外源ASA、H2O2對牡丹切花衰老過程中生理活性及細胞學特性的影響
2023-03-06 07:09:30邱永杰何松林張翔宇胡緩史來琨于嘉倫賈文慶
山東農(nóng)業(yè)科學 2023年1期
關(guān)鍵詞:植物
邱永杰何松林張翔宇胡緩史來琨于嘉倫賈文慶
(河南科技學院園藝園林學院,河南 新鄉(xiāng) 453003)
牡丹(Paeonia suffruticosa)是芍藥科(Paeoniaceae)芍藥屬(Paeonia)落葉灌木,因其華麗的姿態(tài)和美好的寓意,千百年來深受人們喜愛。近年來,隨著人們對牡丹觀賞價值的深入開發(fā),其作為鮮切花的潛在價值被逐漸挖掘。然而,與月季、菊花、香石竹等其它暢銷切花相比,牡丹花期短、花瓣脆弱及不易運輸、難以持續(xù)保存的特點導致其在鮮切花市場上的流通受到限制,從而制約牡丹鮮切花產(chǎn)業(yè)發(fā)展[1,2]。大量研究表明有多種因素參與牡丹開花和衰老過程的調(diào)控,包括內(nèi)源激素、膜質(zhì)過氧化、大分子物質(zhì)、水分代謝、呼吸代謝等[3]。近年來,通過施加外源保鮮劑延緩切花的衰老進程已經(jīng)成為普遍的切花保鮮方式[4],國內(nèi)關(guān)于此類化學保鮮的研究多集中于保鮮液的成分。研究表明,以蔗糖為主要保鮮劑的配方對香石竹切花有良好的保鮮效果[5];赤霉素能有效延長蝴蝶蘭、百合切花的保鮮時間[6,7];多胺、多效唑、比久可以明顯延緩牡丹切花的壽命[8,9]。
植物花器官的開放和衰老包括花朵吸水擴展生長到失水萎蔫的過程,此過程中植物體內(nèi)發(fā)生一系列生理學和細胞學上的變化。這些變化反映到表型特征上則表現(xiàn)為花朵直徑大小、花色暗淡程度、花瓣褶皺程度、莖稈彎曲程度的變化等。在細胞和亞細胞水平上,研究發(fā)現(xiàn),衰老的花瓣表皮細胞表現(xiàn)出液泡膜內(nèi)陷、細胞壁變形且核糖體減少、DNA斷裂,包括蛋白質(zhì)、核酸、白色體、溶酶體等在內(nèi)的細胞內(nèi)組分發(fā)生自溶等,而關(guān)于細胞核形態(tài)變化的研究尚未見報道。植物的生命活動廣泛存在細胞程序性死亡(PCD)。花器官的衰老作為植物衰老的一部分,是植物生長發(fā)育的一個階段,也是一個細胞程序性死亡的過程。在這個過程中,植物體大量積累各種生理代謝的副產(chǎn)物,其中活性氧(reactive oxygen species,ROS)是主要的代謝產(chǎn)物,主要包括過氧化氫(H2O2)、超氧陰離子()、羥自由基(·OH)、單線態(tài)氧(1O2)等,這些部分被還原或活化氧的衍生物,具有高活性和毒性,可以氧化破壞細胞,導致可溶性蛋白降解、膜脂過氧化程度升高、游離氨基酸積累以及多種酶活性變化等[10,11]。過量的ROS會觸發(fā)植物體內(nèi)抗氧化清除機制,引起酶清除系統(tǒng)中的SOD、POD、CAT等保護酶活性發(fā)生變化[12],因此可以作為判斷細胞PCD程度的關(guān)鍵性指標。
抗壞血酸(ASA)是植物體內(nèi)重要的非酶類小分子抗氧化劑,能與谷胱甘肽(GSH)形成循環(huán)系統(tǒng),在清除植物體內(nèi)抗氧化酶不能清除的O2·-和·OH等自由基、阻止或減輕氧化傷害方面發(fā)揮重大作用[13,14]。陳嬌等[15]研究指出,香蕉表面噴灑外源ASA推遲果實成熟衰老時間;楊慶賀等[13]指出施加外源ASA能提高菊花對低溫弱光的耐性。H2O2是一類ROS,會對植物細胞造成毒害作用,此外研究表明,H2O2還能作為一種信號分子參與信號轉(zhuǎn)導,從而調(diào)節(jié)植物代謝、提高植物的抗逆性[16]。朱利君等[17]研究表明,高鹽脅迫下外源H2O2介導抗氧化酶、ABA和GA可促進黃瓜種子的萌發(fā);蔣景龍等[18]研究指出,施加適宜濃度的外源H2O2可緩解低溫脅迫下大紅柑葉片的卷曲萎蔫,降低低溫對柑橘葉片細胞膜的傷害。目前,關(guān)于施加外源ASA和H2O2對牡丹切花保鮮影響的研究尚未見報道。因此,本研究參考史國安等[19]的牡丹花自然開放進程分級標準,以‘鳳丹’牡丹為試材,選擇切花保鮮處理最適宜時期——破綻期牡丹花苞作為研究對象,研究不同濃度ASA、H2O2溶液對瓶插切花衰老和不同瓶插時期花瓣相關(guān)生理指標的影響,以進一步探究牡丹切花衰老過程中抗氧化系統(tǒng)的運作機制、衰老生理及細胞學特性,以期為牡丹切花的保存及開發(fā)利用提供理論和技術(shù)依據(jù)。
1 材料與方法
1.1 試驗材料與地點
供試牡丹品種‘鳳丹’(Paeonia ostii)取自河南科技學院牡丹資源圃。試驗于2020年4月在河南省園藝植物資源利用與種質(zhì)創(chuàng)新工程研究中心進行。
1.2 材料培養(yǎng)與處理
于基地剪取破綻期生長態(tài)勢良好、大小一致的‘鳳丹’切花,共分為7組(T1~T7),每組6枝,分別置于盛有蒸餾水的切花瓶中培養(yǎng),其中每組預留一枝用作外表形態(tài)觀察和花徑測量,不用作生理指標測定。處理組分別滴加提前配置好的H2O2與ASA溶液,使瓶插液濃度分別為2.5 mg/L ASA(T1)、5 mg/L ASA(T2)、10 mg/L ASA(T3)和0.003 mg/L H2O2(T4)、0.03 mg/L H2O2(T5)、0.3 mg/L H2O2(T6),共6個處理組,對照組(T7)瓶插液為蒸餾水。將切花置于(22±1)℃室溫下培養(yǎng)觀察,全程保持自然光照,避免陽光直射。瓶插當日起,每日對切花的花徑和生理指標進行測定:于每日上午8時用游標卡尺測量每瓶預留切花兩個垂直方向的直徑大小,取其平均值作為該支切花的花徑;分別取切花內(nèi)、中、外輪花瓣的中間部位共0.5 g,切碎混勻,測定生理生化指標,每處理重復3次。
1.3 生理生化指標測定
采用考馬斯亮藍G-250法測定可溶性蛋白質(zhì)含量(參考Bradford[20]的方法并加以修改)。氮藍四唑(NBT)法測定SOD活性,愈創(chuàng)木酚法測定POD活性,紫外吸收法測定CAT活性[21]。根據(jù)硫代巴比妥酸(TBA)的三氯乙酸溶液與MDA的顯色反應測定MDA含量[22]。
1.4 DAPI染色觀察細胞核形態(tài)學特征
瓶插期間于每日上午8時分別取每個處理切花的中間層花瓣觀察細胞核變化,即從距離花瓣基部三分之一處切取長2~3 mm、寬1~2 mm的小塊,置于70%乙醇中固定2 h,之后用PBS緩沖液反復沖洗3次。將沖洗干凈的切片材料置于載玻片上,吸干表面水分,滴加10μg/mL DAPI染液20μL,于黑暗處染色15 min,然后置于尼康OLYMBUS熒光倒置顯微鏡下觀察拍照(熒光激發(fā)波長為UV330-380 nm)。判斷細胞核完整性的標準:細胞核完整時位于細胞正中央、形態(tài)完整、邊緣清晰圓滑,為標準的圓形,排列整齊。
1.5 數(shù)據(jù)整理與分析
采用Microsoft Execl制圖,SPSS軟件分析數(shù)據(jù),Photoshop軟件進行圖片整合。
2 結(jié)果與分析
2.1 不同濃度ASA和H2O2對牡丹切花花徑的影響
由圖1(a)可以看出,外源不同濃度ASA處理牡丹切花直徑變化顯著,第4天時‘鳳丹’切花花徑達到最大值,為完全盛開狀態(tài);第5天時濃度2.5、10 mg/L ASA處理切花的花徑仍然保持在最大值,其它處理切花花徑都存在一定程度縮減。這說明適宜濃度的ASA具有維持切花保鮮的作用。圖1(b)表明,外源H2O2處理下,切花花徑第2天時綻放程度明顯低于對照,并且在接下來幾天里始終低于對照。

圖1 不同濃度ASA和H2O2處理‘鳳丹’切花花徑的變化
2.2 不同濃度ASA和H2 O2對牡丹切花SOD活性的影響
由圖2(a)可知,第2天時各ASA處理花瓣內(nèi)SOD活性均比對照低,第3天時10 mg/L ASA處理(T3)切花SOD活性顯著升高且高于對照,到第5天處理組SOD活性整體高于對照。由圖2(b)可知,第2天時H2O2處理組切花SOD活性低于對照,之后開始逐漸升高,第4天達到峰值,之后開始下降,而0.3 mg/L H2O2處理(T6)SOD活性峰值提前1天,第5天T4處理切花SOD活性明顯低于對照,達到最低。

圖2 不同濃度ASA和H2O2瓶插處理‘鳳丹’切花SOD活性變化
2.3 不同濃度ASA和H2 O2對牡丹切花POD活性的影響
由圖3(a)可以看出,瓶插前4天各處理切花POD活性變化并不明顯,維持在較低狀態(tài),第5天顯著升高,以2.5、5 mg/L ASA處理組(T1、T2)最為明顯。圖3(b)顯示,0.3 mg/L H2O2處理(T6)切花POD活性第2天時顯著升高并達峰值,而后逐漸降低,而對照在第5天時升到最高,高于處理組。
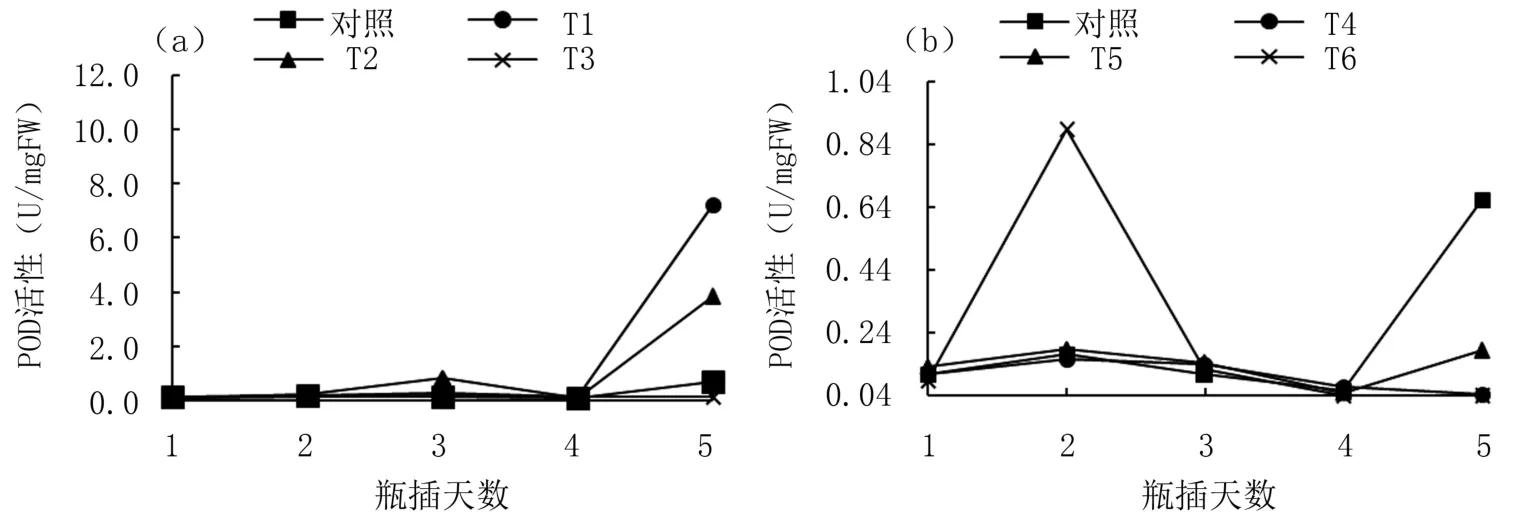
圖3 不同濃度ASA和H2 O2瓶插處理‘鳳丹’切花POD活性變化
2.4 不同濃度ASA和H2 O2對牡丹切花CAT活性的影響
圖4表明ASA和H2O2處理均對切花CAT活性變化有顯著影響。由圖4(a)可知,處理組切花CAT活性變化均較對照平穩(wěn),其中5 mg/L ASA處理(T2)切花CAT活性第2天升至最高,之后開始下降,第4天降至最低但仍高于對照。由圖4(b)可知,對照切花CAT活性先升高再降低,0.003 mg/L H2O2處理(T4)CAT活性峰值延后,其余濃度H2O2處理下CAT活性變化趨勢較穩(wěn)定,基本呈現(xiàn)先升高再降低趨勢,且在第5天均低于對照。
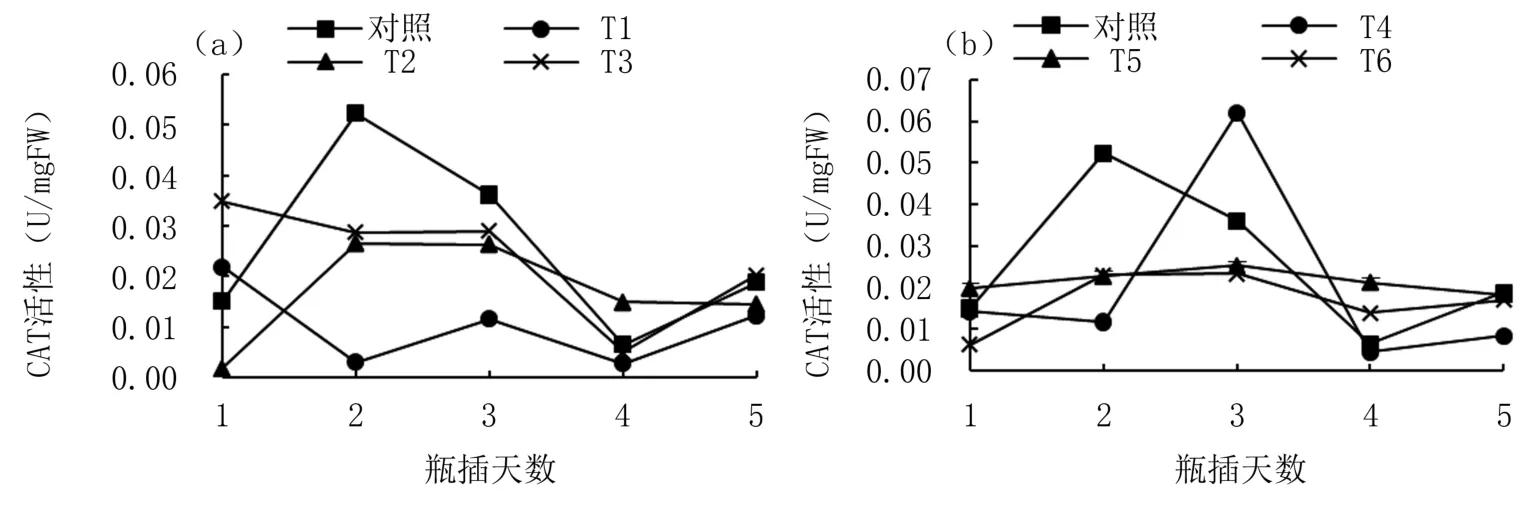
圖4 不同濃度ASA和H2O2瓶插處理‘鳳丹’切花CAT活性變化
2.5 不同濃度ASA和H2 O2對牡丹切花MDA含量的影響
由圖5(a)可知,ASA處理切花花瓣內(nèi)MDA含量第2天時明顯降低,之后開始緩慢上升,第4天花朵完全盛開時MDA含量均低于對照。這說明ASA能有效緩解‘鳳丹’花瓣細胞的膜質(zhì)過氧化程度,且T3處理MDA含量最低,即10 mg/L ASA對切花保鮮效果最好。圖5(b)顯示,第4天時切花花瓣細胞膜受到嚴重傷害,0.003、0.03 mg/L H2O2處理(T4、T5)下MDA含量的升高得到抑制,可有效緩解這種傷害;但T6處理MDA含量高于對照,這可能是由于過高濃度的H2O2對花瓣細胞膜造成損傷所致。

圖5 不同濃度ASA和H2O2瓶插處理‘鳳丹’切花MDA含量變化
2.6 不同濃度ASA和H2O2對牡丹切花細胞核變化的影響
細胞核內(nèi)部的DNA和染色體與DAPI結(jié)合后會在紫外光下發(fā)出強烈的藍色熒光,正常的植物細胞核形態(tài)表現(xiàn)為規(guī)整的圓形或者橢圓形(圖6-T7-1)。逆境脅迫下,為了滿足生理代謝所需的營養(yǎng)物質(zhì),細胞核會放大甚至變形來加強細胞吸收物質(zhì)的能力[23]。因此細胞核的形態(tài)變化可以作為檢驗植物細胞衰老的指標之一。
由圖6可知,瓶插第2天,對照切花有少數(shù)細胞核出現(xiàn)變形,細胞核向兩極拉伸,逐漸變?yōu)榧忓N狀或新月狀(圖6-T7-2),這是由核內(nèi)染色質(zhì)凝縮向核邊緣聚集所致。第2天、第3天,變形的細胞核數(shù)目逐漸增多。從第4天開始,顯微鏡視野下細胞核發(fā)出的熒光強度開始減弱、細胞核變小、大面積的細胞核發(fā)生固縮現(xiàn)象而緊貼在細胞邊緣,推測這是由染色質(zhì)凝縮聚集、液泡膜破裂擠壓細胞核所致。第5天,顯微鏡下的熒光信號進一步減弱,細胞核凝縮至最小,染色質(zhì)分散在細胞核邊緣發(fā)出微弱熒光。
T1~T3處理(圖6)為‘鳳丹’切花瓶插于不同濃度ASA中的細胞核變化情況:瓶插初期,與對照相比其細胞核的變形數(shù)目差異并不明顯,僅有部分細胞核發(fā)生拉長彌散現(xiàn)象;瓶插后期,2.5、5 mg/L ASA處理下細胞核維持高正常率且持續(xù)發(fā)出熒光的時間較對照久,直到第5天才出現(xiàn)部分細胞核濃縮現(xiàn)象,且5 mg/L ASA處理細胞核第5天依然保持高亮狀態(tài),但10 mg/L ASA處理細胞核第5天已經(jīng)完全彌散變形。
T4~T6處理(圖6)顯示:0.3、0.03 mg/L H2O2處理能較好維持‘鳳丹’切花細胞和熒光信號,瓶插第5天時大部分細胞核仍然保持規(guī)則明亮,以0.03 mg/L H2O2處理的效果最好。

圖6 不同瓶插處理‘鳳丹’切花花瓣細胞核(×10)變化情況
3 討論
花朵在脫離植物主體之后,光合作用和呼吸作用會繼續(xù)進行,若無法從瓶插液中吸收水分、礦質(zhì)營養(yǎng)、激素等植物生長必需的營養(yǎng)物質(zhì),切花花瓣和花梗就會發(fā)生一系列生理生化響應,最終導致細胞衰老和死亡。乙烯和超氧陰離子含量的迅速增加,會加速花瓣的衰老。因此,調(diào)節(jié)植物生理平衡是延長切花花期的重要因素。牡丹切花衰敗期的典型特征是花瓣枯萎、鮮重下降、花徑變小。本研究結(jié)果表明,ASA處理對切花的盛花期時長有影響,2.5 mg/L ASA促進切花綻放,使其花徑迅速達到最大,并在之后保持一段時間。這表明適宜濃度的ASA刺激切花的營養(yǎng)生長。H2O2處理下切花花徑增加緩慢,盛花期花朵直徑均低于對照,說明H2O2抑制切花的正常生理機制,減緩切花的代謝過程,從而達到長時間保鮮的效果。
正常植物細胞中活性氧的濃度并不高,但當受到逆境脅迫時,植物會加快細胞內(nèi)的生理活動及代謝過程,產(chǎn)生過量的ROS,這些過量的ROS又對植物本身造成氧化傷害,導致植物逐漸衰老[24,25]。因此,植物的衰老也是一種氧化過程[26]。為了控制活性氧的含量,植物進化出一套與之對抗的抗氧化清除系統(tǒng),它通過調(diào)節(jié)相關(guān)保護酶的活性和非酶類抗氧化劑含量,來維持生理機制的正常運行。本研究通過對‘鳳丹’切花施加外源ASA、H2O2,測定其在瓶插期間的花徑大小、MDA含量,觀察其細胞核形態(tài)變化,以此來判斷‘鳳丹’衰老程度,再通過檢測衰老過程中花瓣保護酶活性的變化,來推測‘鳳丹’切花衰老過程中抗氧化清除機制的運作規(guī)律。超氧化物歧化酶(SOD)能夠催化細胞內(nèi)歧化生成O2和H2O2,而過量的會造成生物體內(nèi)的組織損傷,因此SOD是植物細胞重要保護酶,在機體氧化與抗氧化平衡中起到至關(guān)重要的作用。本試驗結(jié)果表明,施加一定濃度的外源ASA和H2O2后,SOD活性在瓶插第2天顯著降低,說明植物細胞內(nèi)超氧陰離子沒有過量增長而刺激SOD產(chǎn)生反應,這可能是由于外源ASA提高ASA和GSH(谷胱甘肽)組成的ASA-GSH循環(huán)系統(tǒng)的代謝活動,清除大部分瓶插初期植物細胞產(chǎn)生的活性氧,因此沒有觸發(fā)SOD活性升高;外源H2O2的刺激作為一種逆境信號分子,激活植物細胞內(nèi)其它抗氧化酶的活性,例如過氧化物酶和過氧化氫酶,短暫維持細胞正常生理代謝,延遲SOD活性的增強。這與楊慶賀[13]、蔣景龍[18]等的研究結(jié)果一致。過高濃度的ASA和H2O2促使SOD活性高于對照或者峰值提前,推測是由于高濃度的外源處理對切花產(chǎn)生脅迫作用,ROS水平急劇增加,繼而細胞產(chǎn)生過氧化應激反應,致使SOD活性迅速升高。
過氧化物酶(POD)是氧化還原酶的一種,在植物體中大量存在,且活性在植物生長發(fā)育過程中不斷發(fā)生變化。有研究表明,POD在生物體老化組織中活性較高,因此這種酶可以作為植物組織老化的標志性酶。本試驗發(fā)現(xiàn),外源ASA處理前期,POD活性變化不顯著;切花瓶插后期2.5、5 mg/L ASA處理下,POD活性升高,且都較對照高。這說明瓶插后期切花花瓣細胞開始受到活性氧的脅迫,為了抵御這種脅迫,植物自身激發(fā)POD活性上升,來清除過多的自由基,而適宜濃度的ASA可以維持這種平衡。外源H2O2處理下,細胞內(nèi)POD活性整體出現(xiàn)類似的變化趨勢,但在0.3 mg/L H2O2處理下,POD活性在第2天出現(xiàn)急劇上升,之后下降并持續(xù)保持在較低水平。這是因為過高濃度的H2O2從一開始就對切花產(chǎn)生逆境脅迫,短暫觸發(fā)抗氧化酶活性,POD活性急劇升高。在植物抗氧化系統(tǒng)感知高濃度的H2O2影響超出植物自身的修復能力之后,POD隨即逐漸失去活性。
過氧化氫酶(CAT)能夠清除植物細胞內(nèi)的過氧化氫,把其分解成H2O和O2,使植物免受H2O2的傷害,通常與SOD協(xié)同作用。本試驗結(jié)果表明,瓶插期間‘鳳丹’切花CAT活性出現(xiàn)明顯變化,瓶插前期出現(xiàn)較大增幅,而適宜濃度的外源ASA、H2O2處理下,相較于對照,這種增長趨勢被減緩或者推遲。這可能是外源ASA和H2O2協(xié)同影響‘鳳丹’切花細胞內(nèi)的ROS含量,從而導致花瓣細胞內(nèi)CAT活性趨于穩(wěn)定。這與蘇軍等[27]關(guān)于小蒼蘭的研究結(jié)果一致。但如何緩解氧自由基引起的傷害以及延長牡丹切花瓶插時間的最合適濃度,還有待進一步研究。
正常狀態(tài)下完整細胞核數(shù)量與花朵的生長狀態(tài)呈正相關(guān),細胞核完整性越高,花瓣細胞排列越整齊一致。正常細胞核邊緣清晰,呈現(xiàn)標準的圓形,當受到外界環(huán)境脅迫時,細胞核產(chǎn)生應激反應變形延伸,導致液泡膜破裂、細胞液將細胞核擠壓至邊緣,此時細胞核呈現(xiàn)向兩極拉伸狀、緊貼細胞壁[32]。本試驗采用形態(tài)學觀察法,利用DAPI與細胞核中DNA特異性結(jié)合發(fā)出的藍色熒光來觀測細胞核形態(tài),結(jié)果顯示:對照‘鳳丹’切花細胞核形態(tài)呈規(guī)律性變化,瓶插后期逐漸固縮變形直至核膜破裂、核質(zhì)流失;而適宜濃度的ASA、H2O2處理下‘鳳丹’切花瓶插期間細胞核發(fā)生變形的時間被延遲,細胞核更加持久地維持規(guī)則明亮狀態(tài),其中濃度5 mg/L ASA和0.03 mg/L H2O2效果最為顯著,而ASA、H2O2濃度過大或過小都有可能使花瓣受到脅迫導致細胞核濃縮或者出現(xiàn)彌散情況。
4 結(jié)論
本試驗關(guān)于牡丹切花瓶插壽命的研究發(fā)現(xiàn):輔以外源化學物質(zhì)調(diào)控,切花花瓣在衰老過程中其生理指標與細胞核形態(tài)指標雖然都存在變化,但是發(fā)生的重要程度、發(fā)生時期與時間順序不完全吻合。外源ASA處理下CAT活性峰值出現(xiàn)最早,其次為SOD、POD。在其它作物研究中,保護酶活性的變化規(guī)律分別有不同的報道[33,34],這可能是因為不同植物其應對逆境脅迫的響應機制和發(fā)揮作用的途徑不同,也可能是內(nèi)部調(diào)控基因的差異所造成。通過對切花生理學及細胞學的研究觀察,結(jié)合‘鳳丹’花徑的變化規(guī)律,得出初步結(jié)論:5 mg/L ASA和0.03 mg/L H2O2處理能有效維持切花細胞抗氧化酶清除系統(tǒng)的平衡,減輕活性氧對細胞膜的破壞作用;外源ASA、H2O2對‘鳳丹’切花具有保鮮作用的細胞學機制在于緩解花瓣細胞核彌散程度,維持細胞核形態(tài)。本研究結(jié)果可為進一步揭示外源ASA、H2O2對牡丹切花保鮮時間的延長提供試驗依據(jù),為未來研究牡丹切花衰老機制提供理論參考。
另外,關(guān)于外源ASA、H2O2對牡丹切花生理生化活性及細胞學特性的具體影響機制還待進一步研究;牡丹切花細胞程序性死亡相關(guān)各類事件的重要程度、發(fā)生的順序和互相作用機制也許能為牡丹切花保鮮研究提供新的思路;細胞核變形彌散情況或?qū)⒊蔀橐环N新的判斷植物衰老的可行性指標。
作者貢獻:邱永杰是本研究的試驗設(shè)計者和試驗執(zhí)行人,完成論文初稿寫作;何松林是項目的構(gòu)思者及負責人;胡緩參與協(xié)助了實驗操作過程;張翔宇、于嘉倫和史來琨完成數(shù)據(jù)整理與分析;賈文慶指導試驗設(shè)計、數(shù)據(jù)分析、論文寫作與修改。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
