關嶺牛MEF2A基因干擾載體構建及其轉染對成肌細胞的影響
2023-02-27 14:26:54孫金魁許厚強石鵬飛
畜牧獸醫學報 2023年2期
關鍵詞:檢測
孫金魁,許厚強,石鵬飛,阮 涌
(1.貴州大學 高原山地動物遺傳育種與繁殖教育部重點實驗室/貴州省動物遺傳育種與繁殖重點實驗室,貴陽 550025; 2.貴州大學動物科學學院,貴陽550025)
關嶺牛具有體質強壯、抗病力強等優良特性,其自身的獨特基因是生物多樣性的重要組成部分[1-2]。與其它黃牛品種相比,關嶺牛產肉率低、生長遲緩,限制了高原山地畜牧業的發展。肌原性細胞的增殖分化是影響肌肉生長發育的關鍵,肌細胞增強因子2(myocyte enhancer factor2, MEF2)在肌肉細胞中廣泛存在,可將該基因視作肌源性基因表達的主要調節物[3-4]。
研究表明,MEF2通過調節肌肉特異基因的表達影響骨骼肌發育分化進程,MEF2屬于轉錄調節因子MADS-Box家族,該家族包括微小染色體維持蛋白(MCM1)、AGAMOUS、DEFICIENS和血清反應因子(SRF)等成員[5]。MEF2蛋白具備DNA結合活性,擁有結合肌肉肌酸激酶(MCK)基因啟動子中富含A/T的DNA序列的能力[6]。前期研究中,MEF2基因家族在脊椎動物中包括MEF2A、MEF2B、MEF2C和MEF2D四個成員[7],該家族成員結構相似,皆含有中心MEF2結構域、N端結合DNA的MADS區域和C端轉錄激活區域;C末端區域主要由選擇性拼接而成,MEF2因子可發生同源二聚化和多種異源二聚化,以具體的方式參與細胞生命周期中許多重要的轉錄過程[8]。MEF2主要通過MADS-box結構域和MEF2結構域與其他轉錄輔因子的互作來調控其下游靶基因的轉錄活性,具備調控細胞凋亡(包括神經元、肌肉、血管、淋巴細胞)和骨骼在內的多種組織的分化、形態發生和維持的作用[9-10]。
MEF2A基因是具有bHLH(basic helix-loop-helix)結構域的轉錄因子,包含MADS-box和HJURP-C兩個保守結構域,兩個結構域可識別肌肉調控基因啟動區中E-box元件(CANNTG),進而激活下游基因的特異性表達[11-12]。Edmondson等[13]研究表明,在肌再生過程中,MEF2A是最早可檢測到的基因之一,在骨骼肌中高度表達,對肌纖維聚合形成、骨骼肌干細胞和成肌細胞增殖分化具有不可或缺的作用[14-15]。Wu等[16]研究發現,MEF2A具備承接結構蛋白與調控因子的功能,該基因異位表達可結合Capn3啟動子區域的預測位點,二者互作可影響L6成肌細胞的分化模型。MEF2A具備獨特的轉錄活性,能夠維持出生后心肌細胞結構的完整性,Clark和Naya[17]研究證實,MEF2A可通過調節Gtl2/Dio3位點,進而刺激轉錄輔因子CITED2來促進心肌細胞的增殖;此外,MEF2A可通過激活SIRT1的表達來保護血管內皮細胞[18]。Blixt等[19]研究報道,MEF2A基因在小鼠破骨細胞模型中具有性別特異性調節作用。因此,MEF2A對于肌肉組織生長發育的調控作用因組織類型而異,對骨骼肌成肌細胞增殖分裂具有重要作用。
目前,有關MEF2A基因在關嶺牛(Bostaurus)成肌細胞上的研究鮮有報道,影響成肌細胞的機理尚不明確。因此,本試驗以關嶺牛成肌細胞作為研究對象,構建篩選MEF2A基因的最佳干擾載體,轉染載體至細胞后,qRT-PCR法檢測成肌細胞中肌生成相關基因的表達;檢測細胞周期與活力特性。為進一步探索MEF2A基因對關嶺牛肌肉發育的分子效應及調控機制提供數據參考。
1 材料與方法
1.1 試劑及儀器
Lipofectamine TM3000CD Reagent脂質體轉染試劑盒購自Thermo Fisher Scientific(美國);DMEM/F12培養基、Gibco澳洲胎牛血清均購自重慶奧怡生物技術有限公司;PBS溶液、青鏈霉素、Opti-MEMTM均購自廣西卓一生物技術有限公司;Trizol試劑購自英駿生物技術有限公司(美國);SYBR Green qPCR Master Mix、逆轉錄試劑盒均購自Genstar公司(北京)。超微量紫外分光光度計購自美國Thermo Fisher公司;酶標儀購自美國Bio Tek公司;實時熒光定量PCR儀(型號為CFX96 Touch)、多功能梯度擴增PCR儀(型號為ABIVeriti-TM)、凝膠成像系統(型號為Uni-versal Hood Ⅱ)均購自美國BIO-RAD有限公司。
1.2 成肌細胞的培養與鑒定
試驗動物來自貴州省安順市西秀區幺鋪鎮屠宰場,選擇健康的3日齡犢牛,采集犢牛背最長肌,生理鹽水(0.9% NaCl)沖洗后放入含雙抗的PBS溶液中保存帶回實驗室。成肌細胞分離培養與鑒定參照宋林錦等[20]的操作方法。率先清理筋膜與雜質,剪碎肌肉組織后加入Ⅱ型膠原酶,放置37 ℃恒溫搖床消化1 h,加入培養基混勻,依次通過滅菌紗布、200目和400目細胞篩,濾液以8 000 r·min-1離心8 min,棄上清液后加入10% DMEM/F-12培養基重懸細胞,放置于37 ℃,5% CO2的細胞培養箱中。利用免疫熒光法鑒定成肌細胞純度,將細胞懸液接種在置有玻璃片的24孔板中,細胞培養箱中培養2 h;細胞爬片后,加入4%多聚甲醛于4 ℃條件下固定35 min,PBS洗滌3次后置于培養皿支撐物上。吸取60 μL破膜封閉液滴于防水膜上,2 h破膜封閉,取60 μL一抗(α-actin)于防水膜上,蓋上玻片于4 ℃放置;避光孵育二抗(goat anti-rabbit lgG)2 h后,PBS洗滌3次,每次4 min,DAPI染色5 min,PBS洗滌3次,每次4 min。加1滴熒光封片劑于玻片上,將有細胞的一面蓋上,用于后續拍照儲存。
1.3 shRNA引物設計與合成
參考NCBI數據庫中的MEF2A基因編碼區序列(GenBank登錄號:NM_001083638.2)與短發夾RNA(short hairpin RNA, shRNA)設計原則,設計MEF2A基因的shRNA片段引物,共設置4個干擾組靶序列和1個NC組序列,分別命名為shRNA-MEF2A-1、shRNA-MEF2A-2、shRNA-MEF2A-3、shRNA-MEF2A-4與shRNA-NC。于序列兩端添加EcoR Ⅰ與SalⅠ酶切位點,序列中段插入Loop (TCAAGAG)結構,序列最后加上6個T,可作為RNA聚合酶Ⅲ的轉錄終止子發揮作用。由上海吉瑪制藥技術有限公司合成片段引物,引物序列信息見表1。

表1 shRNA信息
1.4 干擾載體構建與鑒定
復蘇空載體質粒,利用EcoRⅠ與SalⅠ同時進行酶切,取1.5 μL pGPU6-GFP-Neo、3 μL10×Green Buffer、1.5 μLEcoRⅠ、1.5 μLSalⅠ,加ddH2O補足30 μL,37 ℃條件下酶切2 h,酶切產物用1.2%的瓊脂糖凝膠電泳檢測并切膠回收;將shRNA片段連接到pGPU6-GFP-Neo載體,16 ℃金屬浴過夜連接。連接產物轉化到DH5α感受態細胞中,挑選單克隆菌落擴大培養,37 ℃,200 r·min-1振蕩培養16 h,將菌液送至上海生工生物工程股份有限公司進行測序驗證。
1.5 干擾載體轉染與檢測
成肌細胞接種于6孔細胞培養板中,待細胞生長至80%~90%的融合度時,利用Lipofectamine 3000進行轉染,分別將重組載體pGPU6-GFP-Neo-NC、pGPU6-GFP-Neo-MEF2A-1、pGPU6-GFP-Neo-MEF2A-2、pGPU6-GFP-Neo-MEF2A-3、pGPU6-GFP-Neo-MEF2A-4轉染至成肌細胞內,各孔轉染質粒2 500 ng,空白組不轉染質粒,各轉染組設置3個生物學重復,放置于37 ℃,5% CO2的培養箱中。48 h后將細胞置于熒光倒置顯微鏡下觀察生長狀態并檢測MEF2A基因shRNA干擾效率。
1.6 qRT-PCR檢測基因表達
Trizol法提取細胞總RNA,反轉錄得到cDNA作為模板。依據NCBI中公布的基因序列,利用Primer 5.0設計牛肌細胞增強因子家族基因以及細胞生長相關基因CDK2、BCL2與CCNA2的熒光引物(表2)。qRT-PCR檢測肌細胞增強因子家族基因(MEF2A、MEF2B、MEF2C、MEF2D)以及BCL2、CDK2與CCNA2基因在最佳干擾組和對照組中的表達水平,GAPDH為內參基因。PCR反應體系10 μL:SYBR Green Mix 5 μL,上、下游引物各0.5 μL,cDNA 0.5 μL,ddH2O 3.5 μL。每個樣品做3個生物學重復,反應條件為95 ℃預變性30 s;95 ℃變性5 s,退火5 s,95 ℃延伸5 s,共39個循環。

表2 引物信息
1.7 流式細胞儀檢測細胞周期
將最佳干擾組與對照組載體分別轉染至成肌細胞,用4 ℃預冷的PBS洗滌,加入1 mL 0.25%不含EDTA的胰酶進行消化,1 000 r·min-1離心5 min,棄上清液,加入預冷的1.5 mL的70%乙醇吹打細胞,4 ℃儲存過夜。離心乙醇固定過的細胞,棄上清液;PBS洗滌細胞3次;加入100 μL RNase A重懸細胞,37 ℃水浴鍋中放置30 min,加入500 μL PI染色液,4 ℃避光反應30 min;利用流式細胞儀進行檢測。
1.8 成肌細胞活性檢測
最佳干擾組與對照組載體分別轉染至成肌細胞后,消化計數并接種于96孔細胞板中,各組別設置3個重復,在0、6、12、24、48和72 h時間段內進行檢測,各孔添加10 μL的CCK8試劑,放置于37 ℃,5% CO2培養箱中培養2 h,利用酶標儀(SynerGyH1, BioTek)檢測各組別在450 nm波長處的吸光度。
1.9 關嶺牛MEF2A基因生物信息學分析
利用在線軟件ProtParam(https://web.expasy.org/protparam/)、PSORT II Preadict (https://psort.hgc.jp/form2.html)與STRING(https://string-db.org/)分別對關嶺牛MEF2A的蛋白理化特性、亞細胞結構定位與蛋白網絡圖譜進行預測分析。
1.10 數據分析
采用2-ΔΔCt法計算基因表達量,各組別設置3個生物學重復,數據均用“平均值±標準誤”表示,采用獨立樣本t檢驗和單因素方差分析進行數據處理(SPSS 25.0),多重比較采用Duncan’s多重極差檢驗法,運用Graphpad 6.0作圖。
2 結 果
2.1 關嶺牛成肌細胞培養與鑒定
如圖1所示,固定細胞后,α-actin經異硫氰酸熒光素(fluorescein isothiocyanate, FITC)標記后在細胞質內陽性表達為綠色發光狀態。細胞核為橢圓形狀態,經4′, 6-二脒基-2-苯基吲哚(DAPI)染色后發藍色光。細胞核與細胞質染色重合良好,表明成功培養鑒定關嶺牛成肌細胞,進一步鑒定細胞純度達到90%左右,可開展后續試驗。

圖1 成肌細胞免疫熒光鑒定(200×)
2.2 干擾載體測序
將酶切純化回收的pGPU6-GFP-Neo載體與退火后的shRNA置于16 ℃恒溫金屬浴中過夜連接,連接產物轉化到DH5α感受態細胞中,挑選單克隆菌落進行搖菌培養,篩選陽性菌液送至上海生工生物工程股份有限公司測序驗證。測序結果如圖2所示,插入的重組質粒片段與設計合成的序列一致,說明成功構建了重組質粒。
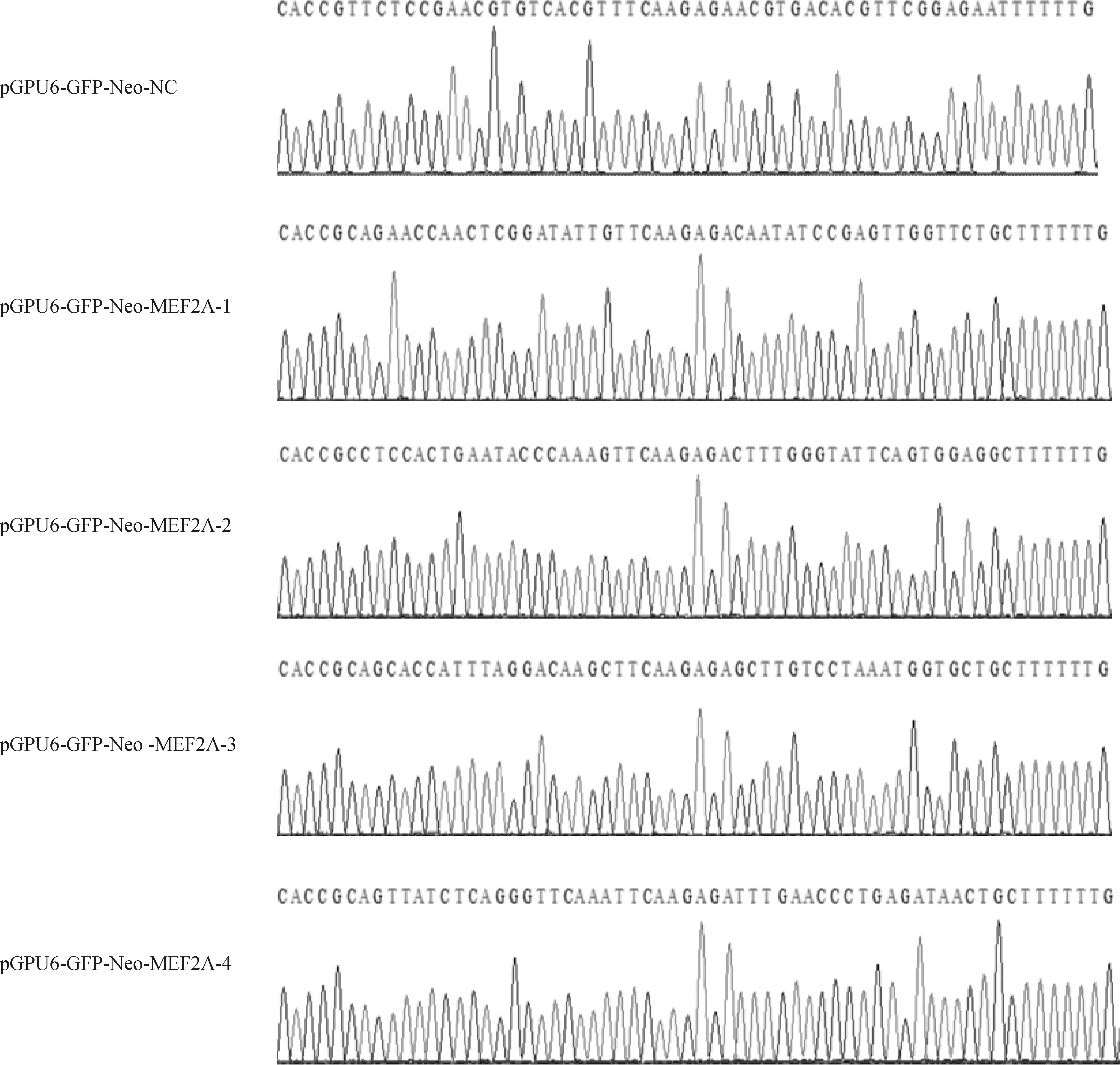
圖2 重組質粒測序分析結果
2.3 干擾載體轉染
將成肌細胞接種到6孔板中,細胞融合度達到90%左右時,將各干擾組與陰性對照組轉染至成肌細胞,放置于細胞培養箱中48 h后用熒光倒置顯微鏡觀察各組別綠色熒光蛋白的發光情況。結果如圖3所示,在熒光倒置顯微鏡下的各干擾組、陰性對照組均有清晰明亮的綠色熒光,其中空白組不發光,說明轉染成功。
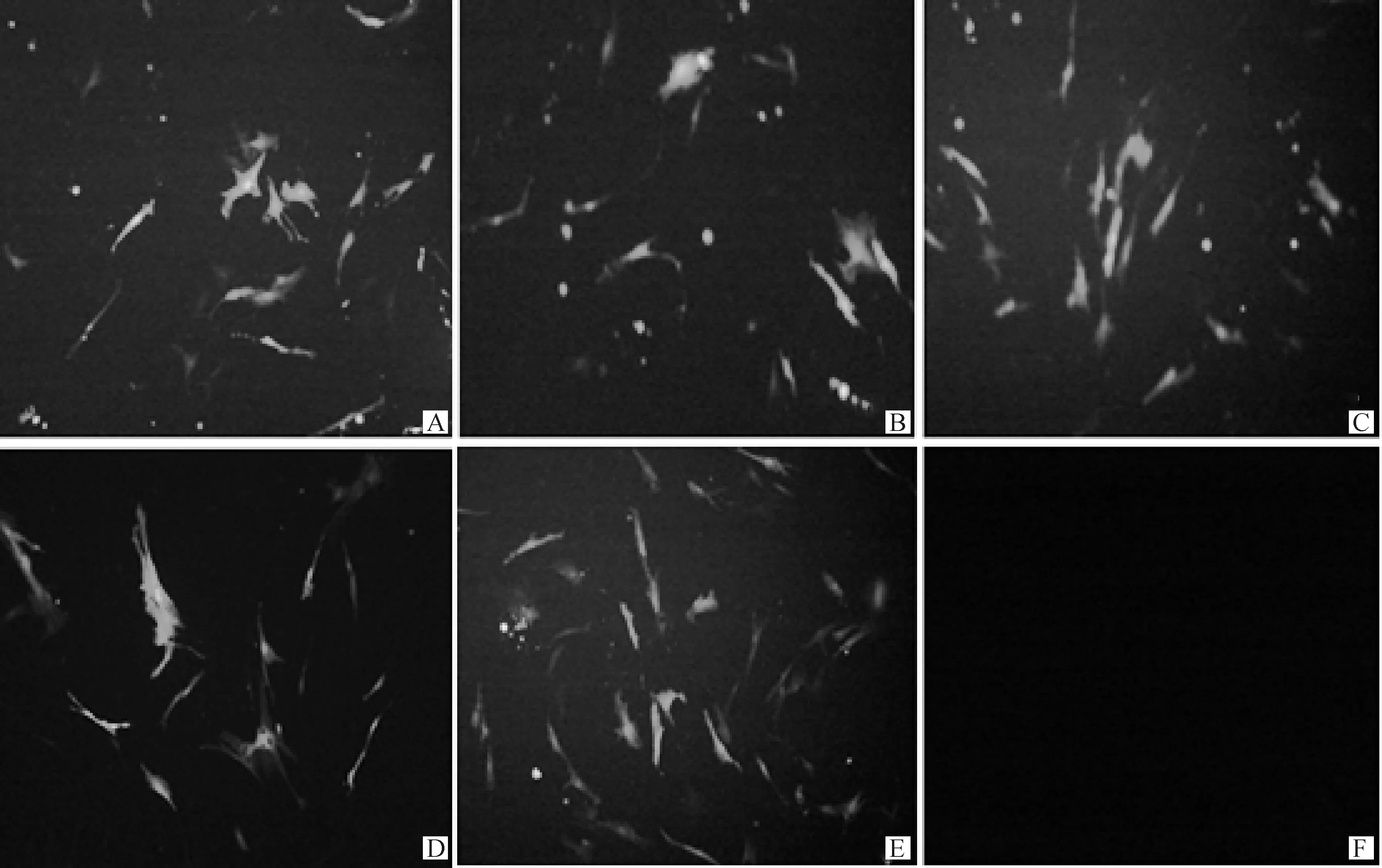
A. shRNA-MEF2A-1;B. shRNA-MEF2A-2;C. shRNA-MEF2A-3;D. shRNA-MEF2A-4;E. shRNA-NC;F. 空白對照
2.4 qRT-PCR檢測干擾載體效率
將各重組質粒分別轉染至牛成肌細胞48 h,提取細胞RNA并逆轉錄為cDNA,qRT-PCR法檢測MEF2A基因在不同組別中的表達量特性,篩選干擾效果最佳的試驗組別。如圖4所示,shRNA-MEF2A-1、shRNA-MEF2A-2與shRNA-MEF2A-3和shRNA-MEF2A-4干擾效率依次為74.45%、58.67%、94.66%與62.98%。結果表明,4個干擾載體均能極顯著下調MEF2A基因表達量(P<0.01),選用干擾效率最高的shRNA-MEF2A-3干擾載體開展后續試驗。

*. 表示差異顯著(P<0.05);**. 表示差異極顯著(P<0.01),下同
2.5 干擾MEF2A對成肌細胞肌生成等相關基因表達的影響
分別提取轉染shRNA-MEF2A-3最佳干擾組和shRNA-NC對照組的細胞總RNA,對肌生成基因及周期與凋亡相關基因表達量進行qRT-PCR檢測。結果如圖5所示,干擾MEF2A基因表達后,肌細胞增強因子MEF2B、MEF2C與MEF2D基因表達量均極顯著上調(P<0.01);細胞周期因子CDK2表達量顯著下調(P<0.05),CCNA2表達量極顯著下調(P<0.01);抗凋亡因子BCL2表達量顯著下調(P<0.05)。推斷MEF2A基因在關嶺牛成肌細胞中作用顯著,與家族其他成員具有相互調控關系;干擾MEF2A可延長關嶺牛成肌細胞周期,對其增殖具有抑制作用。

圖5 干擾MEF2A對成肌細胞基因表達的影響
2.6 流式細胞儀檢測細胞周期
利用流式細胞儀檢測轉染shRNA-MEF2A-3干擾組和shRNA-NC對照組的細胞周期特性。結果如圖6所示,與shRNA-NC相比,轉染shRNA-MEF2A-3的成肌細胞數量在G1期顯著增多(P<0.05),S期與G2/M期細胞數量極顯著減少(P<0.01)。研究結果表明,干擾MEF2A后,細胞周期G1至S期的進程被阻滯,細胞增殖受到抑制。
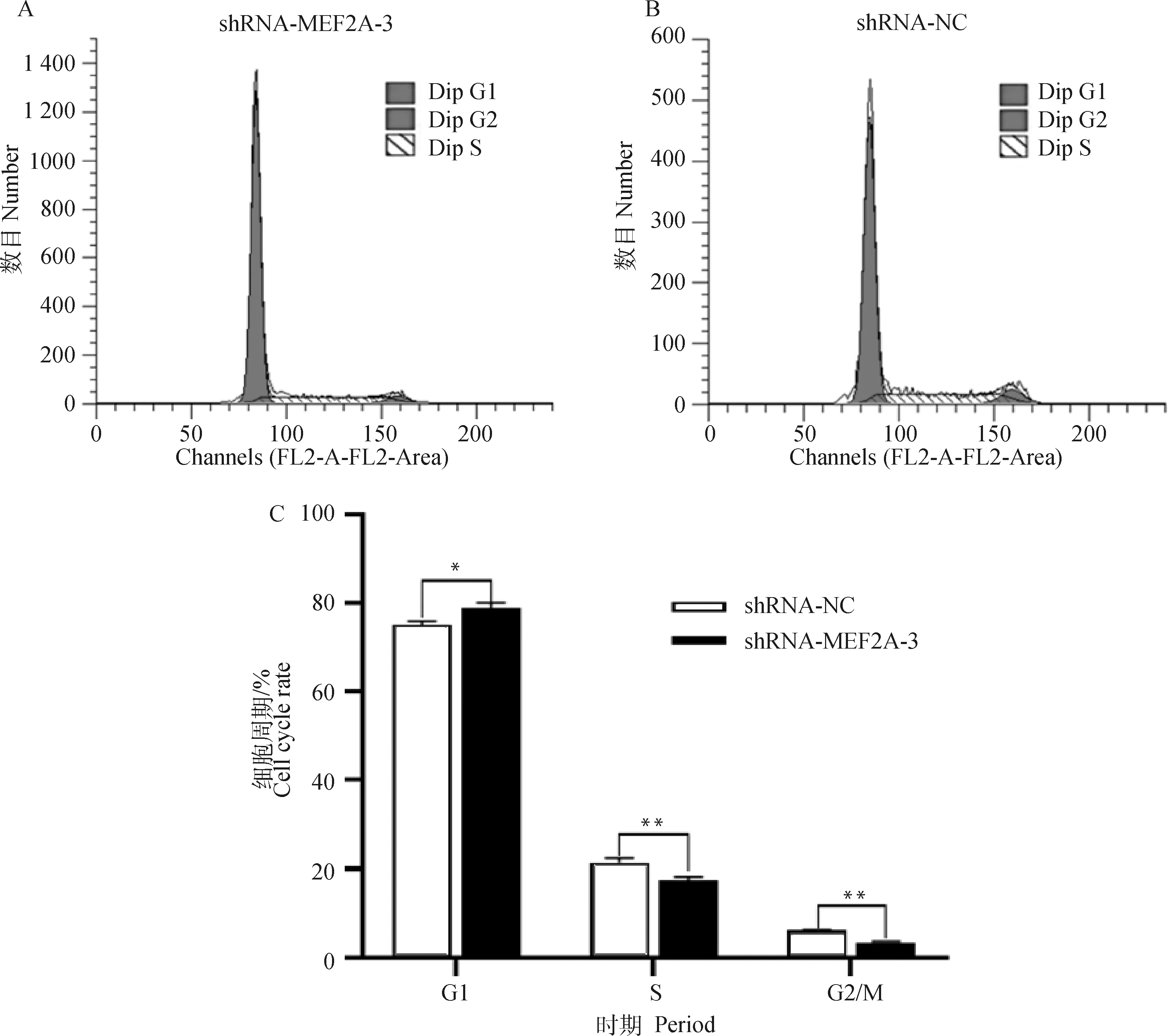
A. 干擾組細胞周期;B. NC組細胞周期;C. 細胞周期比例
2.7 關嶺牛成肌細胞生長檢測
分別在0、6、12、24、48和72 h檢測shRNA-MEF2A-3干擾組和shRNA-NC對照組的細胞生長增殖特性。CCK8檢測結果如圖7所示,自6 h后,shRNA-MEF2A-3干擾組和shRNA-NC對照組在5個不同時間段內的細胞生長情況均達到極顯著差異水平(P<0.01)。
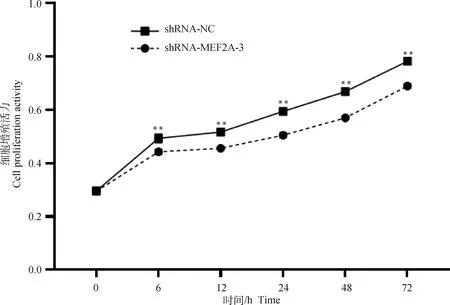
圖7 干擾MEF2A對成肌細胞增殖的影響
2.8 牛MEF2A蛋白理化性質分析
在線軟件預測MEF2A(No.:NM_001083638.2)基因的mRNA序列為5 568 bp,編碼492個氨基酸。MEF2A蛋白分子式為C2282H3649N659O737S21,相對分子質量約為52 782.31 u;絲氨酸(13.8%)在氨基酸中組成含量最高,色氨酸(0.4%)組成含量最低;帶正電荷的氨基酸殘基數量(精氨酸+賴氨酸)為45個;帶負電荷的氨基酸殘基數量(天冬氨酸+谷氨酸)為39個。亞細胞定位顯示,MEF2A主要位于細胞核(60.9%)。STRING預測牛MEF2A蛋白互作網絡圖譜,結果如圖8所示,包含組蛋白去乙酰化酶(HDAC)家族蛋白成員HDAC4、HDAC5與HDAC7,參與核心組蛋白(H2A、H2B、H3和H4)N末端部分賴氨酸殘基的去乙酰化[21];絲裂原蛋白活化激酶(MAPK)家族蛋白成員MAPK11、MAPK14,可通過激活環中TGY基序的雙重磷酸化發揮功效[22],調節細胞分化與生長凋亡,在細胞反應級聯中起重要作用,以及影響心肌發育和成肌細胞分化的MEF2C、EP300、MEF2D、MYOD1、NKX2-5等相關蛋白。
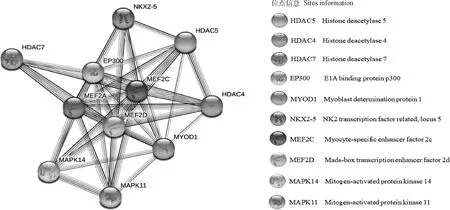
圖8 牛MEF2A蛋白互作網絡預測分析
3 討 論
MEF2本身不具備肌源性活性,當與bHLH(basic helix-loop-helix)蛋白家族互作時,可驅動并放大肌源性分化程序[23]。MEF2蛋白N端具備結合DNA功能的MADS區域和中心MEF2結構域,C端為轉錄激活區域,該區域包含大量的核定位序列和磷酸化位點,在MEF2基因家族成員及不同物種間的保守性較低且序列變化多樣,與基因轉錄活性關系密切,可進一步調控MEF2蛋白活性[24-25]。MEF2主要通過CaMK-HDACs(Ca2+-鈣調素依賴的蛋白激酶-組蛋白去乙酰化酶)、Calcineurin(鈣調磷酸酶)和MAPK(絲裂原活化蛋白激酶)3種信號通路發揮作用[26]。MEF2基因大多通過MADS-box和MEF2結構域與其他轉錄輔因子協同作用,家族成員與許多參與肌生成的基因相結合,以啟動或加強肌源性基因的表達[27-28],通過控制下游靶基因的轉錄活性來調控細胞的分裂、分化和凋亡過程。目前已證實,牛MEF2A基因被定位于21號染色體[29],是一種廣譜表達基因,在肌肉發育過程中,該基因表達具有發育時空性與組織特異性。MEF2A基因的多態性對畜禽屠宰性能存在一定的影響,且該基因為影響畜禽屠宰性狀的候選基因之一[30]。Liu等[31]研究發現,同時敲除MEF2A、MEF2C和MEF2D后,小鼠肌損傷修復過程受到顯著抑制,成肌細胞增殖受到嚴重阻礙。而本試驗在抑制MEF2A基因表達后,關嶺牛成肌細胞生長活性下降,細胞增殖同樣受到嚴重阻礙。同時,有研究表明MEF2A、MEF2C表達水平在蛙類動物冬眠期具有重要作用[32]。
MEF2A作為一種較為保守的轉錄因子,廣泛分布于各種組織或器官中,在包括決定細胞命運、遷移和形狀在內的多種生物學過程中發揮關鍵作用[33]。MEF2A是肌生成的關鍵調節因子和肌再生的重要激活劑,也可參與脂質代謝途徑[34]。Wang等[35]研究發現,MEF2A下游的MEG3-DIO3 miRNA在牛成肌細胞可調節PP2A信號;MEF2A-MEG3/DIO3-PP2A軸在肌再生過程中發揮重要作用,為骨骼肌的發育機制研究提供了新的線索。Hennebry等[36]研究報道,MSTN可通過調控MEF2與MyoD基因家族表達進而調節骨骼肌纖維類型組成;在轉基因小鼠中表達過活性形式的MEF2可促進慢肌纖維形成,但MEF2的失活會導致慢肌纖維形成的喪失[37]。Schiaffino與Serrano[38]研究發現,細胞內Ca2+濃度較低時,HDAC、HDAC5、HDAC7與MEF2蛋白N末端的MADS/MEF2結構域相互作用而抑制MEF2的活性;當Ca2+濃度升高時,Ca2+與鈣調蛋白結合并激活CaMK,其中CaMKIV發揮磷酸化作用使HDACs失去活性,MEF2從HDACs上脫離,與本研究中預測的MEF2A蛋白網絡圖譜結果相吻合。此外,有研究表明MEF2A在抗炎癥和治療癲癇等方面有著重要作用[39-41]。
本試驗初步分析了關嶺牛MEF2A基因的蛋白理化性質,利用qRT-PCR法檢測發現抑制MEF2A基因表達后,肌細胞增強因子MEF2B、MEF2C與MEF2D的表達量均極顯著高于shRNA-NC對照組,進一步推測該家族成員具有協同作用的特性,且MEF2A基因在成肌細胞中的作用比家族其它成員更加顯著。CDK2作為細胞周期蛋白依賴性激酶家族中的一員,與細胞周期蛋白CCNA2相互作用可調控細胞周期進程[42]。MEF2A基因被抑制后,相較于shRNA-NC對照組,成肌細胞中CDK2與CCNA2基因表達量皆降低;流式細胞結果顯示,G1期細胞數量顯著增多,S期細胞數量極顯著減少,表明細胞周期進程受到阻滯。同時發現干擾MEF2A基因可導致細胞抗凋亡因子BCL2基因表達量下降,進一步研究發現轉染干擾組的成肌細胞自6 h后的細胞增殖能力相較于對照組下降明顯。由此推斷,干擾MEF2A基因導致成肌細胞周期延長,細胞增殖受到抑制,與Wang等[43]的研究結果相似。然而,MEF2A影響成肌細胞的具體分子機制還需要更進一步的探究。
4 結 論
本研究成功構建了關嶺牛MEF2A干擾載體,干擾MEF2A基因明顯抑制關嶺牛成肌細胞的增殖。同時推測,MEF2A通過調控成肌細胞中肌細胞增強因子基因和相關生長凋亡基因的表達來影響肌肉發育性能,為進一步完善MEF2A基因對關嶺牛生長發育的影響和探究地方優質種質資源提供了理論參考。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
