減磷條件下含褪黑素的尿素緩釋功能肥對番茄生長、產量、品質和磷素利用效率的影響
2023-03-07 02:31:12劉明慧田虹雨劉之廣鞏彪
中國農業科學 2023年3期
劉明慧,田虹雨,劉之廣,鞏彪
減磷條件下含褪黑素的尿素緩釋功能肥對番茄生長、產量、品質和磷素利用效率的影響

1山東農業大學園藝科學與工程學院/作物生物學國家重點實驗室,山東泰安 271018;2山東農業大學資源與環境學院/土肥高效利用國家工程研究中心,山東泰安 271018
【目的】褪黑素對植物具有多種有益功效,但其化學性質活潑,限制了其在農業生產中的應用。本研究通過包膜技術制備出含褪黑素的尿素緩釋功能肥(以下簡稱功能肥),研究其對番茄生長、產量、品質和磷素利用效率的影響,為褪黑素高效施用和番茄節肥生產提供理論依據。【方法】首先以水泡法研究功能肥中褪黑素的釋放速率,然后以穴盤育苗方式研究功能肥對番茄幼苗生長的影響。采用盆栽方式,設置4個處理,分別是施磷(記作+P)和不施磷(記作-P)兩個磷處理水平,再分別添加尿素緩釋肥(對照,仍記作-P和+P)或功能肥(記作-P+M和+P+M),研究功能肥對植株生長和干物質分配、根系生長、磷素吸收和分配、磷肥利用率、肥料產量貢獻率、根系磷酸酶活性、果實產量和品質的影響。【結果】利用褪黑素制備功能肥時存在一定的損耗,包膜后褪黑素的實際含量為包膜時褪黑素總用量的35%。水泡至60 d時,功能肥中褪黑素殘余量為6.61%。番茄穴盤育苗中施用功能肥能顯著促進幼苗生長,壯苗指數較對照提升70.2%。盆栽試驗表明,-P處理下番茄根、莖、葉、果的生物量分別較+P處理下降低了19.64%、18.51%、28.99%和28.73%,根和莖的干物質分配比例增加了10.03%和11.63%,葉和果的干物質分配比例分別減少了2.74%和2.39%。應用功能肥能顯著提升兩種磷處理條件下所有組織的生物量積累,增加+P條件下根的干物質分配比例達12.14%,降低-P條件下莖和葉的干物質分配比例達6.00%和5.90%,增加9.06%的果干物質分配比例。-P處理降低根系的總長、總表面積和總體積,增加根尖數量,施用功能肥能提高兩種磷水平下根系的總長、總表面積、總體積和根尖數量。-P處理顯著降低了番茄植株根、莖、葉、果的磷含量,增加根、莖和葉的磷分配比例,減少果的磷分配比例。應用功能肥能顯著提升兩種P處理條件下所有組織的磷含量,+P條件下根的磷分配比例,-P條件下莖和葉的磷分配比例,增加果的磷分配比例。應用功能肥能顯著提高磷酸酶的活性、全株磷吸收量、肥料利用率與肥料產量貢獻率。-P處理使番茄產量降低了17.57%,但-P處理下應用功能肥能使產量提高21.32%,而對+P處理下的產量影響不顯著。此外,應用功能肥能全面提高-P或部分提高+P條件下番茄果實的品質。【結論】將褪黑素包膜于緩釋肥能顯著降低褪黑素的用量,提高番茄幼苗及植株全生育期的生長質量,增加磷肥利用效率,提高產量和果實品質。
番茄;含褪黑素的尿素緩釋功能肥;磷肥處理;生長;產量品質;磷利用率
0 引言
【研究意義】磷是植物生長發育所必需的大量營養元素,雖然土壤中的全磷含量可達1—1.5 g·kg-1,但可被植物直接吸收利用的可溶性磷含量卻不足0.05 mg·kg-1,是作物生長和產量形成的重要營養限制因子[1]。施用磷肥能顯著改善土壤磷營養狀況,但作物當季對磷肥的吸收率通常較低,約為10%—25%,其余絕大多數磷素被土壤固定或滲入地下水系統,造成土壤面源污染[2]。褪黑素不僅是一種動物激素,在促進植物生長、提高礦質營養利用效率和生物/非生物脅迫抗性等方面也具有多種功效。但褪黑素化學性質活潑,暴露于空氣中易氧化,見光易分解,限制了其在農業生產中的應用[3]。研究褪黑素高效施用技術,開發褪黑素易存、長效施用產品,明確其在促進作物生長和提高磷利用效率中的協同功效具有重要的理論和應用價值。【前人研究進展】褪黑素與吲哚乙酸(IAA)在生物體內都以色胺為合成前體,其化學性質和生理活性有很多相似之處[4]。生長素受體,信號響應因子和極性運輸載體是磷饑餓條件下植物調控根系生長的重要功能基因[5]。雖然褪黑素調控植物根系形態建成不依賴于IAA信號[6],但是褪黑素仍能與IAA協同,通過微調IAA合成和運輸促進側根和不定根發育[7-8]。外源添加褪黑素在調控植物根系生長的表現特征上與缺磷處理下根系的生育特征具有相似性,說明褪黑素可能通過調控根系構型來促進磷吸收。筆者研究發現,在番茄中超表達褪黑素合成關鍵基因咖啡酸--甲基轉移酶1()能顯著提高水培條件下番茄幼苗的磷吸收利用效率;經代謝組學分析,發現低磷脅迫下能促進根系多種有機酸積累和磷脂代謝,促進磷素吸收和再利用[9]。外源施用100 μmol?L-1褪黑素還能顯著促進高氮條件下黃瓜幼苗的根系生長,提高硝酸還原酶(NR)和谷氨酸合酶(GOGAT)活性,促進氮同化進程,平衡磷吸收[10]。除了促進礦質營養吸收利用外,外源添加褪黑素還能提高干旱脅迫下大豆產量[11],延緩長期干旱脅迫下蘋果葉片的衰老[12],增加番茄幼苗高溫、低溫、干旱等脅迫的綜合抗性[13],促進番茄果實成熟等[14]。說明褪黑素是一種具有促進植株生長、增強抗性、提高產量的多效型植物生長調節劑,具有良好的農業應用前景。褪黑素在不同植物中的生理濃度約為0.1—10 μmol·L-1,但多數外源藥物學試驗卻將褪黑素的最優濃度界定在100—200 μmol·L-1[3,15],說明外源添加的褪黑素被植物直接吸收、利用的效率極低,這可能是由植物吸收能力及褪黑素在環境中的釋放和分解等多重因素造成。【本研究切入點】雖然褪黑素對植物的有益效果已廣泛報道,但有關外源褪黑素的高效施用技術,及其在全生育期對植物磷吸收利用的影響尚屬空白。【擬解決的關鍵問題】本研究借鑒緩釋肥理念,將褪黑素與緩釋尿素共包膜制成含褪黑素的尿素緩釋功能肥(以下簡稱功能肥)。研究功能肥的褪黑素釋放規律,及其對番茄苗期和全生育期植株生長、產量、品質和磷素利用效率的影響。以期為褪黑素在促進植物生長、產量和磷肥利用方面提供新理論、新技術和新產品。
1 材料與方法
試驗于2020—2021年在山東農業大學日光溫室和園藝實驗站(山東泰安)進行。
1.1 試驗材料
供試番茄(L.)品種為‘金棚M6’。采用蠟封法制備功能肥:稱取聚烯烴蠟50 g,油浴加熱使蠟粉熔化,將10 g褪黑素置于熔化后的蠟內,使蠟盡可能地包裹褪黑素,待用。每次將2 kg尿素(3—5 mm,46.4% N)加入轉鼓,預熱至75℃。加熱后,將12 g含褪黑素的聚烯烴蠟加入轉鼓內作為底涂層,反應約5—10 min,使含褪黑素的聚烯烴蠟均勻地涂覆在尿素表面。然后將混合環氧樹脂包膜液(17 g環氧樹脂E-44與3 g三乙烯四胺混合)加入轉鼓中,反應約8—10 min,完成第一次包膜,該過程重復4次,包膜厚度為4%,膜顏色使用紅、黃、藍3種顏色按體積比調制成深黑色,共包裹10 kg尿素,完成褪黑素含量理論值為0.1%的功能肥制作。以同等工藝制造的不添加褪黑素的尿素緩釋肥(以下簡稱對照肥)為對照。
1.2 試驗方法
1.2.1 有效褪黑素含量及其水泡釋放速率測試 參照楊相東等[16]的水泡法,稱取含褪黑素的尿素緩釋功能肥10 g,置于尼龍網袋,將肥料網袋浸泡在含200 mL蒸餾水的密閉玻璃瓶中,于25℃的黑暗環境下靜置,浸提。于處理后10、20、30、40、50和60 d取殘余功能肥,人工破損包膜,充分浸提殘存褪黑素后,采用高效液相色譜法測定殘存褪黑素含量。測定未經浸提的功能肥(0 d)中的褪黑素含量,即為功能肥中褪黑素含量的實際值。將此含量計為100%,計算水泡不同時間下功能肥中褪黑素的殘余量(%)=水泡功能肥中褪黑素含量/未水泡功能肥中褪黑素含量×100。每個處理重復3次。
1.2.2 功能肥的番茄幼苗生長試驗 2020年秋,在山東農業大學日光溫室采用96孔穴盤和商業育苗基質進行番茄育苗。褪黑素生理濃度參照筆者前期研究結果[17],按0.5 μmol?L-1(116 μg·L-1)計,每穴(40 cm3)于播種前施入13.26 mg對照肥(記為CK)或13.26 mg功能肥(記為M)。每個處理9盤,每3盤作為1次重復,共計3次重復。之后將催芽至露白的番茄種子播于CK和M處理下的穴盤中。育苗期保持22—30℃/15—20℃的晝夜溫周期,每天澆灌1/2山崎番茄配方營養液,25 d后統計番茄幼苗生長相關指標。
1.2.3 功能肥的盆栽番茄試驗 2021年春,在山東農業大學園藝實驗站采用盆栽方式,供試陶盆裝干土15 kg(約10 L),土壤基本理化性質見表1。參照番茄最優磷營養需求量(0.53 g?kg-1P2O5)[18],以該施入量作為番茄磷肥施用標準,設置施磷(記作+P)和不施磷(記作-P)兩個磷處理水平。褪黑素生理濃度仍按0.5 μmol?L-1計[17],在兩種磷處理水平下分別添加3.32 g對照肥(記作-P和+P)或3.32 g功能肥(記作-P+M和+P+M),共計4個處理,每個處理30盆,每10盆作為1次重復,共計3次重復。于處理好的土壤中每盆定植一棵30日苗齡的番茄幼苗,番茄留4穗果打頂。除磷素外,其他礦質營養正常供應,水肥及病蟲害管理參考前人文獻描述[19]。

表1 土壤基本理化性質
1.3 數據測定
1.3.1 不同水泡時間功能肥的褪黑素殘余量測定 將未經水泡或水泡不同時間的功能肥磨碎,溶于50 mL蒸餾水中充分浸提,采用高效液相色譜法測定溶液褪黑素含量,色譜條件參照文獻[17]。
1.3.2 番茄幼苗的生長量分析 于播種后25 d測量不同處理的番茄株高、莖粗、莖葉生物量(干重)和根系生物量(干重),計算根冠比=根系生物量/莖葉生物量,壯苗指數=(莖粗/株高+根冠比)×全株生物量[20]。
1.3.3 番茄植株的干物質分配 番茄于定植后生長至90 d時,測定番茄植株根、莖、葉、果的干重,計算總生物量(g)=根干重+莖干重+葉干重+果干重,各器官的干物質分配(%)=各組織的干重/全株干重×100。
1.3.4 番茄根系的生長量分析 番茄于定植后生長至90 d時,取番茄植株完整根系,清水洗凈,利用WinRHIZO根系分析系統掃描根系樣品,獲取數字化圖像,分析根系總長、根系總表面積、根系總體積和根尖數量[21]。
1.3.5 番茄植株的磷素吸收和分配分析 將1.3.3中烘干的根、莖、葉、果用磨樣器研磨成粉,各組織取0.1 g粉末,采用鉬銻抗比色法測定磷含量[22]。計算植株磷吸收量(mmol/plant)=根P含量×根干重+莖P含量×莖干重+葉P含量×葉干重+果P含量×果干重,各器官的磷分配比例(%)=該器官的磷含量×該器官的干重/植株磷吸收量×100[22]。
1.3.6 番茄單株產量、磷肥利用率和肥料產量貢獻率分析 番茄于各穗果實成熟后收獲,稱重,記錄單株產量。計算磷肥利用率(%)=(施磷處理植株磷吸收量-缺磷處理植株磷吸收量)/磷肥中P2O5的施用量×100,肥料產量貢獻率(%)=(施肥處理番茄單株產量-缺素處理番茄單株產量)/施肥處理番茄單株產量×100[23]。
1.3.7 番茄根系磷酸酶活性和果實營養品質分析 于定植后90 d取番茄根系組織1 g,采用對硝基苯酚比色法測定根系磷酸酶活性[24]。摘取第3穗完全成熟的果實用于品質分析,采用糖度計測定可溶性固形物含量,蒽酮比色法測定可溶性糖含量,NaOH滴定法測定可滴定酸含量,糖酸比=可溶性糖含量/可滴定酸含量,鉬藍比色法測定維生素C含量[23]。
1.4 數據分析
數據用SAS Verion 8.1進行方差分析,采用LSD法進行多重比較(<0.05),不同字母表示處理間差異達5%顯著水平。
2 結果
2.1 功能肥的有效褪黑素含量及其浸泡釋放速率分析
對功能肥中褪黑素的含量測定可知,當包膜褪黑素的理論值為1 mg?g-1時,實際值為0.35 mg?g-1,有效包膜的褪黑素含量占35%。浸泡試驗表明水泡至30 d時,功能肥中褪黑素殘余量為53.15%,接近半釋放劑量,至60 d時,功能肥中褪黑素殘余量僅占6.61%(圖1)。說明功能肥中褪黑素的有效釋放周期約60 d。
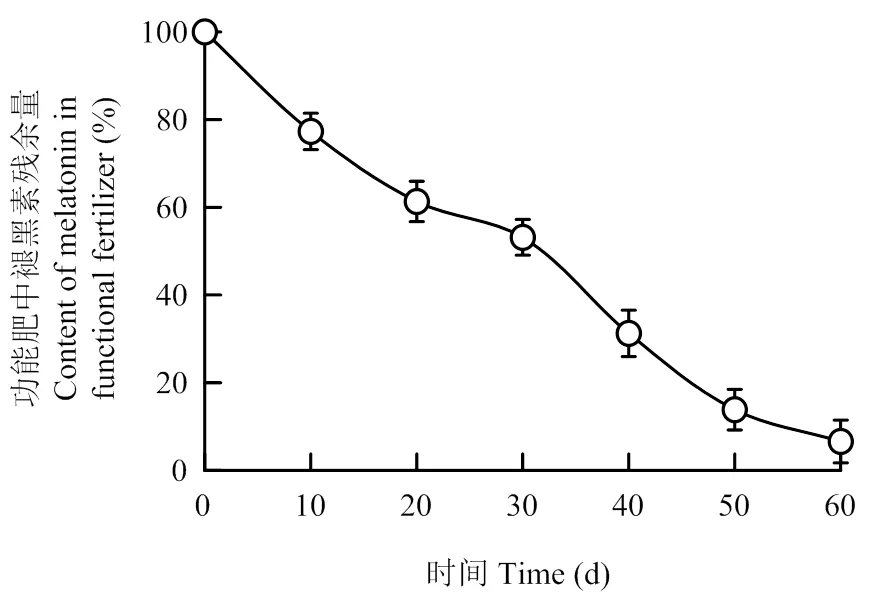
圖1 功能肥中的褪黑素釋放速率
2.2 功能肥對番茄幼苗生長的影響
M處理下番茄幼苗的株高、莖粗、莖葉生物量、根系生物量、根冠比以及壯苗指數分別較CK提高了40.69%、16.92%、38.16%、85.71%、32.14%和68.18%(表2)。即施用功能肥能促進番茄幼苗生長,利于壯苗形成。
2.3 功能肥對番茄收獲期植株生物量和各器官干物質分配的影響
表3中各組織干重數據表明,與+P處理組相比,-P處理組顯著降低了番茄植株根、莖、葉、果及總生物量的積累;而應用含褪黑素的尿素緩釋功能肥卻能顯著提升兩種磷處理條件下所有組織的生物量積累。
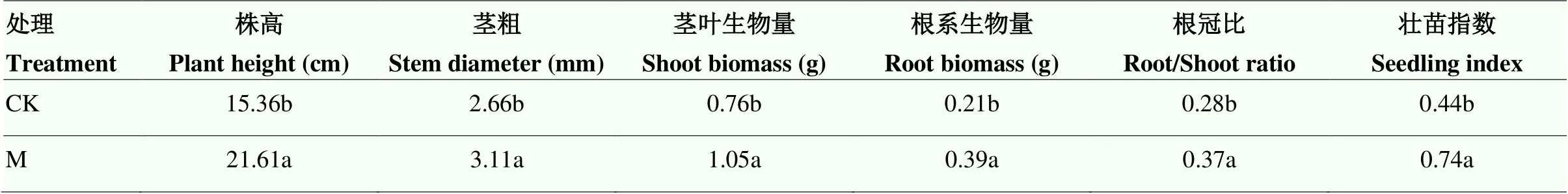
表2 功能肥對番茄幼苗生長的影響
不同小寫字母表示差異顯著(<0.05)。下同
Different lowercase letters indicate significant difference (<0.05). The same as below

表3 功能肥對番茄全生育期植株生長和干物質分配的影響
干重:Dry weight (DW, g/plant);分配比例:Allocation rate (AR, %);總生物量:Total biomass (TB, g/plant)
與+P處理相比,-P處理下的番茄植株根和莖的干物質分配量顯著增加,葉和果的干物質分配量顯著減少。+P條件下施用含褪黑素的尿素緩釋功能肥能顯著增加番茄根的干物質分配量,對莖、葉、果的干物質分配量影響不顯著。而-P條件下施用含褪黑素的尿素緩釋功能肥能顯著降低番茄莖和葉的干物質分配量,增加果的干物質分配量,對根的干物質分配量影響不顯著(表3)。
2.4 功能肥對番茄收獲期根系發育的影響
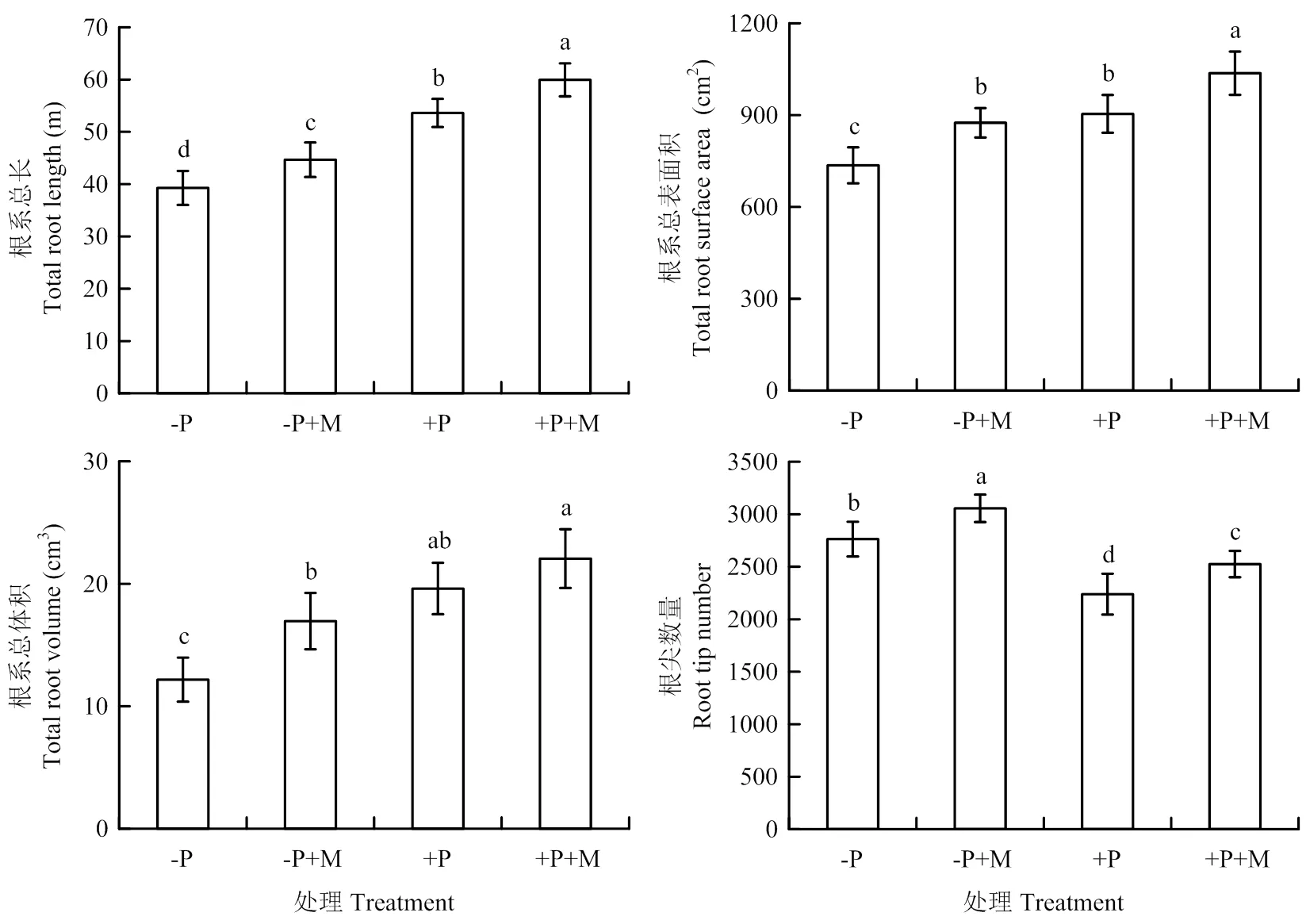
圖2顯示,-P處理下番茄根系的總長、總表面積和總體積顯著低于+P處理,但-P處理下的根尖數量卻顯著高于+P處理。在兩種磷水平下施用含褪黑素的尿素緩釋功能肥均能提高番茄根系的總長、總表面積、總體積和根尖數量。其中,在-P+M處理下的根系總表面積和總體積能恢復至+P處理水平。說明含褪黑素的尿素緩釋功能肥兼具與低磷促進根尖形成和高磷促進根系延長相似的效果。
2.5 功能肥對番茄收獲期個器官磷吸收利用的影響
與+P處理相比,-P處理顯著降低了番茄植株根、莖、葉、果的磷含量;而應用含褪黑素的尿素緩釋功能肥均能提升兩種磷處理條件下各組織的磷含量。-P+M和+P兩個處理組中番茄各組織的磷含量差異不顯著(表4)。
不同小寫字母表示差異顯著(P<0.05)。下同 Different lowercase letters indicate significant difference (P<0.05). The same as below
磷分配數據表明,與+P處理相比,-P處理下的番茄植株根、莖和葉的磷分配量顯著增加,果的磷分配量顯著減少。+P條件下施用含褪黑素的尿素緩釋功能肥能顯著增加番茄根的磷分配量,對莖、葉、果的磷分配量影響不顯著。而-P條件下施用含褪黑素的尿素緩釋功能肥能顯著降低番茄莖和葉的磷分配量,增加果的磷分配量,對根的磷分配量影響不顯著。
與+P處理相比,-P處理顯著降低了番茄全株磷吸收量,應用含褪黑素的尿素緩釋功能肥雖能顯著提高不同磷水平下的磷吸收量,但是仍無法使-P+M處理的磷吸收量達到+P處理水平。應用含褪黑素的尿素緩釋功能肥能顯著提高肥料利用率與肥料產量貢獻率,這與磷吸收量數據相吻合。
以上數據暗示應用含褪黑素的尿素緩釋功能肥可能增強了番茄對土壤有機磷的利用,而根系磷酸酶活性的強弱是影響植物分解有機磷,獲取磷素的關鍵。-P水平下的根系磷酸酶活性顯著高于+P水平,應用含褪黑素的尿素緩釋功能肥能顯著提高兩種磷水平下磷酸酶的活性(圖3)。

表4 功能肥對番茄全生育期磷利用的影響
濃度:Concentration (C, mmol?g-1);分配比例:Allocation rate (AR, %);吸收量:Uptake (U, mmol/plant);肥料利用率:Fertilizer utilization (FU, %);肥料產量貢獻率:Contribution of fertilizer to yield (CFY, %)
2.6 功能肥對番茄產量品質的影響
-P處理較+P處理顯著降低了番茄的單果重和單株產量,但應用含褪黑素的尿素緩釋功能肥能顯著提高-P處理下的單果重和單株產量,而對+P處理下的單果重和單株產量影響不顯著(表5)。果實成熟期數據表明,-P和+M處理條件均能顯著促進番茄成熟(表5)。可溶性固形物、可溶性糖、可滴定酸、糖酸比和維生素C等品質指標顯示,-P降低番茄果實品質,+M能全面提高-P條件下番茄果實的品質,部分提高+P條件下番茄果實的品質。
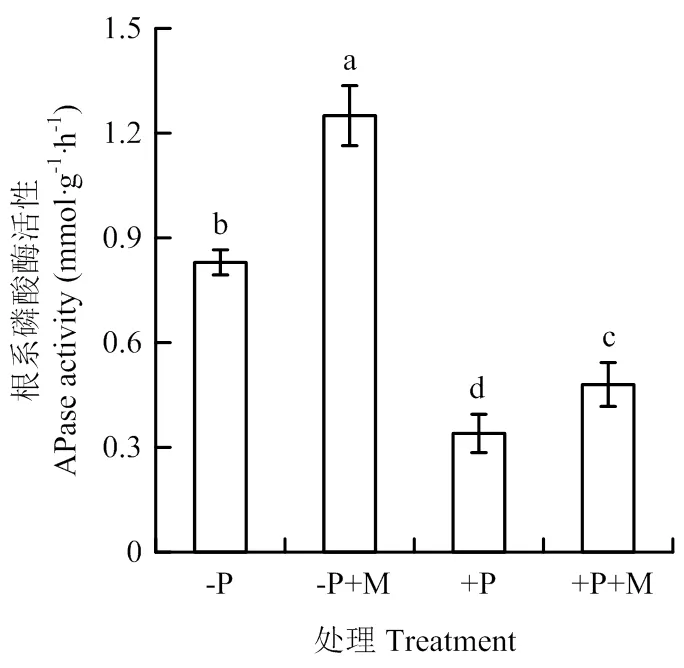
圖3 功能肥對番茄全生育期根系磷酸酶活性的影響
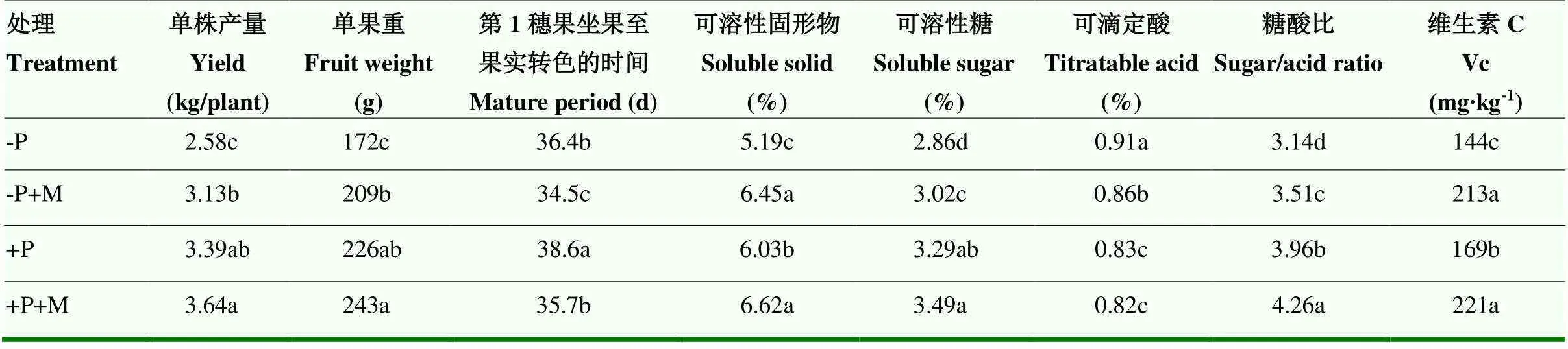
表5 功能肥對番茄全生育期番茄品質的影響
3 討論
3.1 功能肥對番茄磷吸收和轉運的影響
生理濃度的褪黑素有利于作物根系生長,抑制莖(下胚軸)的伸長,形成較高的根冠比[3,25]。而果菜類蔬菜壯苗的典型特征就是具有相對較粗的莖稈、較短的節間距和較大的根冠比。因此,本研究在添加少量褪黑素的條件下即顯著提高了番茄幼苗的壯苗指數,顯著低于以往報道的葉面噴施及澆灌褪黑素的施用濃度[13,26]。
缺磷抑制作物生長發育,降低植株干重,改變了同化物在地上部和根系的分配[5]。生長素是促進根系,尤其是側根和根毛發育的重要激素之一[27]。大田研究表明,適度減少磷肥施用量,能顯著提高番茄的根系生物量和側根數量[28]。這在很大程度上歸因于磷饑餓響應基因()對生長素信號基因表達的誘導效應[5]。褪黑素在促進植物根系發育的表現特征上與生長素極為相似[29],盡管有研究明確指出褪黑素和生長素信號在調控植物根系發育上是相互獨立的[6,30-31],但這兩個信號物質之間的相互誘導和協同增效的特性也被大量報道[7-8]。與+P處理相比,-P處理增加了番茄根系干重的分配比例。施加褪黑素后,根系生長進一步加強,說明本研究中含褪黑素的尿素緩釋功能肥的施用促進了干物質由地上部向根系的分配,增加了根系生物量和密度,有利于提高磷肥利用效率。雖然本研究中-P處理組不施磷肥,但土壤中仍含有大量的有機磷營養。低磷脅迫條件下,植物通過增強酸性磷酸酶的合成和分泌,活化土壤難溶性有機磷和無機磷,提高土壤中磷的有效性[32]。另外,本研究發現,施加含褪黑素的尿素緩釋功能肥會提高番茄根系磷酸酶活性,以此促進根系對磷的吸收和利用,這可能是褪黑素的增益效果在-P條件下高于+P條件下的重要原因之一。
3.2 功能肥對番茄果實產量和品質的影響
植物在干旱、營養虧缺等不利自然條件下會促進營養物質由莖葉器官向生殖器官轉移,以加速繁殖進程,保障世代更替[33]。-P處理較+P處理顯著提高了果實干物質分配比例,縮短了果實成熟時期。乙烯是重要的磷饑餓響應激素,能誘導植物體內合成大量磷酸酶和果膠酶,水解細胞壁固著的磷為可溶性磷,從而實現植物對磷素的二次分配和高效利用[34]。此外,乙烯也能促進果實成熟和品質提升。已有研究表明,外源褪黑素能直接激活番茄果實ACC合酶4(ACS4)促進乙烯合成,加速果實成熟和品質提升[14]。基于褪黑素和磷饑餓在調控乙烯信號中的研究進展,含褪黑素的尿素緩釋功能肥對番茄植株磷分配和果實品質的影響可能與乙烯激素信號調控有關。
本試驗條件下,含褪黑素的尿素緩釋功能肥中有效包膜的褪黑素含量僅占35%,因此,在后續工藝優化方面須考慮加入抗氧化劑等策略,以提高褪黑素的有效率。其次,針對含褪黑素的尿素緩釋功能肥的施用對象,可通過調控包膜厚度,分別制造成釋放周期小于30 d的短效褪黑素型育苗功能肥和釋放周期大于100 d的長效褪黑素型豐產功能肥。
4 結論
含0.1%褪黑素的尿素包膜緩釋肥可顯著提高番茄幼苗的壯苗指數,提高干物質分配比例、肥料利用率與肥料產量貢獻率、全株磷吸收量以及提高磷酸酶的活性。因此,將褪黑素包膜于緩釋肥是一種在農業生產中高效、可行的外源褪黑素應用方案,能顯著降低褪黑素的用量,提高番茄幼苗及植株全生育期的生長質量,增加磷肥利用效率,提高番茄產量和果實品質。
[1] 許秀美, 邱化蛟, 周先學, 于瑞忠, 冷壽慈. 植物對磷素的吸收、運轉和代謝. 山東農業大學學報(自然科學版), 2001, 32(3): 397-400. doi: 10.3969/j.issn.1000-2324.2001.03.031.
XU X M, QIU H J, ZHOU X X, YU R Z, LENG S C. The absorption, translocation and metabolism of phosphorus of plant. Journal of Shandong Agricultural University, 2001, 32(3): 397-400. doi: 10. 3969/j.issn.1000-2324.2001.03.031. (in Chinese)
[2] 胡寧, 袁紅, 藍家程, 袁道先, 傅瓦利, 文志林. 巖溶石漠化區不同植被恢復模式土壤無機磷形態特征及影響因素. 生態學報, 2014, 34(24): 7393-7402. doi: 10.5846/stxb201303190452.
HU N, YUAN H, LAN J C, YUAN D X, FU W L, WEN Z L. Factors influencing the distribution of inorganic phosphorus fractions in different vegetation restoration areas in Karst rocky desertification areas. Acta Ecologica Sinica, 2014, 34(24): 7393-7402. doi: 10.5846/ stxb201303190452. (in Chinese)
[3] 鞏彪, 史慶華. 園藝作物褪黑素的研究進展. 中國農業科學, 2017, 50(12): 2326-2337. doi: 10.3864/j.issn.0578-1752.2017.12.013.
GONG B, SHI Q H. Review of melatonin in horticultural crops. Scientia Agricultura Sinica, 2017, 50(12): 2326-2337. doi: 10.3864/j. issn.0578-1752.2017.12.013. (in Chinese)
[4] ARNAO M B, J HERNáNDEZ-RUIZ. Melatonin against environmental plant stressors: A review. Current Protein and Peptide Science, 2021, 22(5): 413-429.
[5] CRNMBEZ H, MOTTE H, BEECKMAN T. Tackling plant phosphate starvation by the roots.Developmental Cell, 2019, 48(5): 599-615.
[6] RAMóN, PELAGIO-FLORES, EDITHh,MUOZ-PARRA, RANDY, ORTIZ-CASTRO, JOEé, LóPEZ-BUCIO. Melatonin regulatesroot system architecture likely acting independently of auxin signaling. Journal of Pineal Research, 2012, 53(3): 279-288.
[7] WEN D, GONG B, SUN S S, LIU S Q, WANG X F,WEI M, YANG F J, LI Y, SHI Q H. Promoting roles of melatonin in adventitious root development ofL. by regulating auxin and nitric oxide signaling. Frontiers in Plant Science, 2016, 7: 718.
[8] REN S X, RUTTO L, KATUURAMU D, OONO Y. Melatonin acts synergistically with auxin to promote lateral root development through fine tuning auxin transport in. PLoS ONE, 2019, 14(8): e0221687.
[9] XU Y, SHI Q H, GONG B. Characterization of-mediated low phosphorus resistance mechanism by metabolomics in tomato plants. Environmental and Experimental Botany, 2020, 179: 104187.
[10] Zhang M R, Sun Y K, Wen J, Liu Z Y, Yan S. Effects of melatonin on seedling growth, mineral nutrition, and nitrogen metabolism in cucumber under nitrate stress. Journal of Pineal Research, 2017, 62(4): e12403.
[11] 杜昕, 李博, 毛魯梟, 陳偉, 張玉先, 曹亮. 褪黑素對干旱脅迫下大豆產量及AsA-GSH循環的影響. 作物雜志, 2022(1): 174-178.
DU X, LI B, MAO L X, CHEN W, ZHANG Y X, CAO L. Effects of melatonin on yield and AsA-GSH cycle in soybean under drought stress. Crops, 2022(1): 174-178. (in Chinese)
[12] WANG P, SUN X, LI C, WEI Z W, LIANG D, MA F W. Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. Journal of Pineal Research, 2012, 54(3): 292-302.
[13] GONG B, YAN Y Y, WEN D, SHI Q H. Hydrogen peroxide produced by NADPH oxidase: a novel downstream signaling pathway in melatonin-induced stress tolerance in. Physiologia Plantarum, 2017, 160(4): 396-409.
[14] SUN Q Q, ZHANG N, WANG J F, ZHANG H J, LI D B, SHI J, LI R, WEEDA S, ZHAO B, REN S X, GUO Y D. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. Journal of Experimental Botany, 2015, 66(3): 657-668. doi: 10.1093/ jxb/eru332.
[15] 劉德帥, 姚磊, 徐偉榮, 馮美, 姚文孔. 褪黑素參與植物抗逆功能研究進展. 植物學報, 2022, 57(1): 111-126. doi: 10.11983/ CBB21146.
LIU D S, YAO L, XU W R, FENG M, YAO W K. Research progress of melatonin in plant stress resistance. Bulletin of Botany, 2022, 57(1): 111-126. doi: 10.11983/CBB21146. (in Chinese)
[16] 楊相東, 曹一平, 江榮風, 張福鎖. 幾種包膜控釋肥氮素釋放特性的評價. 植物營養與肥料學報, 2005, 11(4): 501-507. doi: 10.3321/ j.issn: 1008-505X.2005.04.012.
YANG X D, CAO Y P, JIANG R F, ZHANG F S. Evaluation of nutrients release feature of coated controlled-release fertilizer. Plant Nutrition and Fertilizer Science, 2005, 11(4): 501-507. doi: 10.3321/ j.issn: 1008-505X.2005.04.012. (in Chinese)
[17] YAN Y Y, JING X, TANG H, LI X, Gong B, SHI Q H. Using transcriptome to discover a novel melatonin-induced sodic alkaline stress resistant pathway inLPlant and Cell Physiology, 2019, 60(9): 2051-2064.
[18] 張彥才, 李若楠, 王麗英, 劉孟朝, 武雪萍, 吳會軍, 李銀坤. 磷肥對日光溫室番茄磷營養和產量及土壤酶活性的影響. 植物營養與肥料學報, 2008, 14(6): 1193-1199. doi: 10.3321/j.issn: 1008-505X. 2008.06.026.
ZHANG Y C, LI R N, WANG L Y, LIU M C, WU X P, WU H J, LI Y K. Effect of phosphorus fertilization on tomato phosporous nutrition, yield and soil enzyme activities. Plant Nutrition and Fertilizer Science, 2008, 14(6): 1193-1199. doi: 10.3321/j.issn: 1008-505X.2008.06.026. (in Chinese)
[19] 王秀峰. 蔬菜栽培學各論: 北方本. 4版. 北京: 中國農業出版社, 2011.
WANG X F. Various Theories of Vegetable Cultivation: Northern Edition. 4th ed. Beijing: China Agriculture Press, 2011. (in Chinese)
[20] 李愷, 張麗麗, 邵長勇, 仲崇山, 曹逼力, 史慶華, 鞏彪. 亞高溫下冷等離子體處理番茄種子對幼苗生長和光能利用的影響. 園藝學報, 2021, 48(11): 2286-2298. doi: 10.16420/j.issn.0513-353x. 2020-0698.
LI K, ZHANG L L, SHAO C Y, ZHONG C S, CAO B L, SHI Q H, GONG B. Effects of cold plasma seed treatment on tomato seedling growth and light energy utilization under daytime sub-high temperature environment. Acta Horticulturae Sinica, 2021, 48(11): 2286-2298. doi: 10.16420/j.issn.0513-353x.2020- 0698. (in Chinese)
[21] 丁紅, 張智猛, 戴良香, 楊吉順, 慈敦偉, 秦斐斐, 宋文武, 萬書波. 水氮互作對花生根系生長及產量的影響. 中國農業科學, 2015, 48(5): 872-881. doi: 10.3864/j.issn.0578-1752.2015.05.05.
DING H, ZHANG Z M, DAI L X, YANG J S, CI D W, QIN F F, SONG W W, WAN S B. Effects of water and nitrogen interaction on peanut root growth and yield. Scientia Agricultura Sinica, 2015, 48(5): 872-881. doi: 10.3864/j.issn.0578-1752.2015.05.05. (in Chinese)
[22] 張麗麗, 劉德興, 史慶華, 鞏彪. 黃腐酸對番茄幼苗適應低磷脅迫的生理調控作用. 中國農業科學, 2018, 51(8): 1547-1555. doi: 10.3864/j.issn.0578-1752.2018.08.012.
ZHANG L L, LIU D X, SHI Q H, GONG B. Physiological regulatory effects of fulvic acid on stress tolerance of tomato seedlings against phosphate deficiency. Scientia Agricultura Sinica, 2018, 51(8): 1547-1555. doi: 10.3864/j.issn.0578-1752.2018.08.012. (in Chinese)
[23] 張麗麗, 史慶華, 鞏彪. 中、堿性土壤條件下黃腐酸與磷肥配施對番茄生育和磷素利用率的影響. 中國農業科學, 2020, 53(17): 3567-3575. doi: 10.3864/j.issn.0578-1752.2020.17.013.
ZHANG L L, SHI Q H, GONG B. Application of fulvic acid and phosphorus fertilizer on tomato growth, development, and phosphorus utilization in neutral and alkaline soil. Scientia Agricultura Sinica, 2020, 53(17): 3567-3575. doi: 10.3864/j.issn.0578-1752.2020.17.013. (in Chinese)
[24] WANG B, GAO Z Y, SHI Q H, GONG B. SAMS1 stimulates tomato root growth and P availability via activating polyamines and ethylene synergetic signaling under low-P condition. Environmental and Experimental Botany, 2022, 197: 104844.
[25] HARDELAND R. Melatonin in plants and other phototrophs: Advances and gaps concerning the diversity of functions. Journal of Experimental Botany, 2015, 66(3): 627-646. doi: 10.1093/jxb/eru386.
[26] MADEBO M P, LUO S M, WANG L, ZHENG Y H, JIN P. Melatonin treatment induces chilling tolerance by regulating the contents of polyamine,-aminobutyric acid, and proline in cucumber fruit. Journal of Integrative Agriculture, 2021, 20(11): 3060-3074.
[27] 陳開, 唐瑭, 張冬平, 陳云, 呂冰. 生長素和細胞分裂素參與構建水稻根系的研究進展. 植物生理學報, 2020, 56(12): 2495-2509. doi: 10.13592/j.cnki.ppj.2020.0398.
CHEN K, TANG T, ZHANG D P, CHEN Y, Lü B. Recent advances in auxin-cytokinin interactions involved in shaping architecture of rice root system. Plant Physiology Journal, 2020, 56(12): 2495-2509. doi: 10.13592/j.cnki.ppj.2020.0398. (in Chinese)
[28] 高利娟, 張猛, 魏建林, 劉冬梅, 丁效東. 磷肥減施對設施番茄根系形態、磷吸收及土壤微生物量磷含量的影響. 天津農業科學, 2019, 25(8): 16-22. doi: 10.3969/j.issn.1006-6500.2019.08.004.
GAO L J, ZHANG M, WEI J L, LIU D M, DING X D. Effects of phosphorus fertilizer reduction on phosphorus uptake, root morphology, and microbial biomass phosphorus content in rhizosphere soil of tomato. Tianjin Agricultural Sciences, 2019, 25(8): 16-22. doi: 10.3969/j.issn.1006-6500.2019.08.004. (in Chinese)
[29] 臧祎娜, 張德閃, 李海港, 程凌云, 張朝春, 申建波. 褪黑素調控根系生長和根際互作的機制研究進展. 植物營養與肥料學報, 2019, 25(4): 671-682. doi: 10.11674/zwyf.18401.
ZANG Y N, ZHANG D S, LI H G, CHENG L Y, ZHANG C C, SHEN J B. Progress in mechanism of melatonin regulation of root growth and rhizosphere interactions. Plant Nutrition and Fertilizer Science, 2019, 25(4): 671-682. doi: 10.11674/zwyf.18401. (in Chinese)
[30] ZIA S F, BERKOWITZ O, BEDON F, WHELAN J, FRANKS A E, PLUMME K M. Direct comparison ofgene expression reveals different responses to melatonin versus auxin. BMC Plant Biology, 2019, 19: 567.
[31] YANG L, YOU J, LI J Z, WANG Y P, CHAN Z L. Melatonin promotesprimary root growth in an IAA-dependent manner. Journal of Experimental Botany, 2021, 72(15): 5599-5611. doi: 10.1093/jxb/erab196.
[32] BOZZO G G, DUNN E L, PLAXTON W C. Differential synthesis of phosphate-starvation inducible purple acid phosphatase isozymes in tomato () suspension cells and seedlings. Plant Cell & Environment, 2010, 29: 303-313.
[33] ZHANG Z X, HU Q, LIU Y N, CHENG P L, CHENG H, LIU W X, XING X J, GUAN Z Y, FANG W M, CHEN S M, JIANG J F, CHEN F D. Strigolactone represses the synthesis of melatonin, thereby inducing floral transition inin an FLC-dependent manner. Journal of Pineal Research, 2019, 67(7): e12582.
[34] ZHU X F, ZHU C Q, WANG C, DONG X Y, SHEN R F. Nitric oxide acts upstream of ethylene in cell wall phosphorus reutilization in phosphorus-deficient rice. Journal of Experimental Botany, 2017, 68(3): 753-760. doi: 10.1093/jxb/erw480.
Effects of Urea Slow-Release Functional Fertilizer Containing Melatonin on Growth, Yield and Phosphorus Use Efficiency of Tomato Under Reduced Phosphorus Application Conditions

1College of Horticultural Science and Engineering, Shandong Agricultural University/State Key Laboratory of Crop Biology, Tai’an 271018, Shandong;2College of Recourses and Environment, Shandong Agricultural University/National Engineering Research Center for Efficient Utilization of Soil and Fertilizer Resources, Tai’an 271018, Shandong
【Objective】Melatonin has multiple beneficial effects on plants. However, its active chemical properties limit its application in agricultural production. In this study, urea slow-release functional fertilizer containing melatonin (hereinafter referred to as functional fertilizer) was prepared by coating technology, and its effects on tomato growth, yield, and quality as well as phosphorus use efficiency were studied, so as to provide the theoretical basis for melatonin application with high use efficiency and reducing fertilizer rate on tomato.【Method】The release rate of melatonin in functional fertilizer was studied by water bubble method. Then the effects of melatonin-functional fertilizer on the growth of tomato seedlings were studied by plug seedling. Finally, a pot experiment was conducted to study the effects of functional fertilizer on plant growth and dry matter distribution, root growth, phosphorus absorption and distribution, phosphorus utilization rate and fertilizer yield contribution rate, root phosphatase activity, and fruit yield and quality. Four treatments were set, namely, phosphorus application (+P), non-phosphorus application (-P), urea slow-release fertilizer and functional fertilizer (-P+M and +P+M).【Result】The real melatonin content after coating was 35% of the total melatonin concentration when coating. When water bubbled to 60 d, the amount of melatonin residue in functional fertilizer was 6.61%. The application of functional fertilizer in conventional tomato seedlings could significantly promote seedling growth, and the seedling index was increased by 70.2% compared with the control. The pot experiment showed that the biomass of tomato roots, stems, leaves and fruits under -P treatment decreased by 19.64%, 18.51%, 28.99% and 28.73% compared with that under +P treatment, respectively. The dry matter distribution ratios of roots and stems increased by 10.03% and 11.63%, respectively, while the dry matter distribution ratios of leaves and fruits decreased by 2.74% and 2.39%, respectively. The application of functional fertilizer could significantly increase the biomass accumulation of all tissues under the two phosphorus treatments. The dry matter distribution ratio of root under the condition of increasing +P was 12.14%, the dry matter distribution ratio of stem and leaf under the condition of reducing -P was 6.00% and 5.90%, and the dry matter distribution ratio of fruit increased by 9.06%. -P treatment reduced the total length, total surface area and total volume of roots, and increased the number of root tips. The application of functional fertilizer could increase the total length, total surface area, total volume and number of root tips under two phosphorus levels. -P treatment significantly reduced the phosphorus content in roots, stems, leaves and fruits of tomato plants, increased the phosphorus distribution in roots, stems and leaves, and reduced the phosphorus distribution ratio in fruits. The application of functional fertilizer could significantly increase the phosphorus content of all tissues under the two P treatments, increase the phosphorus distribution ratio of roots under +P treatments, decrease the phosphorus distribution ratio of stems and leaves under -P treatment, and increase the phosphorus distribution ratio of fruits. The application of functional fertilizer could significantly improve phosphatase activity, whole plant phosphorus uptake, fertilizer utilization rate and fertilizer yield contribution rate. -P treatment reduced tomato yield, but application of melatonin functional fertilizer significantly increased yield under -P treatment reduced the tomato yield by 17.57%, but the application of functional fertilizer under -P treatment increased the yield by 21.32%, and had no significant effect on the yield under +P treatment. In addition, the application of functional fertilizer could comprehensively improve the quality of tomato fruits under -P or partial +P conditions.【Conclusion】The application of melatonin coated in slow-release fertilizer could significantly reduce the amount of melatonin, improve the growth quality of tomato seedlings and plants in the whole growth period, increase the utilization efficiency of phosphorus fertilizer, and finally improve the yield and fruit quality.
tomato; urea slow-release functional fertilizer containing melatonin; phosphate fertilizer treatment; growth; yield and quality; phosphorus utilization rate

10.3864/j.issn.0578-1752.2023.03.010
2022-04-06;
2022-06-06
國家自然科學基金(U1903105)
劉明慧,E-mail:liuminghuiaq@163.com。通信作者鞏彪,E-mail:gongbiao@sdau.edu.cn
(責任編輯 趙伶俐)
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
小讀者(2021年2期)2021-03-29 05:03:48
今日農業(2020年20期)2020-11-26 06:09:10
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
