌肉營養沉積分布[14]。而肌肉的顏色、嫩度、pH、失水率、蒸煮損失等是肌肉品質的主要指標[15],其中剪切力是測定肉嫩度的重要指標,pH是肉類酸度的重要指標,一般介于5.8~6.2之間[16]。Wang等[17]將1日齡的脂尾型綿羊斷尾,待46日齡斷奶后放牧至6月齡屠宰,與未斷尾組相比,斷尾能極顯著地提高日增重、胴體重、屠宰率和背膘厚,顯著地提高眼肌面積;同時,顯著提高背最長肌的L*值和肌內脂肪含量,降低剪切力。劉政等[18]對2月齡的蘭州大尾羊和蒙古羊進行斷尾后發現,斷尾組的蘭州大尾羊和蒙古羊日增重、宰前活重和胴體與未斷尾組相比,均有提高的趨勢,而采食量有降低的趨勢。周瑞等[19]的研究結果表明,斷尾脂尾型羊的背膘厚、GR值有增大的趨勢,而剪切力、失水率有減小的趨勢。Bing?l等[20]對1日齡脂尾型公羔斷尾,在其斷奶后進行70 d的育肥試驗,結果顯示,斷尾組的日增重、屠宰率顯著高于對照組,斷尾組的眼肌面積、肌內脂肪含量在數值上增大,但無統計學差異。也有研究結果顯示,早期斷尾對宰前活重、日增重、屠宰率等無顯著影響[21]。
在本試驗中,蘭州大尾羊羔羊在斷尾后的最初30 d內,斷尾組日增重顯著低于未斷尾組,以后31~60 d、61~90 d、91~120 d、121~240 d各階段兩組羊試驗羊的日增重無顯著差異;斷尾組試驗羊的體重在斷尾后的90 d內顯著低于未斷尾組,但隨著斷尾后時間的延長,兩組試驗羊的體重無顯著差異;斷尾組料重比在斷尾后的61~90 d內,顯著小于未斷尾組,其余階段差異不顯著。這可能是因為在斷尾的最初30 d內,斷尾對羔羊造成一定的損傷,由于斷尾應激使免疫調節機制被激活,使部分營養物質用于免疫反應,從而降低了其生長性能,從而使日增重較對照組減小,隨著斷尾損傷的愈合,在61~120 d內斷尾羊出現補償生長,經過補償生長后兩組羊的體重無顯著差異。在整個試驗期內,斷尾羊的料重比在數值上小于未斷尾組,這可能是斷尾后,尾部沉積脂肪減小,從而使總脂肪沉積也減小,導致飼料轉化率升高的原因。
與未斷尾組相比,斷尾蘭州大尾羊的屠宰率顯著升高,剪切力顯著降低,背膘厚、GR值在數值上增大,這與上述研究結果相似。前人研究認為,脂尾型綿羊尾部脂肪發育較早,羔羊生長發育早期尾部脂肪沉積較快[22],并且尾部脂肪沉積效率高于其他部位脂肪沉積效率[23],具有局部沉積的生物學效應[24]。同時,沉積1 kg脂肪所耗飼草料相當于生產2 kg瘦肉所需的飼草料[3]。本試驗中,蘭州大尾羊早期斷尾人為干預后,使得尾部脂肪代謝通路受損并且部分被阻斷,動物攝入的能量更多的用于機體其他部位的生長發育和脂肪沉積,多余能量便較多地沉積到皮下、腹部和肌內等部位,使尾部沉積的脂肪量降低,胴體總脂肪、皮下脂肪以及肌內脂肪含量增加,屠宰率、背膘厚、GR值、嫩度增大,剪切力降低,從而提高了胴體品質和肉品質。
3.2 斷尾對蘭州大尾羊脂肪沉積分布的影響
動物的脂肪組織分為白色脂肪和棕色脂肪,白色脂肪全身各處都有分布,主要分布于軀干、四肢、皮下、腹部和內臟,線粒體少,主要儲存甘油三酯,是機體能量儲備中心,同時還分泌多種脂肪因子,脂肪因子在調節機體飽腹感、脂質代謝、胰島素敏感性方面具有重要的作用[25]。棕色脂肪主要分布在肩胛、頸部和胸腔,富含線粒體,主要參與機體的適應性產熱[26-28]。畜體脂肪的沉積分布是影響胴體品質和肉品質的重要因素,過多的脂肪沉積在尾部、皮下、腹部、內臟等部位會降低胴體品質,而脂肪沉積在肌肉間則可以提高肌肉品質,肌內脂肪含量是評價肉品質最主要的質量指標,與肉的嫩度、色澤、多汁性和風味有關[29-32]。王立艷等[9]研究結果表明,灘羊早期斷尾后,更多的脂肪沉積在皮下、肌內和腎周,提高了肉的嫩度,改善了肌肉品質。劉政等[18]的研究結果顯示,斷尾使蘭州大尾羊更多的脂肪沉積到了皮下、睪丸、腎周和腸胃周圍,且總脂肪重顯著降低。Tilki等[21]的研究結果也認為,脂尾型綿羊斷尾后,腎周脂肪和腸系膜脂肪沉積也顯著提高。Wang等[17]的研究表明,斷尾使脂尾型綿羊肌內脂肪極顯著升高。
綿羊尾部脂肪沉積是經過長期的自然選擇和人工選擇,在進化過程中形成的穩定遺傳的表型特征,受激素、細胞因子、基因、營養等多因素的調控[22]。研究發現,哺乳動物的脂肪細胞由胚胎期具有多向分化能力的間充質干細胞在成脂因子的誘導下發育而來,其數量在出生時基本恒定,出生后,脂肪的沉積主要靠前脂肪細胞的增大和分化實現[33],因此,從脂肪組織發育的角度看,脂肪細胞的增殖和成脂分化,尤其具有分化潛能的前脂肪細胞的增殖分化在脂肪沉積過程中顯得尤為重要[34],脂肪組織中前脂肪細胞在營養、激素等刺激下,不斷的進行增殖分化以及成熟脂肪細胞的增大是脂肪沉積的主要原因。在常規的營養和外界環境條件下,脂肪細胞的數量、體積、分化速度以及在尾部、皮下、腹部等部位沉積分布會在一定的范圍之內達到平衡和穩態[35]。同時,脂尾型綿羊尾部脂肪沉積較早且幼齡羊尾部沉積脂肪效率更高[22]。本試驗對脂肪沉積部位進行人為干預,蘭州大尾羊羔羊5日齡斷尾處理后,一方面可能是在蘭州大尾羊幼齡期,也就是尾部脂肪發育的關鍵期,斷尾使尾部脂肪代謝通路受損并且部分被阻斷,使得原本沉積在尾根部的脂肪部分轉移到皮下、腹部、內臟等部位,造成尾根部脂肪降低,皮下、腹部、腎周和內臟總脂肪升高;另一方面,斷尾使尾部脂肪代謝通路部分被阻斷,尾巴減小,尾部沉積的脂肪降低,在相同的飼養管理條件下,多余的能量更多的沉積在皮下、腹部、內臟和肌內等部位,從而改變機體脂肪沉積分布。所以,長脂尾型綿羊早期斷尾干預可以作為脂肪沉積部位干預的一種有效手段。
3.3 斷尾對蘭州大尾羊脂肪因子的影響
脂肪組織不僅是重要的能量儲存和代謝器官,也是重要的內分泌器官,其分泌的多種具有生物活性的脂肪因子能夠通過自分泌、旁分泌及內分泌的方式介導一系列的信號轉導通路,在調節機體能量平衡、脂質代謝、糖代謝等方面具有重要的作用[36-38]。脂肪因子還可通過不同的靶點反饋調節脂肪細胞的發育分化,在脂肪沉積發生過程中促進脂肪細胞增殖分化的脂肪因子分泌水平增加,而抑制分化的脂肪因子下降,促進脂肪的沉積,反之則抑制脂肪細胞的增殖分化[39]。目前發現脂肪細胞能分泌600多種多肽物質[40],包括TNF-α、IL-6、LEP、ADPN、RETN、VEGF等,這些多肽物質共同構成了一個非常復雜的內分泌網絡系統[41-43]。TNF-α、IL-6是脂肪細胞和免疫巨噬細胞分泌的炎性脂肪因子,這些因子除了參與免疫反應外,還可以誘發機體局部和全身的脂質沉積炎癥反應,參與脂質代謝的調節[41]。LEP、ADPN、RETN都是脂肪細胞分泌的脂肪因子,與脂肪細胞之間存在一系列的反饋調節機制,LEP除了通過下丘腦的攝食中樞抑制能量攝入外,調控能量平衡從而調節脂肪沉積外,還通過抑制前脂肪細胞分化發育和CEB-α和PPAR-γ的表達,減少細胞內甘油三酯的積聚,最終抑制脂肪細胞的生長發育[44]。ADPN和RETN能反饋刺激脂肪細胞的發育分化,從而調節脂質沉積。TG、GLU也可以反映動物體脂代謝情況[16],TG是肌內脂肪含量的主要決定因素[45-46],GLU是機體對糖的吸收、運轉和代謝動態平衡狀態的反映,也是機體能量代謝水平的體現。
動物在正常生長發育過程中產生的分解代謝物最終會進入到血液里,因而血清相關指標能夠反映動物機體新陳代謝的情況[47]。Eicher等[48]研究認為,小母牛血漿中TNF-α在斷尾后的0、144 h顯著升高。Prunier等[49]研究結果顯示,仔豬斷尾后,血漿GLU濃度不受影響。本研究結果顯示,蘭州大尾羊斷尾后0~60 d內血清TNF-α濃度較未斷尾組顯著升高,之后斷尾組在數值上一直高于未斷尾組。這可能一方面是斷尾激活了免疫系統,導致巨噬細胞和淋巴細胞分泌TNF-α,隨著斷尾損傷時間的延長,斷尾造成的炎癥反應降低;另一方面,斷尾使尾部脂肪代謝通路部分被阻斷,導致脂肪細胞分泌的TNF-α誘導前脂肪細胞和脂肪細胞的凋亡。斷尾組血清GLU、LEP、RETN、ADPN在數值上大于未斷尾組。這可能是在相同飼養管理條件下,通過斷尾干預后,斷尾組尾部脂肪沉積降低,沉積在機體其他部位的脂肪相對量增加,導致脂肪細胞分泌的脂肪因子增加,從而反饋調節脂肪的沉積。
4 結 論
4.1在本試驗條件下,蘭州大尾羊早期斷尾(5日齡)可使尾長指數、尾中寬指數、尾周長指數和尾重指數顯著降低,同時,斷尾改變脂肪的沉積分布,斷尾后尾部脂肪指數、總脂肪指數顯著降低;而皮下脂肪指數、腎周脂肪指數、腹部脂肪指數和內臟脂肪指數顯著升高。
4.2斷尾可以提高屠宰率和改善肉品質,蘭州大尾羊早期斷尾后屠宰率顯著提高,背最長肌剪切力顯著降低。
