
暗色絲孢菌中2個中國新記錄種和1個中國大陸新記錄種
2023-02-27 03:08:48肖仲久錢正敏李小霞陸錚錚宋培勇陳模芳
熱帶作物學報 2023年1期
肖仲久,劉 婷,錢正敏,李小霞,楊 澤,陸錚錚,宋培勇,陳模芳
1.遵義師范學院資源與環境學院,貴州遵義 563002;2.遵義師范學院生物與農業科技學院(食品科技學院),貴州遵義563002;3.貴州省赤水河流域植物資源保護與應用特色重點實驗室,貴州遵義 563002
Beltraniella由 SUBRAMANIAN[1]在馬德拉斯(印度港市)從枯死的厚皮樹Odina wodierRoxb.[≡Lannea coromandelica(Houtt.) Merr.]的葉片上獲得菌株,以B.odinaeSubram.為模式種建立的。目前該屬在 Index Fungorum中共有 29個種被記錄,其中B.porosaPiroz.& S.D.Patil [≡Porobeltraniella porosa(Piroz.& S.D.Patil) Gusm?o]和B.patiliiK.G.Karand.& Patw.[≡Porobeltraniella patilii(K.G.Karand.& Patw.)Gusm?o]2個種的分生孢子最寬處有圓形孔帶,因此 FERNANDO等[2]將這 2個種移入PorobeltraniellaGusm?o中,現今Belraniella有效種共 27個。該屬典型特征如下:剛毛狀分生孢子梗的隔膜處易產生側向分枝,分枝可進一步分支形成產孢細胞或分離細胞,其上以頂側生方式產生鼻甲或倒圓錐形,0個隔膜,光滑,半透明至淺褐色,具明顯透明橫帶的分生孢子。該屬產孢方式及分生孢子的形狀與BeltraniaPenz.、BeltraniopsisBat.& J.L.和Porobeltraniella相似,Beltrania的分生孢子梗產生于剛毛的基部細胞或單獨的放射狀淺色細胞,分生孢子雙錐形,頂端常具透明絲狀附屬物;Beltraniopsis不分枝或分枝的剛毛狀分生孢梗產生不對稱雙錐形且頂端具短喙的分生孢子;Porobeltraniella不育的側剛毛上產生產孢裝置(產孢細胞),其上產生具透明、多個赤道孔的分生孢子[2-4]。
Subulispora由TUBAKI等[5]在日本的長尾栲(Castanopsis cuspidataSchottky var.sieboldii)獲得菌株,以S.procurvataTubaki作為模式種建立,隨后在瓜達爾卡納島、坦桑尼亞、肯尼亞、印度、巴西和中國臺灣等有報道。目前該屬在 Index Fungorum中有16個種,其中S.hareaeB.Sutton因分生孢子鏈生,KIRK[6]將S.hareae[≡Polyscytalum hareae(B.Sutton) P.M.Kirk]移入PolyscytalumRiess中;而S.gracile(Matsush.) de Hoog和S.variabilede Hoog分生孢子梗的薄壁、透明,CASTA?EDA 等[7]將這 2個種移入CylindrosympodiumW.B.Kendr.& R.F.Casta?eda中,故該屬目前共有13個有效種。該屬典型特征如下:粗大,褐色,曲膝延伸,具明顯鋸齒形的分生孢子梗頂生單生,蒼白色,錐形或圓柱形,直或彎曲,基部平截,具0或多個隔膜的分生孢子。Subulispora在產孢方式及分生孢子形狀上與Cylindrosympodium、Polyscytalum、MirandinaG.Arnaud ex Matsush.和ParasympodiellaPonnappa四個屬相似,其中Cylindrosympodium分生孢子梗半透明至透明;Polyscytalum分生孢子梗合軸式延伸,但具不明顯鋸齒,分生孢子鏈生;Mirandina分生孢子梗頂部區域膨大、聚集圓柱形小齒,分生孢子紡錘形;Parasympodiella分生孢子梗以體生式方式產生鏈生分生孢子。
Beltraniella和Subulispora隸屬于暗色絲孢菌(dematiaceous hyphomycetes),該類群真菌在自然生態系統中對木質素、纖維素降解,物質和能量循環轉化及在維持生態系統平衡中具有重要的生態學作用[8-10]。因此,目前深入系統地開展暗色絲孢真菌的鑒定,對豐富我國該類真菌資源及后續對該類真菌開展降解有機物、維持生態系統平衡等研究均具有重要的支撐意義。
1 材料與方法
1.1 材料
2018—2019年7 —8月、10—12月,采集貴州省習水國家級自然保護區和貴州省寬闊水國家級自然保護區的不同生境下的凋落枯枝落葉(腐爛適中),標注采集地、時間、基質(可辨認)、海拔和經緯度等。
1.2 方法
將采集的凋落枯枝裁剪成6~7 cm小段,置入鋪有濕潤吸水紙的塑料培養皿內(落葉可整片保濕在磁盤內),不定期保濕,25℃培養,以備鏡檢。將上述凋落枯枝落葉置于SMZ 745T體視顯微鏡下,用細挑針挑取菌株制成臨時玻片,在Eclipse E200生物顯微鏡下選取合格的臨時玻片制成永久玻片;隨后在 ECLIPSE Ni微分干涉顯微鏡和SMZ745T體視顯微鏡下分別根據菌株的形態大小和基質上菌落大小在相應的倍數下拍照,用Image-Pro Plus 5.0軟件和Adobe Photoshop CS6進行測量及組圖,通過查閱相關資料,結合菌株形態特征及相關測量數據進行屬、種的形態學鑒定。
將分離成功的菌株進行基因組 DNA提取[真菌基因組 DNA提取試劑盒,天根生化科技(北京)有限公司],選擇引物 ITS4/ITS5和28S1/28S3[11];使用 50 μL的 PCR反應體系,包括正反向引物各 2 μL,DNA 模板 2 μL,2×PCR Master Mix 25 μL,ddH2O 19 μL。擴增 ITS 的反應條件為:94℃預變性5 min;94℃變性30 s,57℃退火50 s,72℃延伸90 s,35個循環,72℃延伸10 min。將獲得的DNA溶液置于-20℃冰箱保存備用。
測序工作由北京諾賽基因組研究中心有限公司完成。序列剪接使用Chromas軟件,序列對比使用 NCBI中的Blast工具。系統發育樹的構建采用Mega X的ML(maxium likelihood)法,自展重復次數設置為 1000。參與分析的序列包括自測序列(其DNA序列均提交至GenBank)和GenBank中下載的相似屬和同屬相似種的序列(表1和表2)。
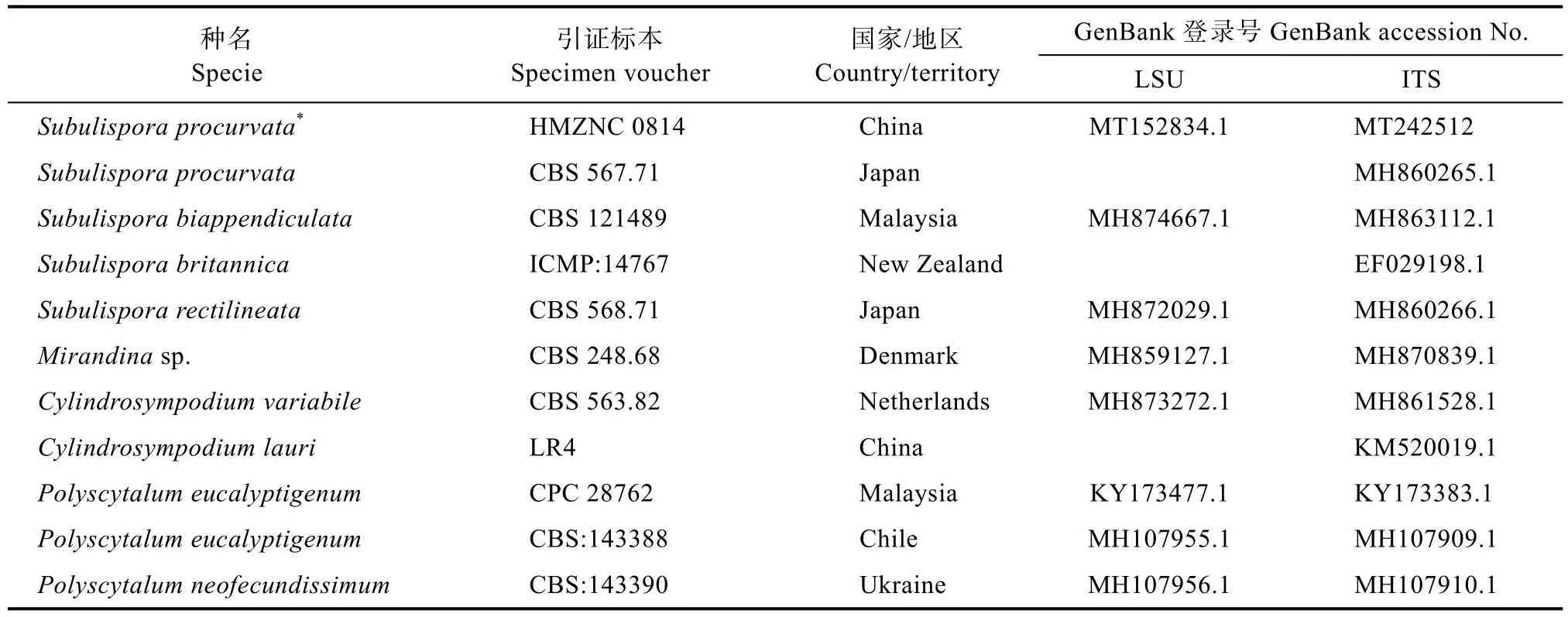
表1 用于構建系統進化樹的菌株及GenBank登錄號Tab.1 Strains used in phylogenetic analyses and their corresponding GenBank accession numbers
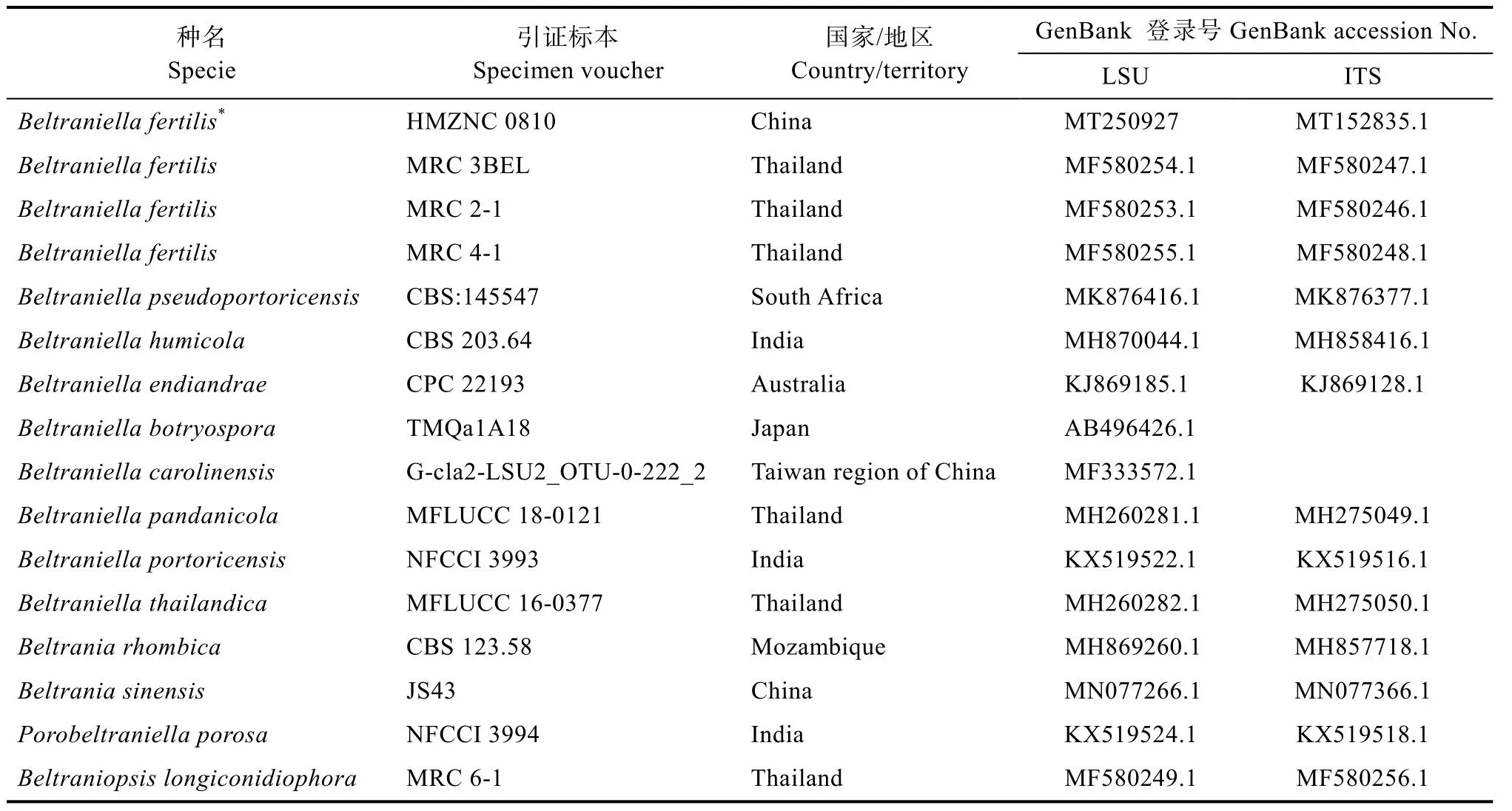
表2 用于構建系統進化樹的菌株及GenBank登錄號Tab.2 Strains used in phylogenetic analyses and their corresponding GenBank accession numbers
2 結果與分析
2.1 可育小貝氏孢(Beltraniella fertilis Heredia, R.M.Arias, M.Reyes & R.F.Casta?eda)
在腐爛的枯葉上腐生。有性態:未發現。無性態:菌落腐生在葉片的兩面,舒展,薄壁,灰褐色;在PCA培養基上,菌落最初白色,隨后變為灰色至灰褐色。菌絲部分埋生在基質內,由具隔、分枝、壁光滑、半透明至淺褐色,寬1.5~2 μm的菌絲體組成。剛毛簡單,產生于放射狀、淺裂的基部細胞,寬6.5~15.5 μm,直立,直或稍彎曲,單生或少部分聚集,黑褐色,薄壁,0~3個隔膜,長57~125 μm,基部和頂部光滑,中下部具有稀疏的疣狀突起,中下部寬4~5 μm,頂部逐漸變細,淺褐色。分生孢子梗粗大,單生,具長的剛毛狀和短的非剛毛狀2種類型:長的剛毛狀的分生孢子梗產生于放射狀、淺裂的基部細胞,單生或小數聚生,直,7~12個隔膜,大小156~392 μm×4.5~8 μm,基部和頂部光滑,中下部具有稀疏的疣狀突起,厚壁,頂部區域分枝;短的非剛毛狀分生孢子梗分枝,0~2個隔膜,壁光滑,半透明至透明,薄壁,6~12 μm×3.5~5.5 μm。產孢細胞多芽生,合生,端生,棍棒狀至圓柱形,頂端或亞頂端具小齒,淺褐色,6~12 μm×3~5 μm。分離細胞卵圓形或倒卵形,薄壁,光滑,透明,6.5~11 μm×3.5~5.5 μm,兩端各具一個小齒。分生孢子直接產生與產孢細胞上或產生于分離細胞上,遠極端平截,近極端具喙狀凸起,光滑,赤道帶以上具半透明至淺橄欖色的橫帶,17~23 μm×5~7 μm(圖1)。

圖1 可育小貝氏孢(HMZNC0810)Fig.1 Beltraniella fertilis (HMZNC0810)
研究標本:貴州省習水國家級自然保護區,28°31′2.04″N,106°23′56.05″E,海拔 1033.9 m;基質為廣玉蘭(Magnolia GrandifloraL.)凋落枯葉;2019年7月25日;標本號為HMZNC0810。
Beltraniella所記錄的27個有效種中與菌株相似的種為B.fertilisHeredia, R.M.Arias, M.Reyes& R.F.Casta?eda、B.ramosiphoraC.G.Lin & K.D.Hyde和B.pseudoportoricensisCrous,其中B.ramosiphora的長分生孢子梗不分枝,且短的分生孢子梗較長(20~55 μm)[12],B.pseudoportoricensis的分生孢子較長些(25~27 μm)[13];而B.fertilis與本菌株相比分生孢子的長度(19~22 μm vs 17~23 μm)基本一致[14],同時在剛毛、2種類型的分生孢子梗、分離細胞上也基部一致。從圖2可知,各菌株明顯分為2支,Beltraniella所供試菌株均在一個亞枝上,菌株(HMZNC 0810)與B.fertilis的親緣關系最近,與Beltraniella的其他種能明顯分開;另外,菌株在形態與相似屬種也存在較大差異。結合形態特征和系統學分析,將菌株(HMZNC 0810)定為B.fertilis。B.fertilis在墨西哥韋拉克魯斯、泰國、巴西有報道,但在中國屬于首次報道,故將其定為中國新記錄種。

圖2 基于LSU和ITS序列對可育小貝氏孢構建的進化樹Fig.2 Phylogentic tree of Beltraniella fertilis generated based on combined LSU and ITS sequences
2.2 柱孢錐孢(Subulispora cylindrospora P.M.Kirk)
在枯枝上腐生。有性態:未發現。無性態:菌落舒展,薄壁,絨毛狀,灰褐色。菌絲部分埋生在基質內,由光滑、分枝、具隔、淺褐色菌絲體組成。分生孢子梗粗大,單生,直立,簡單,直或稍彎曲,上部具明顯的膝曲狀,呈“之”字形,褐色,頂部淺色,光滑,2~5個隔膜,大小33~78 μm×2.5~4.3 μm,基部寬 3~5.7 μm。產孢細胞合生,端生,多芽生,合軸式延伸,具明顯的產孢痕。分生孢子單生,干質,頂側生,光滑,透明,圓柱形,稍錐型,頂部鈍圓,基部平截,并具一個增厚、稍顯色素的疤痕,(2-) 5~6 (-8)個真隔膜,大小 34.7~77 μm×2~3 μm(圖3)。

圖3 柱孢錐孢(HMZNC0813)Fig.3 Subulispora cylindrospora (HMZNC0813)
研究標本:貴州省寬闊水國家級自然保護區,28°02′3.2″N,107°10′51.24″E;海拔 1103 m;基質為未鑒定枯枝;2019年 7月 26日;標本號為HMZNC0813。
Subulispora所記錄的13個有效種中,分生孢子為圓柱形且頂端無附屬物的種為S.minimaP.M.Kirk、S.africanaP.M.Kirk和S.cylindrosporaP.M.Kirk。S.minima的分生孢子梗較短(4~15 μm)且透明;S.africana的分生孢子隔膜數較少(3~4個),且較短(31~33 μm);S.cylindrospora在分生孢子梗長度(24~68 μm)、分生孢子隔膜數(5個)及大小(37~56 μm)(Kirk 1985)與菌株基本一致[15-17]。HMZNC 0813菌株在PDA、PCA等不同培養基質上進行單孢分離,通過平板劃線分離法進行分離純化,可能由于自然基質上存在過多的雜菌、目的菌株數量較少、不集中、目的菌株上表面附帶其他菌類等原因,最終未獲得純菌種,但通過形態學特點及測量數據,可將其明確定為S.cylindrospora。S.cylindrospora在國外肯尼亞有報道,但在中國未見報道,因此將該菌株定為中國新記錄種。
2.3 前彎錐孢(Subulispora procurvata Tubaki)
在枯枝上腐生。有性態:未發現。無性態:菌落生長在葉子的兩面,舒展,薄壁,絨毛狀,灰褐色;在PCA培養基上最初白色,后期灰色至灰褐色,而PCA的反面深褐色。菌絲部分埋生在基質內,由光滑、分枝、具隔、淺褐色,寬2~2.5 μm的菌絲體組成。分生孢子梗粗大,單生或少數聚生,直立或稍彎曲,上部明顯的膝曲彎曲,呈“之”字形,褐色,頂端淺色,光滑,具長和短2種類型:長的分生孢子梗 4~6個隔膜,大小 72~145(-179)μm×3~5 μm,產生于放射狀淺裂的基部細胞,寬9.3~15 μm;短的分生孢子梗0~2 (-3)的隔膜,42.5~52.5 μm×3~5 μm,產生于放射狀淺裂的基部細胞,褐色,寬 5~7 μm。產孢細胞合生,端生,多芽生,合軸式延伸,具明顯的產孢痕。分生孢子單生,干質,頂側生,光滑,透明,錐形,0~3個隔膜,長25~33.5 μm(不包括附屬喙),平截的基部寬 2.2~3.5 μm,基部彎曲的頂端延伸形成一個細長的附屬喙,6~11.6 μm×0.8~1.1 μm(圖4)。
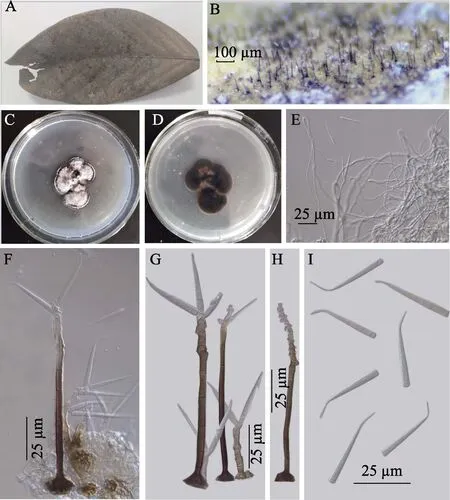
圖4 前彎錐孢(HMZNC0814)Fig.4 Subulispora procurvata (HMZNC0814)
研究標本:貴州省寬闊水國家級自然保護區,28°15′2.25″N,107°11′18.70″E,海拔 1314.5 m;基質為未鑒定的凋落枯葉;2019年7月26日;標本號為HMZNC0814。
Subulispora所記錄的 13個種中,在形態上分生孢子錐形、頂端不分枝且具一定程度彎曲的附屬絲的種為S.procurvataTubaki和S.longirostrataNawawi & Kuthub。S.longirostrata的分生孢子具較長的附屬絲(15~25 μm),在附屬絲的拐點上具弧形缺口[18];S.procurvata的分生孢子梗長度(20~190 μm)、分生孢子的隔膜數(3個)、分生孢子大小(28~40 μm)(不包含附著絲)等[18]特征基本與HMZNC 0814菌株一致,但模式種S.procurvata未提到有2種類型的分生孢子梗,本菌株形態上具明顯的2種類型的分生孢子。從圖5可知,各菌株明顯分為2支,Subulispora所供試的菌株均聚在一個亞分枝上,其中HMZNC 0814菌株和S.procurvata以94%進化支持率聚為一分支,親緣關系最近,與Subulispora屬的其他種能明顯分開。結合形態特征和系統學分析,將HMZNC 0814菌株定為S.procurvata。S.procurvata在日本、巴西、澳大利亞、中國香港等有報道過,故將其定為大陸新記錄種。
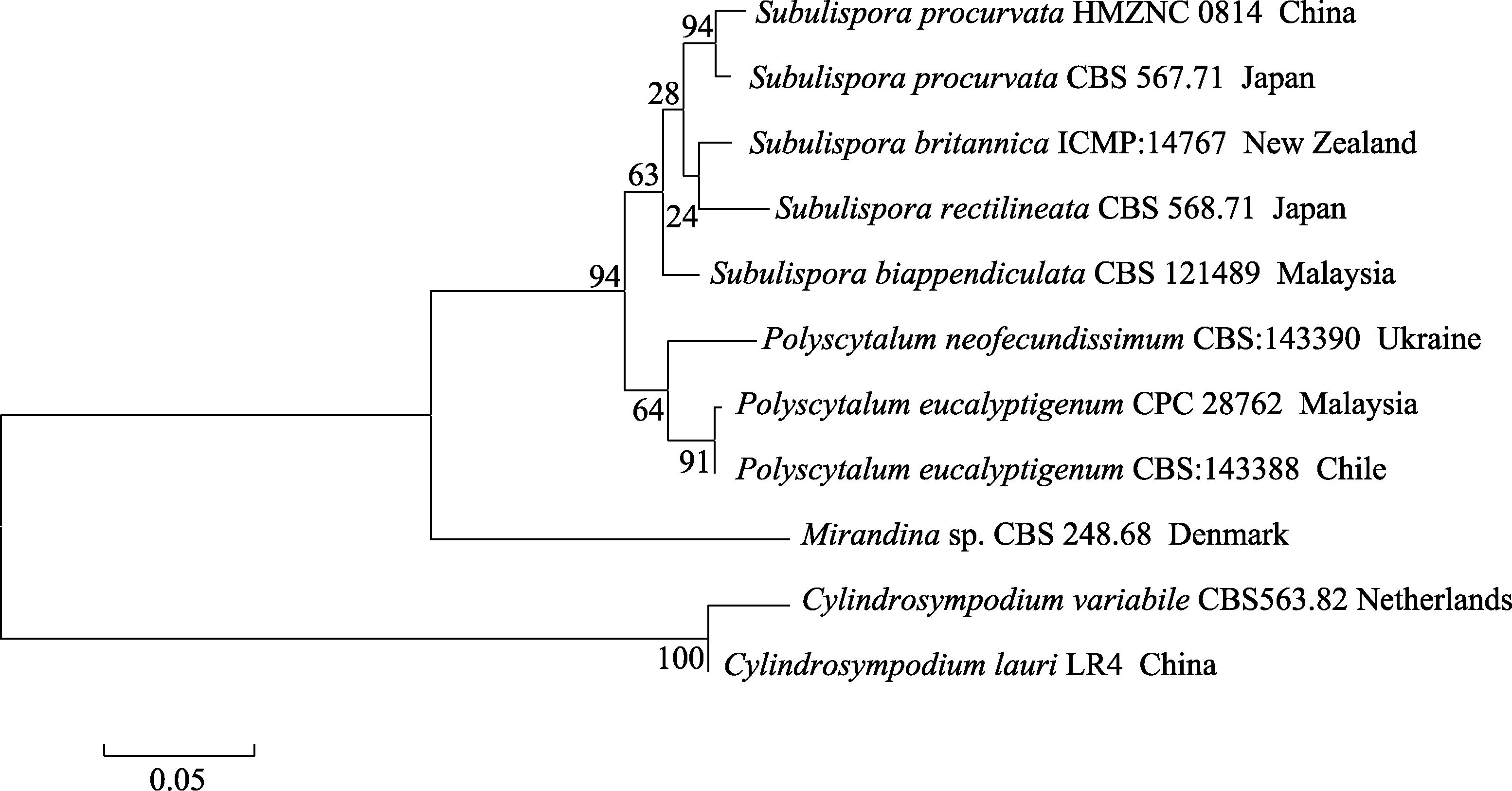
圖5 基于LSU和ITS序列對前彎錐孢構建的進化樹Fig.5 Phylogentic tree of Subulispora procurvata generated based on combined LSUand ITS sequences
