牛源壞死梭桿菌43K外膜蛋白的黏附特性研究
2023-02-27 14:19:00賀顯晶王志慧郭東華
畜牧獸醫(yī)學報 2023年2期
賀顯晶,劉 嬌,王志慧,武 瑞,郭東華*
(1.黑龍江八一農墾大學動物科技學院,大慶 163319; 2.佳木斯大學,佳木斯 154007)
牛源壞死梭桿菌(Fusobacteriumnecrophorum,F(xiàn)n)是一種革蘭陰性多形桿狀專性厭氧菌,是牛肝膿腫、腐蹄病和壞死性喉炎的主要致病菌,同時在奶牛乳房炎和子宮內膜炎的發(fā)生中也具有重要作用,在牛群感染中具有普遍性的趨勢。細菌黏附宿主細胞是病原菌建立感染的關鍵步驟,而有關于牛源壞死梭桿菌黏附相關蛋白研究尚屬探索階段,因此深入開展牛源壞死梭桿菌相關黏附蛋白研究,對闡明牛壞死梭桿菌病的致病機制奠定理論基礎,也為動物厭氧菌病的機制研究提供參考。
外膜蛋白(outer membrane protein,OMPs)是革蘭陰性細菌外膜的主要結構,在細菌黏附宿主細胞中發(fā)揮重要作用。壞死梭桿菌根據(jù)DNA同源性分為F.necrophorumsubsp.necrophorum(Fnn亞種)和F.necrophorumsubsp.funduliforne(Fnf亞種)[1-2]。不同亞種壞死梭桿菌的OMPs譜存在很大差異,這也解釋了不同亞種在毒性及參與壞死梭桿菌感染發(fā)病機制方面的差異[3]。43K OMP(43 ku outer membrane protein)是本課題組首次鑒定的Fnn亞種牛源壞死梭桿菌的主要OMPs,其與梭菌屬其他成員的OMPs表現(xiàn)出高度的相似性[4]。進一步研究發(fā)現(xiàn),43K OMP在牛源壞死梭桿菌不同分離株中廣泛存在[5]。因與壞死梭桿菌同菌屬的其他厭氧菌如具核梭桿菌、放線桿菌等對宿主細胞的黏附是由其表面的外膜蛋白介導[2, 6-8]。據(jù)此,作者推測43K OMP是牛源壞死梭桿菌的一種重要黏附相關蛋白,在牛源壞死梭桿菌致病中發(fā)揮重要作用。目前,有關于牛源壞死梭桿菌43K OMP的功能研究尚屬探索階段,為更好地揭示43K OMP的黏附功能,本研究通過構建43K OMP原核表達系統(tǒng),進一步研究其對細菌黏附細胞的影響及43K OMP對細胞的黏附作用,為牛源壞死梭桿菌43K OMP的生物學功能拓展研究奠定理論基礎。
1 材料與方法
1.1 菌株、細胞及載體
牛源壞死梭桿菌A25菌株(Fusobacteriumnecrophorum,F(xiàn)nn亞種,ATCC 25286)保存于黑龍江八一農墾大學獸醫(yī)分子病理學實驗室;牛子宮內膜上皮細胞(BNCC 340413)購于北京北納創(chuàng)聯(lián)生物技術有限公司;牛乳腺上皮細胞為MAC-T細胞系,鼠乳腺上皮細胞(EpH4-Ev)由吉林大學農學部動物生理教研室惠贈,小鼠肝細胞(AML12 SCSP-550)購于中國科學院干細胞庫;pET-32a表達載體、E.coliBL21(DE3)感受態(tài)細胞由黑龍江八一農墾大學病理實驗室保存;43K OMP多克隆抗體血清、牛源壞死梭桿菌高免血清、43K OMP單克隆抗體、兔陰性血清及鼠陰性血清由黑龍江八一農墾大學獸醫(yī)分子病理學實驗室制備并保存[9-11]。
1.2 主要試劑
細菌基因組DNA提取試劑盒、質粒小提取試劑盒購于天根生物技術有限公司,ExTaqDNA聚合酶購于大連寶生物工程有限公司,DMEM培養(yǎng)基、1640培養(yǎng)基、DMEM/F12培養(yǎng)基、HEPE,LiCl、LDS、SLS、ITS、地塞米松均購于美國Sigma公司,F(xiàn)BS購于CLARK公司,SDS-PAGE凝膠配制試劑盒、DMSO購于索萊寶生物技術有限公司,Alexa Fluor 488標記山羊抗兔IgG、Alexa Fluor 488標記山羊抗鼠IgG購于碧云天生物技術有限公司,辣根過氧化酶標記的山羊抗兔IgG和山羊抗鼠山羊購于武漢三鷹生物技術有限公司,所有引物合成由上海生工生物工程股份有限公司完成。
1.3 細菌的培養(yǎng)
牛源壞死梭桿菌A25菌株在預先還原厭氧的苛養(yǎng)肉湯培養(yǎng)基中37 ℃厭氧培養(yǎng)12 h以上備用。用于重組表達的大腸桿菌BL21 DE3在100 μg·mL-1氨芐青霉素的LB肉湯培養(yǎng)基培養(yǎng)。
1.4 天然43K OMP的提取
參照王志慧[9]和徐晶(Xu)等[12]的方法提取牛源壞死梭桿菌天然43K OMP。牛源壞死梭桿菌A25菌株1∶100接種于FAB液體中厭氧培養(yǎng)至OD600nm為0.6~0.8時,將500 mL菌液用無菌的生理鹽水清洗3次,5 000×g離心5 min,將菌液沉淀用50 mL無菌的PBS(pH=7.2)重懸后置于冰水混合物中,功率60 W,超聲時間6 s,間歇6 s進行超聲破碎1 h。將菌液混合物4 ℃,3 000×g離心30 min,收集上清。收集的上清進行4 ℃,12 000×g離心30 min,收集沉淀。將沉淀用50 mL 0.5%的SLS重懸后,室溫孵育30 min,4 ℃,12 000×g再次離心30 min,棄上清取沉淀。將沉淀用25 mL 2% LDS的20 mmol·L-1的HEPEs-LiCl(pH=7.4)重懸后,4 ℃孵育30 min。提取液經(jīng)4 ℃,12 000×g離心30 min,收集沉淀。將沉淀用2 mL 20 mmol·L-1的HEPEs混合均勻,分裝,提取后的天然43K OMP經(jīng)SDS-PAGE分析后,43K OMP重組蛋白的表達純化復性及OMP的去內毒素處理由北京海蓋德科技有限公司完成。
1.5 43K OMP重組蛋白的表達與純化
參照課題組前期的方法進行克隆表達牛源壞死梭桿菌43K OMP基因[10-11],參照牛源壞死梭桿菌H05菌株43K OMP基因序列(GenBank accession No.JQ740821.1)PCR擴增43K OMP基因,在序列的5′端和3′端加入BamHⅠ和XhoⅠ兩種酶切位點,隨后克隆連接至表達載體pET-32a上,將pET-32a-43K OMP質粒轉化入大腸桿菌BL21 DE3細胞中。將攜帶pET-32a-43K OMP質粒的大腸桿菌BL21 DE3細胞標記為H2019,將H2019經(jīng)IPTG(濃度為1 mmol·L-1)37 ℃誘導表達4 h,超微破碎后,應用蛋白純化試劑盒對重組蛋白進行純化,將純化后的重組蛋白進行Western blot鑒定。重組蛋白純化及復性委托北京海蓋德科技有限公司完成。
1.6 H2019與細胞的黏附試驗
分別將牛子宮內膜細胞、MAC-T細胞、鼠乳腺上皮細胞及鼠肝細胞接種于6孔板,5%CO2培養(yǎng)箱,37 ℃培養(yǎng)至密度為90%以上,棄培養(yǎng)液,PBS洗3次。H2019和pET-32a載體的大腸桿菌 BL21 DE3細胞經(jīng)IPTG 37 ℃誘導3 h后,PBS離心洗3次后,通過麥氏比濁法用不含雙抗的細胞培養(yǎng)基將菌液濃度為1×107CFU·mL-1備用。將各孔與IPTG誘導的H2019和pET-32a載體的大腸桿菌 BL21 DE3細胞37 ℃孵育1 h,棄培養(yǎng)液,PBS洗3~5次,每次3 min,加入1%TritonX-100的PBS 10 min,吸取懸液倍比稀釋后,在含有氨芐抗性的LB固體培養(yǎng)基上37 ℃孵育過夜,菌落計數(shù),每個處理3個重復。
1.7 天然蛋白競爭試驗
將細胞接種于6孔板中(1×107·mL-1),5%CO2培養(yǎng)箱,37 ℃培養(yǎng)至密度為90%以上,每孔加入提取并去內毒素的牛源壞死梭桿菌天然43K OMP(100 μg·孔-1)37 ℃孵育1 h。PBS清洗3次,以去除未黏附的OMP。誘導表達的H2019菌液經(jīng)PBS清洗3次后,用加入10%FBS的細胞培養(yǎng)基重懸,與細胞37 ℃共孵育1 h。PBS清洗后收集細胞懸液倍比稀釋后計數(shù)。未與OMP共孵育的細胞作為對照組。每個處理重復3次。
1.8 抗體抑制試驗
將H2019在37 ℃經(jīng)IPTG誘導3 h后,PBS清洗,用10%FBS的細胞培養(yǎng)基重懸。將1 mL的細菌懸液按照1∶200和1∶50的濃度加入43K OMP多抗或單抗37 ℃搖床內孵育1 h。將細胞接種于6孔板中(1×107·mL-1),5%CO2培養(yǎng)箱中,37 ℃培養(yǎng)至密度為90%以上,經(jīng)PBS清洗后與細菌懸液共孵育1 h。同時設置陰性血清對照組和細菌對照組。PBS清洗3~5次,進行菌落計數(shù)。每個處理重復3次。
1.9 蛋白水解試驗
將細胞接種于6孔板中,待細胞生長密度至90%左右。將H2019在37 ℃經(jīng)IPTG誘導2 h后,分成4等份,分別與蛋白酶K(0、25、50、100 μg·mL-1)37 ℃孵育2 h,PBS清洗3次后,用10% FBS的細胞培養(yǎng)基重懸后,與細胞共孵育1 h,PBS清洗3~5次,以去除未黏附的細菌后進行菌落計數(shù)。每個處理重復3次。
1.10 43K OMP對細胞的黏附試驗
將牛子宮內膜細胞或MAC-T細胞消化按照1×107·mL-1濃度接種于6孔板,37 ℃培養(yǎng)24 h后,細胞生長密度為85%左右,PBS漂洗細胞3次,4%多聚甲醛4 ℃過夜固定,PBST漂洗3次,除去多余的固定液。3%BSA室溫封閉2 h,PBST充分漂洗3次。試驗組加入重組蛋白或天然43K OMP(終濃度為50 μg·孔-1),同時只加PBS的孔作為陰性對照組,室溫孵育1 h。孵育結束后,PBST充分漂洗3次,去除未黏附的蛋白和培養(yǎng)基。加入43K OMP單克隆抗體(稀釋度為1∶800)室溫孵育1 h,PBST充分漂洗3次,去除游離抗體,加入Alexa Fluor 488標記的二抗(1∶500稀釋),37 ℃孵育1 h,PBST充分漂洗3次,倒置熒光顯微鏡下觀察43K OMP蛋白與細胞是否黏附及結合位置。
1.11 數(shù)據(jù)分析
2 結 果
2.1 43K OMP的誘導表達
H2019經(jīng)IPTG誘導后,菌體超聲破碎物經(jīng)SDS-PAGE分析顯示,在59 ku出現(xiàn)特異性的表達條帶,表明43K OMP蛋白成功表達,且目的蛋白以包涵體形式存在于菌體沉淀中(圖1A)。純化蛋白經(jīng)Western blot鑒定,在59 ku處出現(xiàn)明顯的目的條帶(圖1B)。
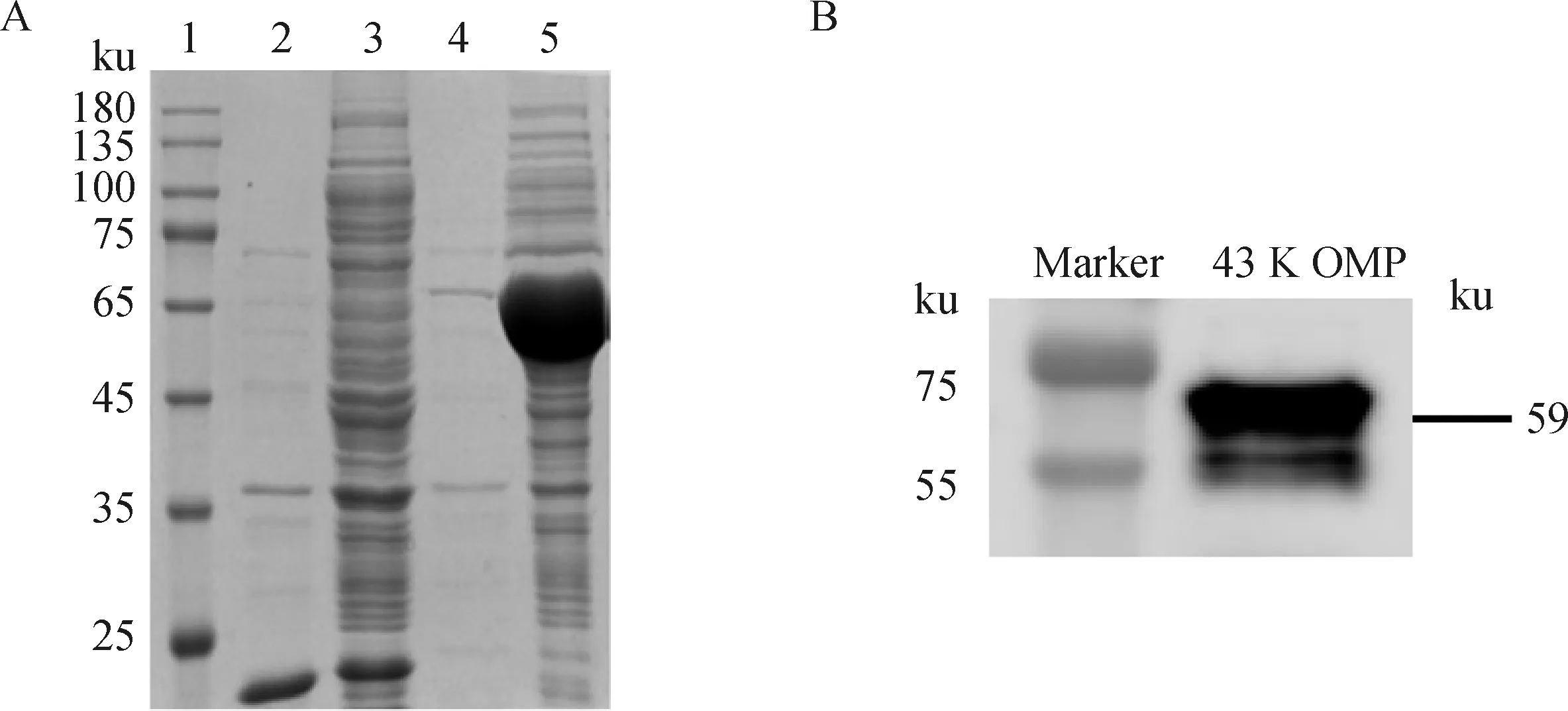
A.43K OMP重組蛋白的誘導表達(1.蛋白質相對分子質量標準;2.誘導后pET-32a超聲破碎上清;3.誘導后pET-32a超聲破碎沉淀;4.誘導后H2019超聲破碎上清;5.誘導后H2019超聲破碎沉淀);B. 43K OMP重組蛋白的Western blot鑒定
2.2 H2019對細胞的黏附試驗結果
黏附試驗結果表明,與空載體對照組孵育的細胞相比,當牛子宮內膜細胞、MAC-T細胞、鼠乳腺上皮細胞、鼠肝細胞與誘導的H2019共孵育后,細胞黏附的細菌數(shù)量均極顯著增加(P<0.01)(圖2)。
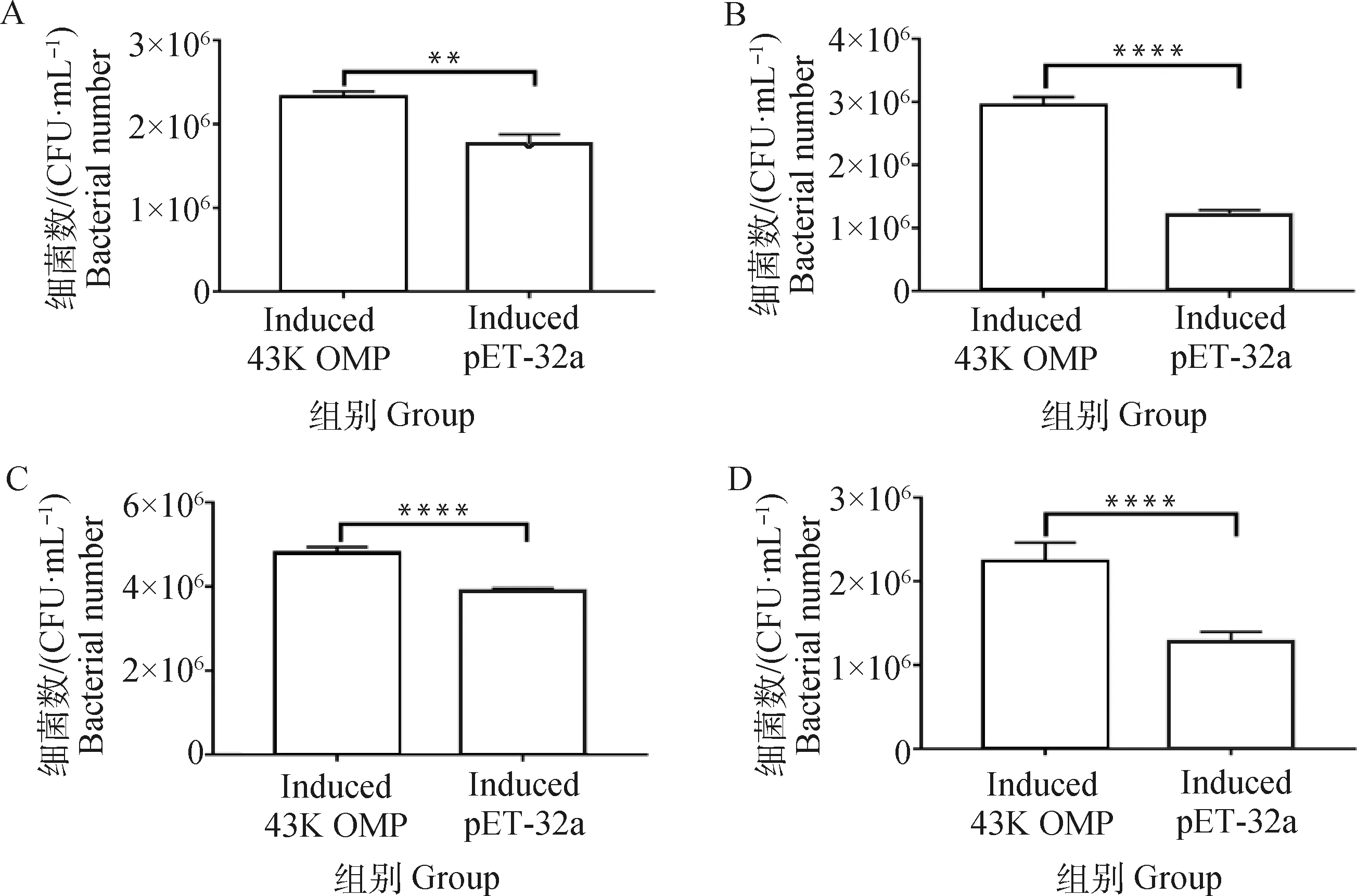
A. BEND;B. MAC-T;C. MMECs;D. AML12; **. P<0.01, ****. P<0.000 1
2.3 天然43K OMP對H2019黏附宿主細胞的影響
與未加入天然43K OMP蛋白的對照組相比,當牛子宮內膜細胞、MAC-T細胞、鼠乳腺上皮細胞、鼠肝細胞與天然蛋白預孵育后,黏附于細胞上的H2019數(shù)量極顯著降低(P<0.01)(圖3)。
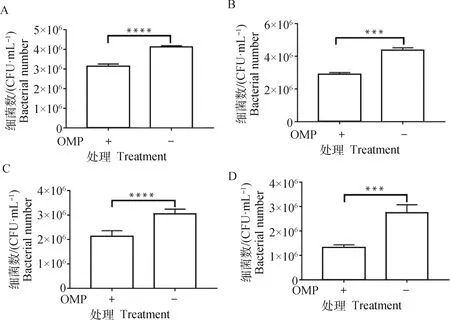
A. BEND;B. MAC-T;C. MMECs;D. AML12; ***. P<0.001, ****. P<0.000 1
H2019與43K OMP多抗共孵育后,黏附于牛子宮內膜細胞、MAC-T細胞、鼠乳腺上皮細胞和鼠肝細胞上的H2019數(shù)量極顯著降低(P<0.01),且在牛子宮內膜細胞降低趨勢更顯著,而陰性血清組與對照組無顯著性差異(P>0.05)(圖4)。
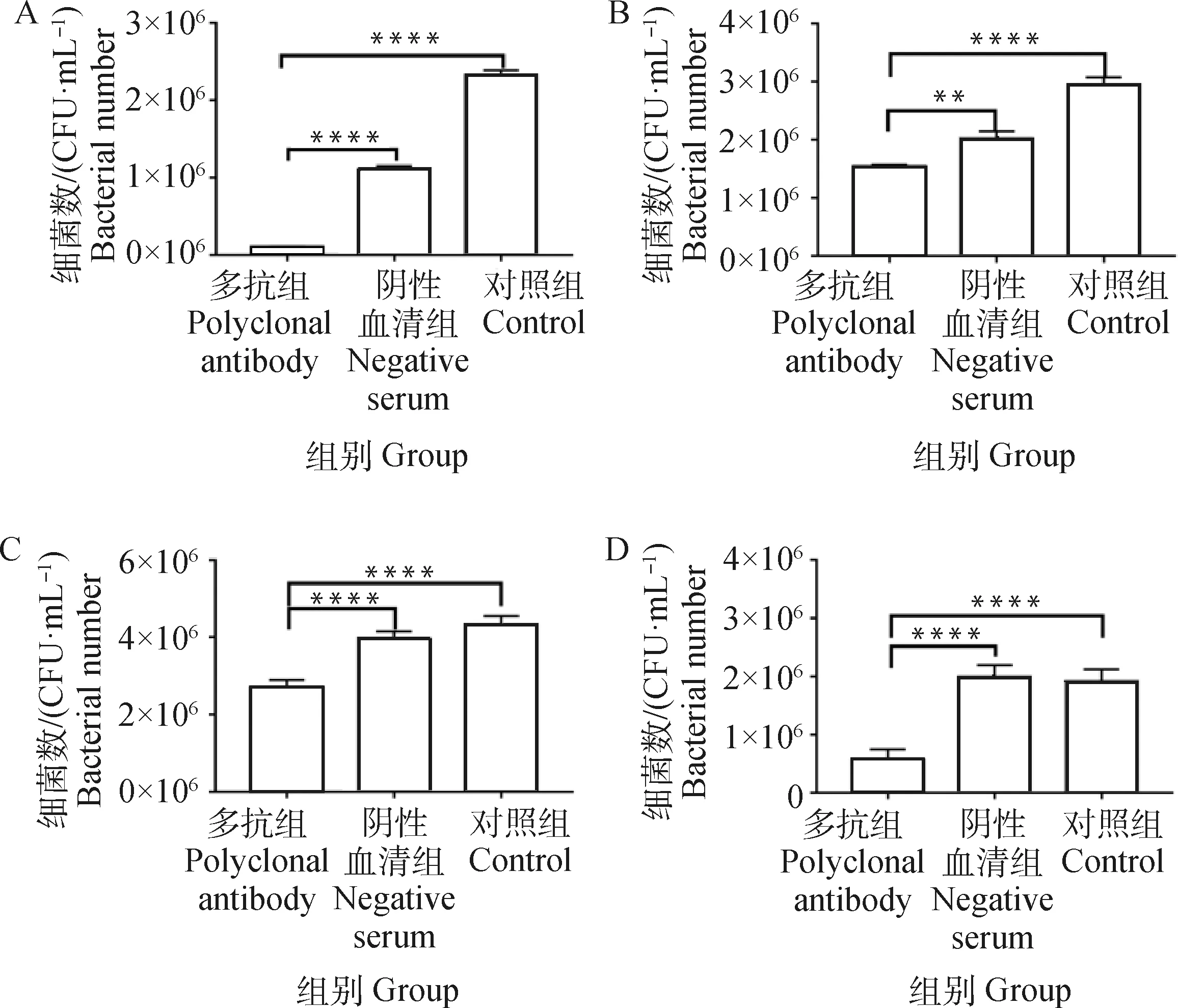
A. BEND;B. MAC-T;C. MMECs;D. AML12; **. P<0.01, ****. P<0.000 1
當H2019與43K OMP單抗共孵育后,黏附于牛子宮內膜細胞、MAC-T、鼠乳腺上皮細胞和AML12細胞的H2019數(shù)量呈現(xiàn)不同程度的降低(P<0.05),而陰性血清組與對照組無顯著性差異(P>0.05)(圖5)。
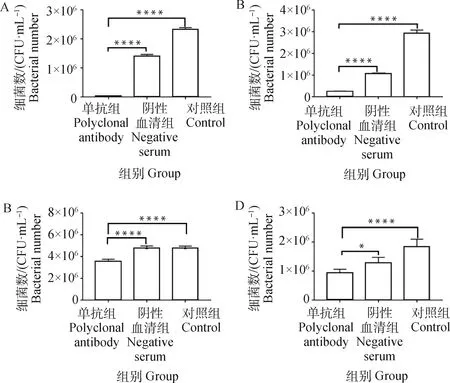
A. BEND;B. MAC-T;C. MMECs;D. AML12; *. P<0.05, ****. P<0.000 1
2.4 蛋白酶K對H2019黏附宿主細胞的影響
將誘導表達43K OMP的H2019與不同濃度的蛋白酶K(0、25、50、100 μg·mL-1)孵育,SDS-PAGE分析顯示:表達的43K OMP量被蛋白酶K按劑量依賴性的方式消化降解(圖6 A)。同時黏附試驗也顯示了黏附于牛子宮內膜細胞、MAC-T細胞、鼠乳腺上皮細胞和鼠肝細胞的H2019數(shù)量也呈現(xiàn)劑量依賴性降低(圖6B、C、D和E)。
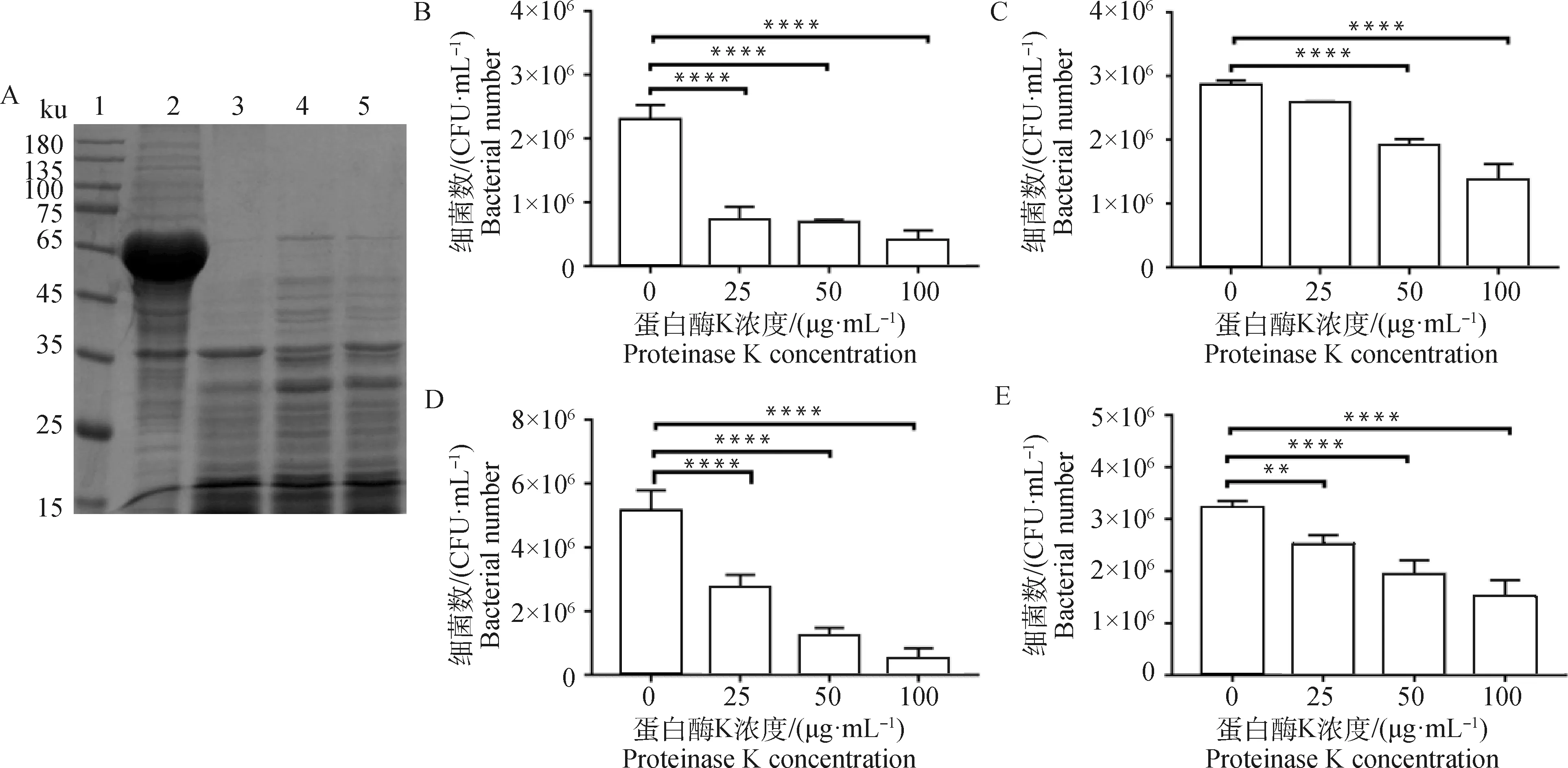
A. 蛋白酶K對H2019誘導表達的影響(1.蛋白質相對分子質量標準;2~5. 蛋白酶K濃度 0、100、50、25 μg·mL-1);B~E. 蛋白酶K對H2019黏附牛子宮內膜細胞 (B)、MAC-T細胞(C)、鼠乳腺上皮細胞 (D) 和AML12細胞 (E) 的影響。**. P<0.01, ****. P<0.000 1
2.5 43K OMP對細胞的黏附作用
將牛乳腺上皮細胞、子宮內膜細胞分別與43K OMP重組蛋白、天然43K OMP蛋白共孵育,間接免疫熒光結果顯示,43K OMP重組蛋白或天然蛋白組均發(fā)出明顯的綠色熒光,而對照組無熒光出現(xiàn),表明牛源壞死梭桿菌43K OMP能黏附牛子宮內膜細胞和乳腺上皮細胞,且綠色熒光分布輪廓與細胞形態(tài)相似(圖7)。

掃描文章首頁OSID碼可查看彩圖
3 討 論
細菌黏附宿主細胞是細菌定植和侵襲的先決條件,細菌對宿主細胞的特異性黏附是病原菌感染的重要步驟。牛源壞死梭桿菌作為條件致病菌,對瘤胃上皮細胞及肝竇內的血管內皮細胞、蹄部損傷的皮膚組織細胞、乳腺上皮細胞和子宮內膜細胞的黏附是牛肝膿腫、腐蹄病、乳房炎及子宮內膜炎等疾病感染的重要步驟。在梭桿菌屬中,OMPs對壞死梭桿菌的黏附和感染至關重要。與牛源壞死梭桿菌同菌屬的具核梭桿菌中,OMPs在細菌黏附細胞、細菌共聚及生物膜形成中發(fā)揮重要作用。其中,RadD能結合變形鏈球菌SpaP,介導其與變形鏈球菌或酵母念珠菌的共聚[13-15]。FadA促進具核梭桿菌黏附KB、CHO細胞和胎盤內皮細胞[16-17],增強牙齦卟啉單胞菌和放線菌的黏附入侵能力[18-19]。Fap2蛋白促進具核梭桿菌與癌細胞的識別和結合是直腸癌發(fā)病的重要機制[20]。而與43K OMP同源性高的具核梭桿菌FomA蛋白在具核梭桿菌黏附宿主細胞和外膜囊泡致病機制中發(fā)揮重要作用[21-23]。因此,推斷43K OMP在牛源壞死梭桿菌上黏附內皮細胞具有潛在作用。
在有關于牛源壞死梭桿菌黏附機制的相關研究中,早期認為牛源壞死梭桿菌能通過血凝素黏附于Vero表面,且牛源壞死梭桿菌黏附瘤胃上皮細胞也與血凝素相關[24-25]。通過抗血凝素血清能降低牛源壞死梭桿菌與兔、鼠等易感動物細胞的黏附[26]。近年來,有學者發(fā)現(xiàn)胰酶能顯著降低壞死梭桿菌黏附腎上腺血管內皮細胞的能力,表明介導壞死梭桿菌黏附細胞的是蛋白質,推測起作用的可能是OMPs[27]。同時,對Fnn亞種和Fnf亞種壞死梭桿菌的OMPs進行系統(tǒng)研究,發(fā)現(xiàn)相對分子質量為17、24、40和74 ku的OMPs可能與壞死梭桿菌黏附細胞有關,且40 ku大小的OMP在壞死梭桿菌黏附細胞中可能發(fā)揮重要作用[27-30]。43K OMP是首次在牛源壞死梭桿菌臨床分離株H05鑒定的,本課題組前期發(fā)現(xiàn)43K OMP對牛源壞死梭桿菌黏附BHK-21細胞上具有一定作用[10,31-32]。同時,應用重組純化的43K OMP免疫小鼠,發(fā)現(xiàn)43K OMP具有一定的保護性,且43K OMP與牛源壞死梭桿菌Lkt和Hly截短表達蛋白PL-4和H2共同制備的亞單位疫苗保護性更好[33]。
本研究中,以牛子宮內膜細胞、乳腺上皮細胞、鼠乳腺上皮細胞和鼠肝細胞為細胞模型,為進一步驗證牛源壞死梭桿菌43K OMP對不同宿主細胞的黏附特性,將43K OMP基因克隆到pET-32a載體中,隨后在大腸桿菌BL21DE3細胞上表達該蛋白。當攜帶重組質粒(H2019)的大腸桿菌經(jīng)IPTG誘導表達后,與細胞的黏附顯著增加,顯示43K OMP的表達對細菌黏附具有促進作用。細胞與天然蛋白共同孵育,細菌與多抗或單抗共孵育后,H2019黏附細胞的數(shù)量明顯降低,天然蛋白或抗體可以降低細胞與H2019的結合。其機制可能是天然蛋白競爭了細胞上的結合位點,抑制H2019與細胞的結合;而抗體中和了H2019表達的43K OMP,降低H2019對宿主細胞的黏附。同時也發(fā)現(xiàn)43K OMP的多抗或單抗并不能完全抑制H2019對細胞的黏附,提示牛源壞死梭桿菌中還存在未發(fā)現(xiàn)的其他黏附相關蛋白。而多抗的抑制效果明顯好于單抗與不同抗體的抗原表位差異存在一定的關系。經(jīng)不同濃度的蛋白酶K處理后,蛋白酶K降解了表達的43K OMP,隨之H2019黏附數(shù)量呈劑量依賴性降低。間接免疫熒光結果也顯示了天然43K OMP和重組蛋白均能特異性黏附于細胞膜表面。因此,43K OMP是牛源壞死梭桿菌的一種重要黏附素,有助于促進牛源壞死梭桿菌對宿主細胞的黏附和定植。但43K OMP介導牛源壞死梭桿菌黏附機制尚未可知,43K OMP是通過宿主細胞表面的受體蛋白互作介導細菌黏附,還是與細胞的多糖作用介導細菌黏附尚未可知,還需要人們進一步研究。深入研究43K OMP介導牛源壞死梭桿菌的黏附機制,將對闡明牛源壞死梭桿菌致病機制具有重要意義。
4 結 論
43K OMP是牛源壞死梭桿菌的一種重要黏附蛋白,在牛源壞死梭桿菌黏附宿主細胞中發(fā)揮重要作用。
