非釀酒酵母屬酵母的葡萄酒發酵應用研究進展
2023-03-07 02:29:38王春曉俞俊竹周文亞許引虎
中國農業科學 2023年3期
關鍵詞:高產
王春曉,俞俊竹,周文亞,許引虎
非釀酒酵母屬酵母的葡萄酒發酵應用研究進展
王春曉1,俞俊竹1,周文亞1,許引虎2
1貴州大學釀酒與食品工程學院/貴州省發酵工程與生物制藥重點實驗室,貴陽 550025;2國家酵母技術研究推廣中心,湖北宜昌 443003
葡萄皮上天然存在著非釀酒酵母屬酵母(non),主要在葡萄酒浸漬和發酵初期發揮作用,近年來非釀酒酵母屬酵母在葡萄酒發酵中的應用受到越來越多的關注。相對于釀酒酵母,非釀酒酵母屬酵母在酒精發酵中具有較弱的發酵力,可將還原糖轉化為乙醇及其他代謝副產物,是生產復雜風味和低酒度葡萄酒的潛在優良酵母。不同非釀酒酵母屬酵母菌種在葡萄酒發酵應用中具有不同的代謝特征,選擇具有一定特征的優良非釀酒酵母屬酵母應用于發酵中,可以提高葡萄酒的特色化品質。本研究在總結商業化非釀酒酵母屬酵母的種類、釀造特點和應用方式的基礎上,重點綜述了不同非釀酒酵母屬酵母對葡萄酒顏色、香氣、口感和安全健康4個方面的積極作用、代謝機理和研究熱點:具有高產酸、多糖、胞外丙酮酸及低吸附性等特性的非釀酒酵母屬酵母可通過不同代謝機理促進葡萄酒顏色的穩定;不同非釀酒酵母屬酵母通過低產乙醇、乙醛、降低揮發性酚類,高產乙酸乙酯、乙酸酯類化合物、乙酯類化合物、高級醇、與萜烯或硫醇釋放相關的酶類等途徑促進葡萄酒果味香氣的提升,增加香氣復雜性;非釀酒酵母屬酵母通過高產甘油、多糖和乳酸,降解蘋果酸等方式調節葡萄酒的口感特征;非釀酒酵母屬酵母作為生物防治劑可以降低葡萄酒釀造中二氧化硫的用量,通過代謝降解作用減少有毒化合物,提升葡萄酒的安全質量。本文進一步解析了非釀酒酵母屬酵母的基因組和微衛星位點分析研究現狀,探討了目前非釀酒酵母屬酵母葡萄酒發酵應用研究的主要接種策略,提出了未來研究仍需關注的熱點方向,為非釀酒酵母屬酵母在葡萄酒酒精發酵中的應用研究提供理論參考。
混合發酵;顏色穩定性;香氣復雜性;基因型;商業化
0 引言
葡萄酒釀造是一個復雜的生物化學過程,主要由酵母菌完成酒精發酵。非釀酒酵母屬(non-)是相對于釀酒酵母屬()提出的一個概念,主要是指在葡萄酒釀造中存在的除釀酒酵母屬之外的所有酵母菌種[1]。Kurtzman等[2]鑒定了149個屬的將近1 500個非釀酒酵母屬菌種,然而據Jolly等[1]2014年的統計只有40余種分離自葡萄或葡萄醪,這一數字在2021年更新至293個菌種[3]。293個非釀酒酵母屬菌種在分類地位上屬于2個門、9個綱、18個目和37個科,其中191個非釀酒酵母屬菌種屬于子囊菌門(Ascomycota),101個菌種屬于擔子菌門(Basidiomycota),由于非釀酒酵母屬酵母大多高產揮發酸,曾經一度被認為是葡萄酒釀造的敗壞酵母[3-4]。近年來隨著產區特色葡萄酒、自然葡萄酒、有機葡萄酒、生物動力葡萄酒等在市場推廣和全球變暖對產區風土條件的影響[5–7],具有不同風土特色的葡萄酒越來越受到人們的關注,因此,一部分具有某些釀酒特征的非釀酒酵母屬酵母得以篩選和商業化(表1),并主要以與釀酒酵母結合應用的方式賦予葡萄酒獨特的風格[5,8-9]。
按照發酵能力和耐受性的強弱,非釀酒酵母屬酵母可以分為兩類(圖1)[10]:一類為發酵型酵母,擁有較強的產酒精能力和耐受性,因此具備單菌種發酵的潛力,但目前仍主要以與釀酒酵母結合應用的方式提升葡萄酒的風味復雜性和釀造可控性[11-12]。戴爾有孢圓酵母()、粟酒裂殖酵母()和耐熱克魯維酵母()是目前主要商業化的發酵型非釀酒酵母屬酵母(表1)。另一類為氧化發酵型酵母(也稱為半發酵型酵母),產酒精能力和耐受性一般弱于釀酒酵母,單菌種發酵往往導致發酵遲緩或發酵中止,無法將葡萄醪中主要的葡萄糖和果糖代謝完全,因此需與代謝力強的釀酒酵母或發酵型非釀酒酵母屬酵母結合使用來完成酒精發酵。目前商業化的氧化發酵型酵母主要包括美極梅奇酵母()、克魯維畢赤酵母()和瑟氏哈薩克斯坦酵母()等(表1),葡萄酒有孢漢遜酵母()等其他具有代謝特色的酵母受到日益增加的研究關注,具有商業化應用的潛力。非釀酒酵母屬酵母的研發推動了混合接種發酵方式在葡萄酒釀造工業中的應用,部分商業酵母為釀酒酵母與1—2種非釀酒酵母屬酵母的混合菌劑(如Melody,表1)或非釀酒酵母屬酵母之間的混合菌劑(如Thrill yeast,表1)。
不同非釀酒酵母屬酵母菌種在葡萄酒釀造過程中所起的作用各具特色,歸納總結其代謝機理、發酵特征,對在葡萄酒釀造接種過程中根據葡萄酒釀造特性要求,選擇相應的非釀酒酵母屬酵母具有重要意義。本研究綜述了可積極改善葡萄酒品質的非釀酒酵母屬酵母菌種及其代謝機理和混合發酵應用研究熱點,并總結了非釀酒酵母屬酵母基因組和種內基因型分析方法(微衛星位點分析)的研究現狀,為非釀酒酵母屬酵母在葡萄酒發酵中的應用提供理論基礎,為釀造多樣化和特色化的葡萄酒提供理論參考。
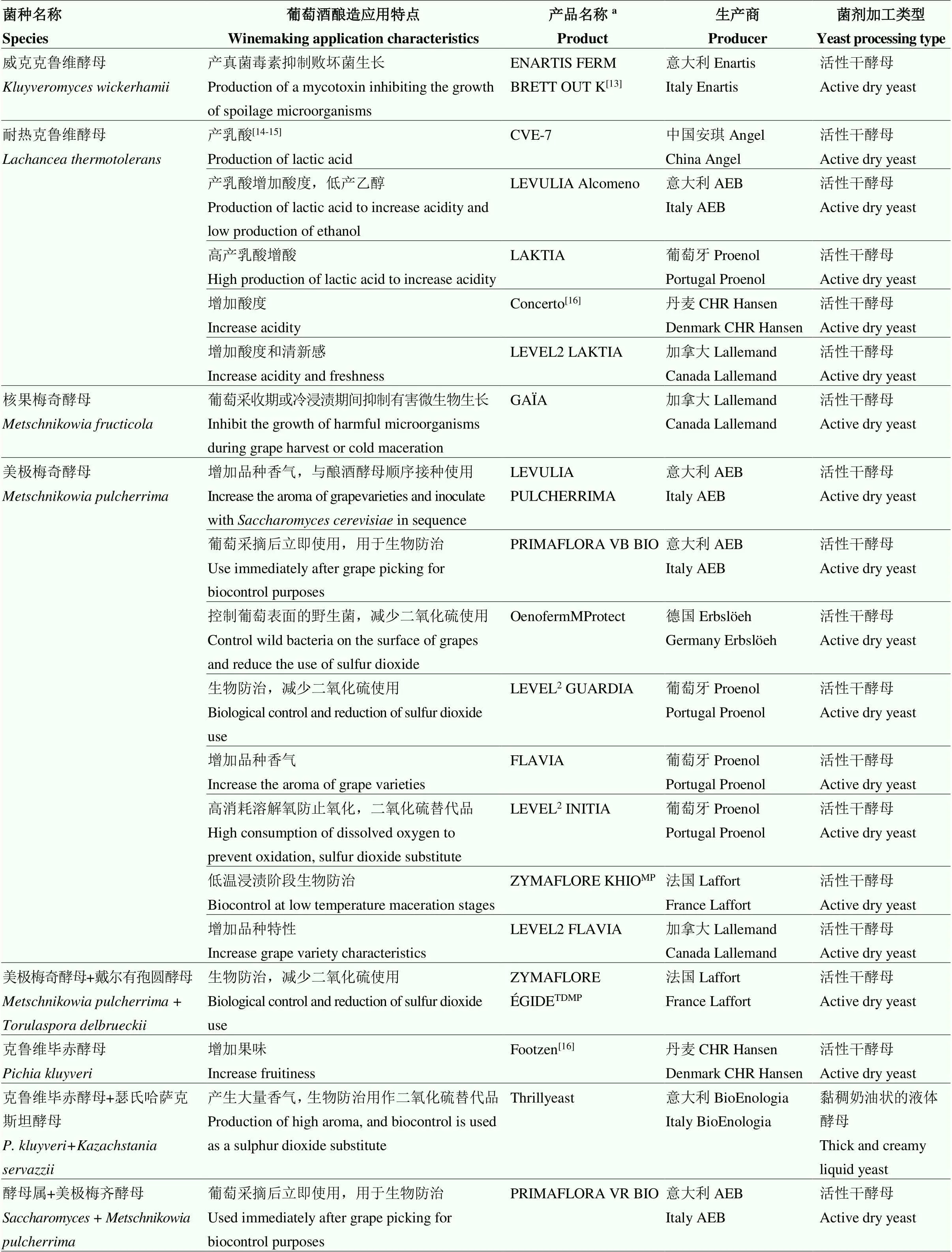
表1 已經商業化應用于葡萄酒釀造領域的非釀酒酵母屬酵母
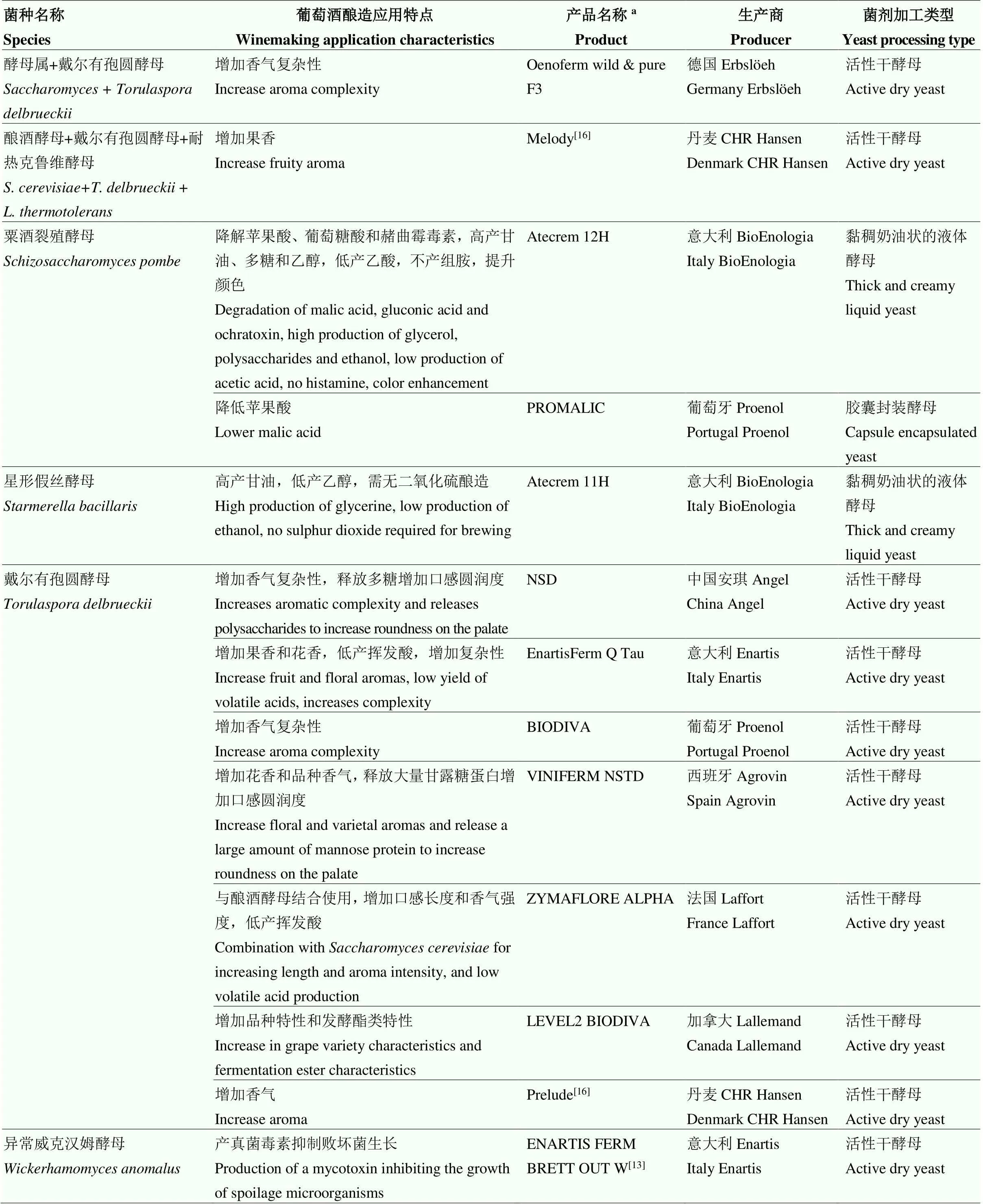
續表1 Continued table 1
a產品名稱源自參考文獻或網址(2022年2月4日檢索結果):加拿大Lallemand(https://www.lallemandwine.com/)、意大利Enartis(https://www. enartis.com/)、法國Laffort(https://laffort.com/)、西班牙Agrovin(https://www.agrovin.com/)、葡萄牙Proenol(https://www.proenol.com/web/)、意大利BioEnologia(https://www.bioenologia.com/vino/)、德國Erbsl?eh(https://erbsloeh.com/)、意大利AEB(https://www.enartis.com/)
aProduct name was derived from reference or website (search results on February 4, 2022): Canada Lallemand (https://www.lallemandwine.com/), Italy Enartis (https://www.enartis.com/), France Laffort (https://laffort.com/), Spain Agrovin (https://www.agrovin.com/), Portugal Proenol (https://www.proenol.com/web/), Italy BioEnologia (https://www.bioenologia.com/vino/), Germany Erbsl?eh (https://erbsloeh.com/), Italy AEB (https://www.enartis.com/)
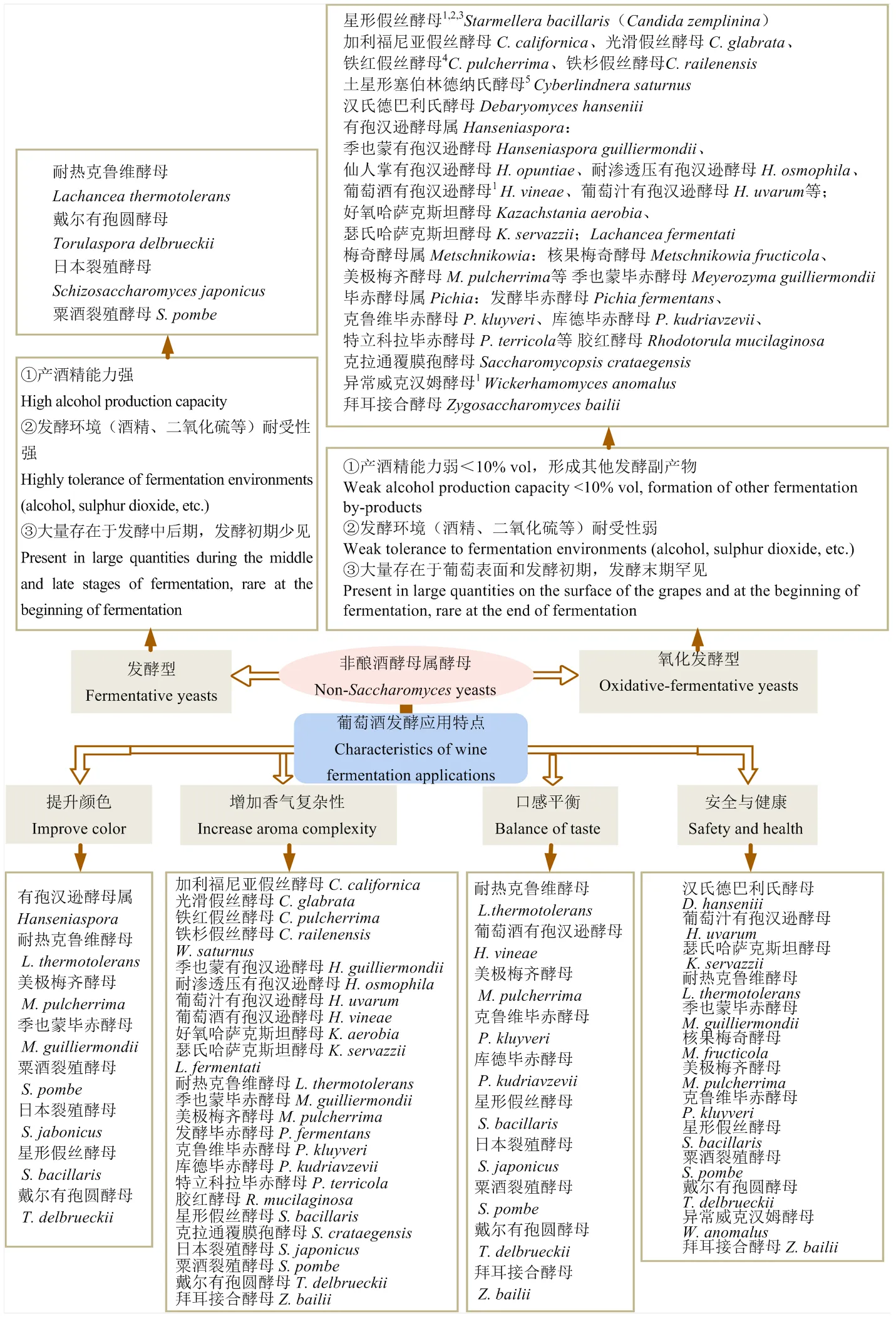
1有些菌株耐受10% vol左右酒精;22011年星形假絲酵母Candida stellata被重新歸類為澤普林假絲酵母Candida zemplinina[1],2012年澤普林假絲酵母C. zemplinina更名為星形假絲酵母Starmerellabacillaris[21];3有些菌株產酒精能力強(11%—13% vol);4是美極梅齊酵母的無性型;5同土星擬威爾酵母
非釀酒酵母屬酵母與釀酒酵母的結合應用促進了葡萄酒顏色、香氣、口感和安全健康等方面的品質提升(圖1):①非釀酒酵母屬酵母通過5種不同的代謝機理改善花青素的聚合狀態,從而提升葡萄酒顏色;②非釀酒酵母屬酵母通過7種發酵相關代謝途徑改善葡萄酒香氣復雜性;③非釀酒酵母屬酵母通過4種發酵副產物影響葡萄酒口感平衡;④非釀酒酵母通過3方面代謝提升葡萄酒的安全性與健康。
1 可提升葡萄酒顏色的非釀酒酵母屬酵母
葡萄酒中的花青素源自葡萄果皮,主要有5種:矢車菊素、芍藥素、飛燕草素、矮牽牛素和錦葵素[22]。在葡萄酒發酵和陳釀過程中,花青素經過一系列聚合作用不斷變化,形成陳年葡萄酒中賦予顏色穩定性的花青素化合物,如吡喃型花青素類化合物[23]。吡喃型花青素類化合物包含多種類型,如vitisin A型、vitisin B型、portisin型和pinotins型等(圖2),其中vitisin A型是陳年葡萄酒中含量最豐富的吡喃型花青素,其顏色深、穩定性好且耐二氧化硫漂白,因此,vitisin A型花青素的增加可提升葡萄酒顏色穩定性,而選擇高產丙酮酸和乙醛的非釀酒酵母屬酵母能提高葡萄酒中vitisins型吡喃花青素含量[24]。葡萄酒中vitisin A型花青素提供了橙紅色,vitisin B型花青素提供了橘黃色,portisin型花青素則提供了藍色、藍紫色與紫紅色,在發酵初期由于丙酮酸的生成,vitisin A型花青素累積速度高于vitisin B型花青素和portisin型花青素,多種吡喃型花青素在葡萄酒發酵過程中累積,提高了顏色穩定性[23,25]。
研究表明,高產丙酮酸[26],單菌種酒精發酵結束后發酵液中丙酮酸含量為287 mg·L-1,而釀酒酵母為89 mg·L-1[27],因為可通過蘋果酸酒精發酵途徑將蘋果酸分解形成丙酮酸,從而增加胞外丙酮酸的含量[28]。釋放到胞外的丙酮酸與花青素反應形成vitisin A型花青素(圖2),因此,純種發酵和混合發酵都增加了顏色強度、vistin A型花青素和總花青素的含量[27],且混合發酵時增加的接種比例能夠加強紅色色澤[29]。值得注意的是,丙酮酸含量并非越高越好,在模擬葡萄汁中加入880.6 mg·L-1的丙酮酸會對釀酒酵母形成壓力,促進部分花青素轉化形成山柰酚及其衍生物,最終造成顏色的損失[24]。因此,在篩選高產丙酮酸酵母時需綜合分析其丙酮酸產量及對顏色的影響。除外,也具有蘋果酸酒精發酵途徑,研究表明,與釀酒酵母同時接種發酵對顏色的提升弱于順序接種發酵(接種24 h后再接種釀酒酵母)[30]。接種后48—72 h再接種能顯著增加葡萄酒中vistin A型花青素的含量,推斷可能與在酒精發酵過程中高產丙酮酸有關,但仍需證實[31]。除vitisin A型花青素外,酵母菌酒精發酵過程中形成的乙醛能形成乙基橋(CH-CH3)將單體花青素與黃烷醇縮合形成更穩定的聚合色素——vitisin B型花青素[23,32]。葡萄酒中丙酮酸和乙醛的含量決定了vitisin A型與vitisin B型含量。在酵母菌相應酶的催化作用下,vitisin A型花青素還可以進一步與乙烯基黃烷-3-醇、羥基肉桂酸、乙烯基苯酚反應形成portisin型花青素(圖2)。季也蒙畢赤酵母()具有最高的羥基肉桂酸脫羧酶活性,可以產生pinotins型花青素。
非釀酒酵母屬酵母還可通過降低pH、細胞壁的低吸附性、促進花青素和黃烷醇低聚物的形成、促進或調解單寧與花色素之間的聚合等方式提升葡萄酒顏色(圖2)。可代謝丙酮酸形成大量乳酸[14-15,33],降低葡萄酒pH,從而提升葡萄酒的顏色強度和穩定性。酵母菌細胞壁上球狀的甘露糖蛋白更易與酚類物質發生相互作用,因此對花青素具有一定的吸附作用[34]。與釀酒酵母和分別混合發酵釀造的葡萄酒中花青素的含量高于這兩個菌單獨發酵所釀葡萄酒中的花青素含量,說明對花青素的吸附性較弱[27]。、、可促進花青素和黃烷醇形成低聚物,從而提升葡萄酒顏色[28,31]。高產的多糖(混合發酵產生多糖的含量是釀酒酵母發酵產生多糖的2倍多)可能通過疏水作用和氫鍵與單寧結合,促進單寧與花色素聚合物的穩定性,而高產的乙醛也可以調節單寧與花色素之間的聚合反應[30]。
非釀酒酵母屬酵母對葡萄酒顏色提升的研究目前集中于8個菌種(圖1),它們提升葡萄酒顏色的機理各不相同,且需要從發酵、陳釀等工藝環節評價顏色的穩定性,增加了研究復雜性。這8種可提升葡萄酒顏色的非釀酒酵母屬酵母在應用時需注意評價不同接種方式(同時接種、順序接種、接種比例)對顏色提升的影響,獲取所需最佳接種方案。若需要篩選提升葡萄酒顏色的非釀酒酵母屬酵母可關注發酵過程是否高產丙酮酸或乙醛、是否具有高羥基肉桂酸脫羧酶活性或低吸附性、是否降低pH、是否促進花青素和黃烷醇低聚物的形成等。而發酵產物如多糖、乙醛等對陳釀過程中單寧與花色素聚合物的促進機理還需進一步研究解析。
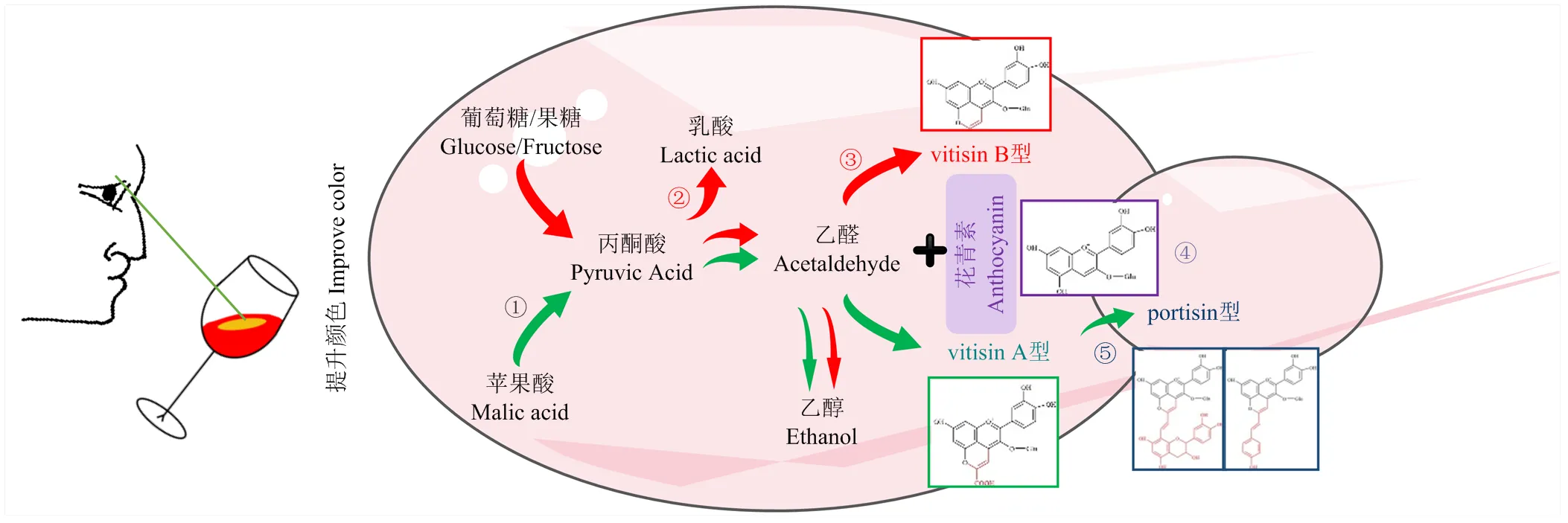
以矢車菊素為例展現花青素及吡喃型花青素類化合物的結構式。①釋放胞外丙酮酸,形成vitisin A型:日本裂殖酵母、粟酒裂殖酵母;②高產乳酸,降低葡萄酒pH值,提升顏色強度和穩定性:耐熱克魯維酵母;③釋放胞外乙醛,形成vitisin B型:釀酒酵母等;④細胞壁對花青素的低吸附性:耐熱克魯維酵母;促進花青素和黃烷醇形成低聚物:美極梅齊酵母、星形假絲酵母、戴爾有孢圓酵母;⑤vitisin A型進一步與乙烯基黃烷-3-醇/羥基肉桂酸/乙烯基苯酚形成portisin型:日本裂殖酵母、粟酒裂殖酵母
2 可增加葡萄酒香氣復雜性的非釀酒酵母屬酵母
許多非釀酒酵母屬酵母引起研究者的關注是因為它們對葡萄酒香氣復雜性的貢獻[19],尤其是品嘗時可感知的果香等香氣,如貢獻于草莓和獼猴桃香氣(甲基丙酸乙酯)及黑莓香氣(甲基丙酸乙酯)的[35],貢獻于烘烤焦糖和干果果仁香氣的[36],貢獻于瓊瑤漿典型性香氣的[37],貢獻于甜香和花香的克拉通覆膜孢酵母()和[38]等。目前關于非釀酒酵母屬酵母對葡萄酒香氣特征的影響研究是葡萄酒發酵中非釀酒酵母屬酵母的主要研究內容之一。與葡萄酒果味直接相關的化合物主要包括酯類、揮發性硫醇、內酯、呋喃酮、高級醇等,有些化合物如乙酸和乙酸乙酯可通過進一步的化學反應形成與果味直接相關的化合物[35],有些化合物的濃度高低會對果味的感知產生不同的影響,如低乙醇含量促進果味感知[10]。因此,本研究主要分析了可以增加葡萄酒香氣復雜性的非釀酒酵母屬酵母及主要代謝機理(圖3),包括低產乙醇、高產乙酸乙酯、高產乙酸酯類化合物、高產乙酯類化合物、高/低產高級醇類化合物、高產與萜烯或硫醇釋放相關的酶類、高/低產乙醛、降低揮發性酚類。
2.1 產乙醇能力
由于全球變暖對產區風土條件的影響導致成熟葡萄的含糖量增加,因此所釀造葡萄酒的酒度提升,高酒度會掩蓋果味[6],而低乙醇含量促進果味的感知[10]。因此,優選非釀酒酵母屬酵母、、和被用來與釀酒酵母開展混合發酵,實現低殘糖和低乙醇的釀造目標。
2.2 產乙酸乙酯能力
乙酸乙酯的含量超過12 mg·L-1會產生不愉快氣味[9],超過200 mg·L-1會產生敗壞風味[44],而很多可以增加香氣復雜性的非釀酒酵母屬酵母同時也高產乙酸乙酯,如圖3所示。有8種酵母為高產乙酸乙酯酵母,而的乙酸乙酯產量具有菌株差異。近年來,部分研究指出乙酸乙酯可能會通過轉酯作用形成酯類[55];與釀酒酵母混合發酵結束后,乙酸乙酯含量為600—700 mg·L-1經過,24個月陳釀后降為140—170 mg·L-1,乙酸乙酯在葡萄酒陳釀期間的降低可能與水解或酯化反應有關[30]。因此,酒精發酵結束后的高乙酸乙酯似乎充當了葡萄酒陳釀過程中揮發性酯類化合物演化的儲存庫,而高產乙酸乙酯酵母可考慮篩選用于陳釀型葡萄酒釀造中。
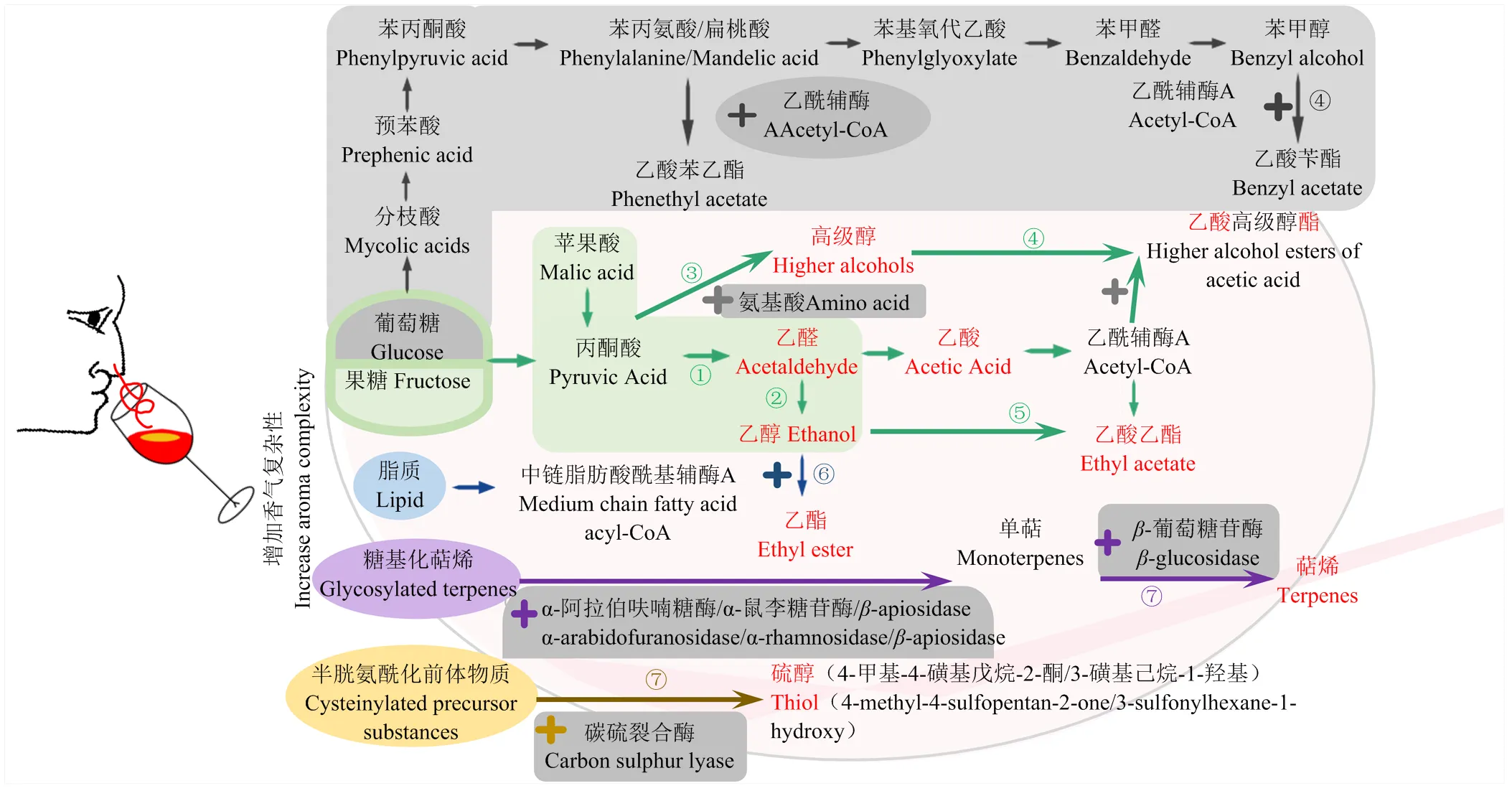
①高產乙醛:日本裂殖酵母、粟酒裂殖酵母、星形假絲酵母;低產乙醛、乙酸或3-羥基丁烷-2-酮:耐熱克魯維酵母、L. fermentati、美極梅奇酵母、特立科拉畢赤酵母、庫德畢赤酵母、星形假絲酵母、戴爾有孢圓酵母;②降低乙醇:耐熱克魯維酵母、美極梅奇酵母、星形假絲酵母、戴爾有孢圓酵母;③高產高級醇類化合物(Ehrlich途徑、糖代謝合成途徑):葡萄酒有孢漢遜酵母、季也蒙畢赤酵母、美極梅奇酵母、發酵畢赤酵母、膠紅酵母、日本裂殖酵母;低產高級醇類化合物:有孢漢遜酵母屬、星形假絲酵母、粟酒裂殖酵母、接合酵母屬;④高產乙酸酯類化合物:加利福尼亞假絲酵母、土星形塞伯林德納氏酵母、季也蒙有孢漢遜酵母、耐滲透壓有孢漢遜酵母、葡萄汁有孢漢遜酵母、有孢漢遜酵母屬、好氧哈薩克斯坦酵母、瑟氏哈薩克斯坦酵母、美極梅奇酵母、克魯維畢赤酵母、特立科拉畢赤酵母、庫德畢赤酵母;葡萄酒有孢漢遜酵母通過分枝酸-預苯酸途徑高產乙酸酯類化合物;⑤高產乙酸乙酯:季也蒙有孢漢遜酵母、葡萄酒有孢漢遜酵母、葡萄汁有孢漢遜酵母、特立科拉畢赤酵母、庫德畢赤酵母、克魯維畢赤酵母、日本裂殖酵母、拜耳接合酵母;⑥高產乙酯類化合物:有孢漢遜酵母屬、葡萄汁有孢漢遜酵母、克魯維畢赤酵母、庫德畢赤酵母、特立科拉畢赤酵母、克拉通覆膜孢酵母、星形假絲酵母、戴爾有孢圓酵母、拜耳接合酵母;⑦高產酶類,釋放萜烯類化合物或品種硫醇等:加利福尼亞假絲酵母、鐵紅假絲酵母、有孢漢遜酵母屬、葡萄汁有孢漢遜酵母、耐熱克魯維酵母、美極梅奇酵母、克魯維畢赤酵母、庫德畢赤酵母、膠紅酵母、星形假絲酵母、克拉通覆膜孢酵母、戴爾有孢圓酵母
2.3 產乙酸酯能力
除乙酸乙酯之外的其他乙酸酯大多由Ehrlich途徑或糖代謝合成途徑形成的高級醇與乙酸酯化形成,研究發現高產乙酸酯類的非釀酒酵母屬酵母通常只高產其中某些乙酸酯,這可能與酵母菌的特殊代謝途徑有關,如通過分枝酸-預苯酸途徑生成乙酸苯乙酯和乙酸芐酯(圖3)。非釀酒酵母屬酵母對氮源的消耗與產生的香氣種類相關,在模擬汁中分別加入24種不同氮源作為單一氮源,總酵母可同化氮含量控制在200 mg·L-1時發現、和的單菌種發酵雖然體現了氨基酸利用和所產香氣的差異,但是不論氮源是什么,這3個菌種都可以產生所有的發酵類香氣(高級醇和酸、中鏈脂肪酸和它們的乙酸或乙酯類衍生物),說明了中心碳代謝(如糖代謝合成途徑)對香氣的重要作用[46]。高產乙酸酯的非釀酒酵母屬酵母除了包含大家熟知的、和外,最近有研究表明和可增加乙酸酯含量,尤其是乙酸苯乙酯和乙酸異戊酯含量,賦予了西拉紅葡萄酒更高的果醬和紅色水果香氣[17]。
2.4 產酯能力
酯類通常給葡萄酒帶來果香和花香,非釀酒酵母屬酵母不但可以通過高級醇和乙酸之間的酯化作用形成乙酸高級醇酯,還可以通過脂質代謝形成的中鏈脂肪酸與乙醇酯化形成乙酯類化合物,目前高產乙酯類的非釀酒酵母屬酵母有9種,其中、、和為高產乙酸酯和乙酯類化合物的菌種(圖3)。拜爾結合酵母()因高產酯酶,在與釀酒酵母混合發酵中能產生大量乙酯如乙酸乙酯、己酸乙酯、辛酸乙酯和癸酸乙酯等[54]。中國本土非釀酒酵母屬酵母和共同與釀酒酵母進行順序接種發酵,增加了葡萄酒中大部分乙酯的濃度,貢獻了菠蘿香氣[56]。
2.5 產高級醇能力
葡萄酒中高級醇含量高于300 mg·L-1時會產生不愉快的味道[30],超過400 mg·L-1會產生香氣負面效應[44]。低含量的高級醇通常賦予葡萄酒果香,研究表明高產高級醇的菌種包含6種,低產高級醇的菌種包含4種(圖3),由于高級醇可以進一步酯化形成乙酸酯,因此,不建議簡單通過酒精發酵結束后高級醇的含量來篩選非釀酒酵母屬酵母。此外,有些酵母菌的高級醇產量呈現明顯的菌株差異,如在不同的研究報道中有些菌株高產高級醇,有些低產[9]。
2.6 產酶能力
目前發現12種非釀酒酵母屬酵母因高產酶類增加品種香氣或典型性,部分酵母是高產相關糖苷酶釋放萜烯類化合物如和[45],而有些酵母是高產碳硫裂合酶形成揮發性硫醇[57],產生熱帶水果的品種香氣,如[9]。因此,非釀酒酵母屬酵母篩選中通常會檢測-葡萄糖苷酶等相關酶的活性[58],和高產-D-木糖苷酶活性,和高產-L-鼠李糖苷酶和-L-阿拉伯糖苷酶[59-60]。研究人員從中國河北葡萄酒產區篩選了高產-葡萄糖苷酶的膠紅酵母(),在與釀酒酵母順序接種發酵中,該酵母顯著增加了萜烯類和-大馬酮含量,帶給葡萄酒強烈的花香和果香味[48],且隨著接種比例增加而增加[49-50]。
2.7 產乙醛能力
葡萄酒中乙醛的感知閾值是100 mg·L-1,高含量乙醛會給葡萄酒帶來氧化味[30]。因具有蘋果酸酒精發酵途徑而高產乙醛、乙醇和乙酸乙酯(圖3),研究發現與釀酒酵母酒精發酵剛結束時乙醛含量為18—36 mg·L-1,其中混合發酵高于釀酒酵母純種發酵,順序接種混合高于同時接種混合,而經過24個月陳釀后,幾種發酵之間差異不大,降至7—8 mg·L-1,說明乙醛在陳釀過程中發生了進一步轉化[30]。
2.8 降低揮發性酚
揮發性酚濃度低時呈現煙熏味,濃度高時呈現汗味[10],因此,可降低揮發性酚類的非釀酒酵母屬酵母如spp獲取研究者的關注[43]。
研究人員早期對非釀酒酵母屬酵母的研究多開展以模擬汁或葡萄汁為原料的非釀酒酵母屬酵母純種發酵試驗,如在模擬汁純種發酵中可產生8—19 mg·L-1乙偶姻,在葡萄汁發酵中可產生17.1—34.4 mg·L-1乙偶姻,而不產乙偶姻[61]。研究發現大多非釀酒酵母屬酵母的產乙醇能力弱于釀酒酵母,但可產大量發酵副產物,對葡萄酒的香氣產生積極的影響。因此,近年來對非釀酒酵母屬酵母的研究大多直接對比混合發酵與釀酒酵母純種發酵之間的差異,混合發酵常用兩種接種方式,同時混合接種方式(將非釀酒酵母屬酵母和釀酒酵母以一定的比例接種,一般為1﹕1或非釀酒酵母屬比例更高)和順序接種方式(先接種非釀酒酵母屬酵母,接種后24、48或72 h后再接種釀酒酵母),由于順序接種方式給予非釀酒酵母屬酵母較充分的代謝時間,因此獲取的香氣復雜性和感官評價往往高于同時混合接種方式,而混合發酵方式在香氣改善方面往往優于釀酒酵母純種發酵[58,62-63]。部分研究還嘗試應用2種以上非釀酒酵母屬酵母與釀酒酵母開展混合發酵[39]。因此,圖3中高產或低產的結論大多獲取自非釀酒酵母屬酵母在與釀酒酵母混合發酵時相對于釀酒酵母純種發酵時的表現,研究結論受所用葡萄品種(是否富含糖基化萜烯或半胱氨酰化前體物質)、非釀酒酵母屬酵母菌株(純種發酵是否提升香氣復雜性)、對照釀酒酵母菌株(是否嗜殺、是否提升香氣復雜性)等因素的影響,不同研究所獲取的非釀酒酵母屬酵母增加香氣復雜性的結論之間往往無法直接對比評價。建議設定數值化參數標準或開展同一模擬汁條件下發酵試驗,以便于橫向比較非釀酒酵母屬酵母的香氣改善能力,提升高產或低產結論判斷的準確性。
3 可改善葡萄酒口感平衡的非釀酒酵母屬酵母
葡萄酒的主要成分是來自于葡萄漿果的水分(70% —90%),其次是乙醇(7%—17%),這兩種物質是葡萄酒口感平衡物質的主要載體[22]。而甘油、多糖和乳酸是酵母菌酒精發酵代謝產生的可影響葡萄酒口感平衡的主要物質(圖4)。
3.1 產甘油和多糖能力
甘油是最主要的酒精發酵副產物,釀酒酵母可發酵形成一定量的甘油,非釀酒酵母屬酵母可能比釀酒酵母多產零點幾到4 g·L-1的甘油,從生物化學的角度來看,非釀酒酵母屬酵母擁有較弱的乙醇脫氫酶活性,但是有更強的3-磷酸甘油脫氫酶活性[9]。目前已報道有6種非釀酒酵母屬酵母高產甘油如,在與釀酒酵母的混合發酵過程中提升甘油產量,增加葡萄酒的柔和度和酒體。、和既可以提升甘油,又可以在發酵過程中釋放多糖,增加的甘油和多糖會提高口感的甜度,降低收斂性[30]。研究表明,葡萄酒中的多糖來源于葡萄(阿拉伯糖蛋白)或酵母菌的細胞壁成分(甘露糖蛋白),酵母菌細胞壁最外面存在甘露糖蛋白,由85%—90%的甘露糖和10%—15%的蛋白組成。酵母在出芽生長的過程中通過控制性水解母細胞的細胞壁可釋放甘露糖蛋白,因此,非釀酒酵母屬酵母的釀造應用特征之一是釋放更多的多糖[9,64]。在發酵過程中釋放的多糖成分主要為半乳糖-甘露糖蛋白,與其他酵母釋放的甘露糖蛋白一樣,可以與多酚類物質如單寧聚合,降低收斂性[65],其在混合發酵產生的多糖是釀酒酵母單菌種發酵的2倍多[30]。酵母多糖除了可改善口感平衡,還可提升葡萄酒蛋白和酒石酸鹽的穩定性,保留葡萄酒香氣物質[30,65]。
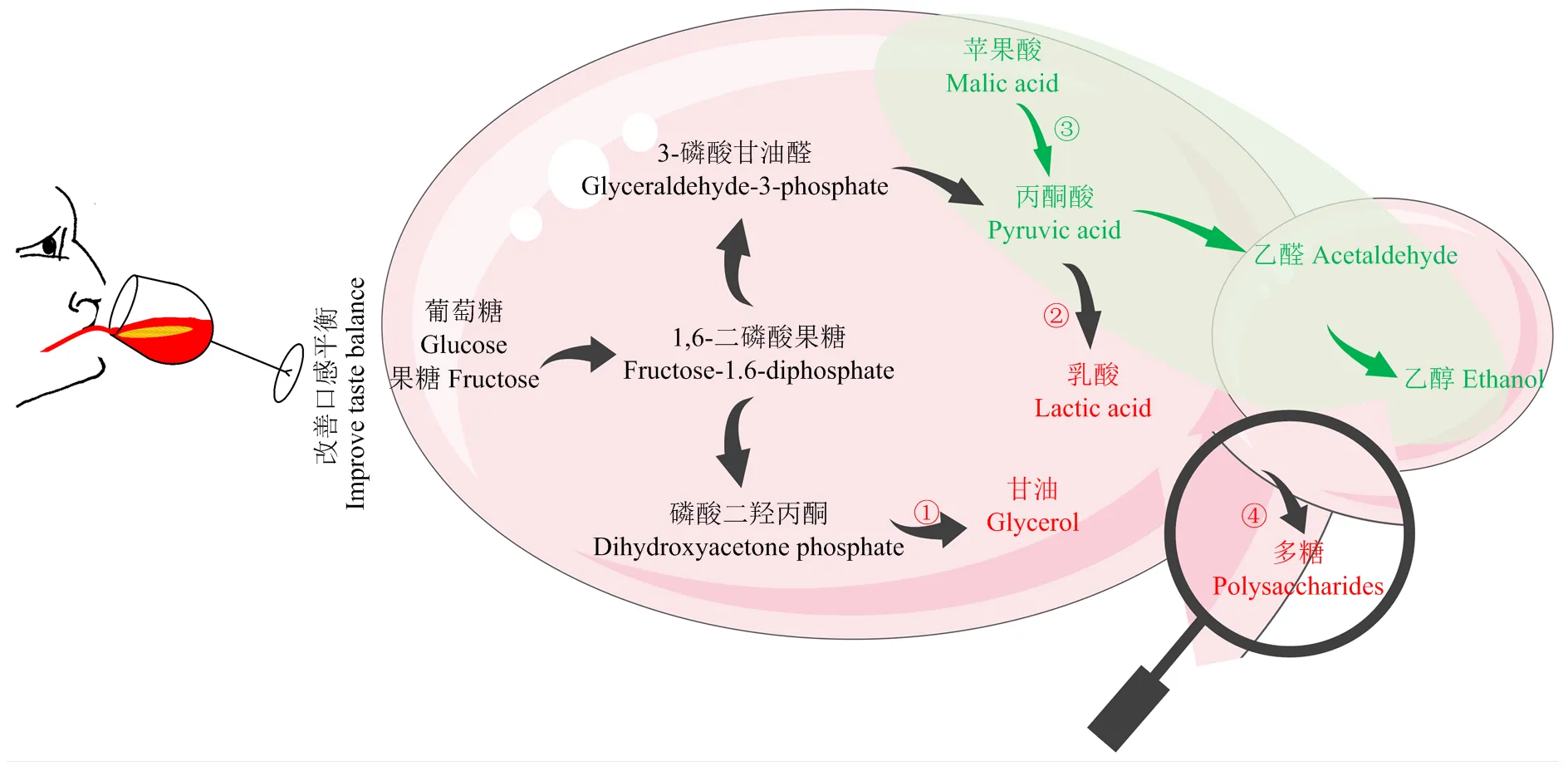
①高產甘油:耐熱克魯維酵母、美極梅齊酵母、克魯維畢赤酵母、庫德畢赤酵母、星形假絲酵母、日本裂殖酵母、戴爾有孢圓酵母;②產生乳酸:耐熱克魯維酵母;③降低蘋果酸(蘋果酸酒精發酵):美極梅齊酵母、庫德畢赤酵母、日本裂殖酵母、粟酒裂殖酵母、戴爾有孢圓酵母;④產生多糖如甘露糖蛋白等:耐熱克魯維酵母、粟酒裂殖酵母、日本裂殖酵母、戴爾有孢圓酵母、拜耳接合酵母
3.2 降低蘋果酸和產乳酸能力
擁有較強的乳酸脫氫酶活性,可代謝丙酮酸生成乳酸(圖4),乳酸的酸感類似于檸檬水果,可增加葡萄酒口感的清新感。研究表明,某些菌株可產高達16 g·L-1乳酸,明顯降低葡萄酒的pH(可降0.5左右),因此,可用于氣候炎熱的葡萄酒產區如中國新疆產區增加葡萄酒的酸度[6,14-15,28]。與之相反,氣候冷涼區的葡萄酒往往因含有較高的蘋果酸而需要降低酸度,維持葡萄酒的生物穩定性[9]。裂殖酵母和擁有蘋果酸酒精代謝途徑(圖4),可將蘋果酸完全代謝形成乙醇和二氧化碳。一方面,具有特定的蘋果酸通透酶轉運體(mae1p)從而促進蘋果酸鹽進入細胞質;另一方面,的蘋果酸脫氫酶親和力是釀酒酵母的15倍,可在細胞質中代謝蘋果酸[28]。對蘋果酸的降解率普遍在75%—100%,可明顯提升葡萄酒的pH,若接種高濃度的則能完全降解蘋果酸至含量為0.096 mg·L-1[29]。、和也具有在混合發酵過程中降低蘋果酸的能力,如對蘋果酸的降解率可達91.6%,但降酸機理尚不明確[9,45,66-67,69]。
值得注意的是,接種方式對改善葡萄酒口感平衡的影響,非釀酒酵母屬酵母純種發酵和與釀酒酵母混合發酵的對比研究將促進對非釀酒酵母屬酵母改善葡萄酒口感平衡方面的認識。此外,對于高產甘油、產生多糖等尚缺乏標準化的數值范圍界定,一般對比于釀酒酵母純種發酵。因此,建議開展同一模擬汁條件下的發酵試驗,便于不同研究者比較非釀酒酵母屬酵母的口感平衡改善能力,提升高產或低產結論判斷的準確性。
4 可提升葡萄酒安全與健康的非釀酒酵母屬酵母
在葡萄酒釀造中應用非釀酒酵母屬酵母面臨的主要質疑是是否會產生影響葡萄酒質量或安全的物質。近年來,相關研究表明某些非釀酒酵母屬酵母具備生物防治作用,釀造過程中可降低有毒化合物的產量,產生生物活性物質,因此可提升葡萄酒安全質量與健康屬性。
4.1 生物防治作用
布魯塞爾酒香酵母()因大量產生揮發性酚類物質——4-乙基酚和4-乙基愈創木酚,使葡萄酒產生不好的動物類氣味而被認為是葡萄酒敗壞性酵母[70-71]。研究表明,釀酒酵母、、、和可分泌毒素抑制的生存[18,72-73],可通過分泌-1,3-葡聚糖酶、明膠酶和蛋白酶降解真菌細胞壁從而抑制的生長[74],因此,非釀酒酵母屬酵母可以通過生物防治作用提升葡萄酒的安全性。、和對灰霉菌有拮抗作用,還可拮抗鏈格孢屬真菌,還可拮抗曲霉和青霉,對曲霉具有拮抗作用[18],因此,非釀酒酵母屬酵母可用作抗真菌劑噴灑在葡萄上提高最終酒的質量[75]。非釀酒酵母屬酵母作為生物防治劑,一方面可以降低二氧化硫的用量,因為二氧化硫雖然可抑菌和抗氧化,但對人體具有毒性或引起過敏反應,因此國際組織一直在呼吁降低二氧化硫在葡萄酒中的用量,現代的消費者群體也更傾向于選擇使用較少防腐劑的食物[18,72]。另一方面,釀酒商在實踐中發現,紅葡萄酒發酵過程中使用非釀酒酵母屬酵母替代二氧化硫作為生物保護劑可增加果味,經檢測主要增加了乙酯類化合物的含量,這些酯類貢獻于紅色漿果和新鮮水果香氣[35]。目前,已經應用于葡萄酒釀造生物防治的商業非釀酒酵母屬酵母包括、、、和(表1)。
4.2 降低有毒化合物
葡萄酒釀造中應用非釀酒酵母屬酵母可減少可能出現的有毒化合物如赭曲霉毒素A、氨基甲酸乙酯(發酵型食品中的一種致癌物質)和生物胺[76]。①葡萄采收前敗壞性真菌侵染會產生赭曲霉毒素A,一方面,等前述生物防治劑可噴灑在葡萄表面抑制赭曲霉毒素真菌的生長,從而減少該毒素的產生[9];另一方面,通過細胞壁吸收作用能在發酵過程中去除70%的赭曲霉毒素A[77]。②氨基甲酸乙酯主要由乳酸菌產生,是尿素與乙醇在葡萄酒陳釀期間通過化學反應生成,葡萄酒工業中最常用的方法是使用商業脲酶去除尿素,從而避免氨基甲酸乙酯的生成。具有脲酶活性,因此應用于避免氨基甲酸乙酯的生成[9]。③葡萄酒中生物胺的含量為痕量至150 mg·L-1,部分來自于葡萄,部分是在酒精發酵、蘋乳發酵和陳釀過程中產生。有些非釀酒酵母屬酵母高產氨基酸,如在酒精發酵過程中會產生高濃度的賴氨酸、鳥氨酸和酪氨酸[9]。從原理上講,酵母菌可將精氨酸轉化為胍丁胺,鳥氨酸轉化為腐胺,甲硫氨酸轉化為亞精胺和精胺;賴氨酸為尸胺,組氨酸為組胺,色氨酸為色胺,酪氨酸為酪胺[77]。具有脫羧酶活性的蘋果酸乳酸菌能將某些氨基酸轉化為生物胺[9]。白葡萄酒中主要的生物胺是腐胺、尸胺和乙醇胺,紅葡萄酒中主要的生物胺是組胺、酪胺和腐胺,通常紅葡萄酒中生物胺的含量高于其他葡萄酒,而健康葡萄酒中生物胺的含量應小于50 mg·L-1[77]。人體中的單胺氧化酶可作用于生物胺起脫毒作用,但乙醇抑制該酶活性,因此,大量飲用葡萄酒時需關注生物胺的影響。目前避免葡萄酒中生物胺生成的主要方法是使用優選的不具有脫羧酶活性的蘋果酸乳酸菌。有些非釀酒酵母屬酵母可通過避免蘋果酸乳酸發酵的發生來控制生物胺的生成,如具有較強蘋果酸代謝能力的[9]。和結合使用,可以避免蘋乳發酵,產生更少生物胺和氨基甲酸乙酯的前體物質[77]。此外,H525具有胺氧化酶活性,可以降解生物胺[77]。
4.3 合成生物活性物質
葡萄酒因富含具有極強抗氧化性的多酚類物質而被認為有益于健康[22],最新的研究表明,酵母菌可以在酒精發酵過程中合成生物活性物質如褪黑素、血清素和羥酪醇,使用色氨酸N15標記和酪氨酸C13標記發現,酵母菌在酒精發酵過程中可以以色氨酸為前體物質合成褪黑素,以酪氨酸或Erlich途徑的中間產物形成羥酪醇[78]。非釀酒酵母屬酵母、、、和都可以在酒精發酵中產生胞內和胞外褪黑素,其中胞內褪黑素主要在酒精發酵前6 h內產生,因此,褪黑素可能有助于酵母菌適應新的發酵環境。非釀酒酵母屬酵母與釀酒酵母混合發酵沒有改變褪黑素的形成規律,和相對高產褪黑素[79]。
5 非釀酒酵母屬酵母的基因組和種內基因型分析方法研究現狀
新一代測序技術的發展促進了不同葡萄酒產區本土非釀酒酵母屬酵母的資源調查[80]和非釀酒酵母屬酵母的基因組及種內基因型分析方法的發展[81]。本研究列舉的可積極貢獻于葡萄酒發酵的非釀酒酵母屬酵母中只有1個菌種尚無基因組序列,9個菌種、、、、、、、和基因組測序菌株數量超過10個(表2),基于基因組信息開發的種內基因型分析方法主要為微衛星基因分型,其多態性和穩定性可以用于大量菌株和不同實驗室菌株之間的種群遺傳分析,為葡萄酒發酵過程中特定基因型非釀酒酵母屬酵母的追蹤監控提供可靠的分子依據。然而,目前微衛星位點分析只集中于5個非釀酒酵母屬酵母菌種、和(表2),今后應加大對各產區廣泛存在的本土非釀酒酵母屬酵母和具有葡萄酒發酵應用價值的非釀酒酵母屬酵母的微衛星位點分析。此外,關于大部分非釀酒酵母屬酵母有性生殖的數據比較少(表2),至今仍然不能成功進行非釀酒酵母屬酵母的經典遺傳操作[81-82],因此,對于非釀酒酵母屬酵母相應基因功能的認識往往通過與釀酒酵母功能基因的比對和代謝物質分析來完成。有研究者指出可通過原生質體融合實現種內或種間雜交,最終確定與葡萄酒發酵應用特點相關的基因,為葡萄酒質量的準確控制提供理論基礎。
6 結論與展望
非釀酒酵母屬酵母曾經一度被認為是導致葡萄酒敗壞的罪魁禍首,但實際上無論是否接種釀酒酵母,酒精發酵前期的葡萄醪中都存在大量非釀酒酵母屬酵母如等。因此,近年來葡萄酒發酵研究熱點轉向控制性地應用非釀酒酵母屬酵母,主要采用與釀酒酵母同時接種混合發酵和順序接種混合發酵兩種方式,將非釀酒酵母屬酵母在顏色、香氣、口感和安全健康等方面的優勢與釀酒酵母強發酵和強耐受的特點結合起來,釀造具有產區特色的高質量葡萄酒。
為更好地應用非釀酒酵母屬酵母,未來研究者們仍需關注以下6個方面:
第一,關注中國本土野生酵母菌的發掘與應用,促進本土非釀酒酵母屬酵母商業化。近年來,中國葡萄酒在國際賽事上頻頻獲獎,分離自新疆、寧夏產區的釀酒酵母CECA、CEC01和分離自東北產區的CVE-7,已經成功商業化并走出國門服務于世界葡萄酒產業(表1)。而中國不同葡萄酒產區的非釀酒酵母屬酵母多樣性分析研究,展現了各個產區不同風土條件下豐富的非釀酒酵母屬酵母資源[37,58,80,88],分離自新疆的優選和分離自遼寧的優選及已被安琪公司加工成活性干酵母用于研究[56]。進一步開發具有中國產區特色的優選非釀酒酵母屬酵母,對保護中國本土酵母資源,釀造具有中國不同產區特色的葡萄酒具有重要意義。
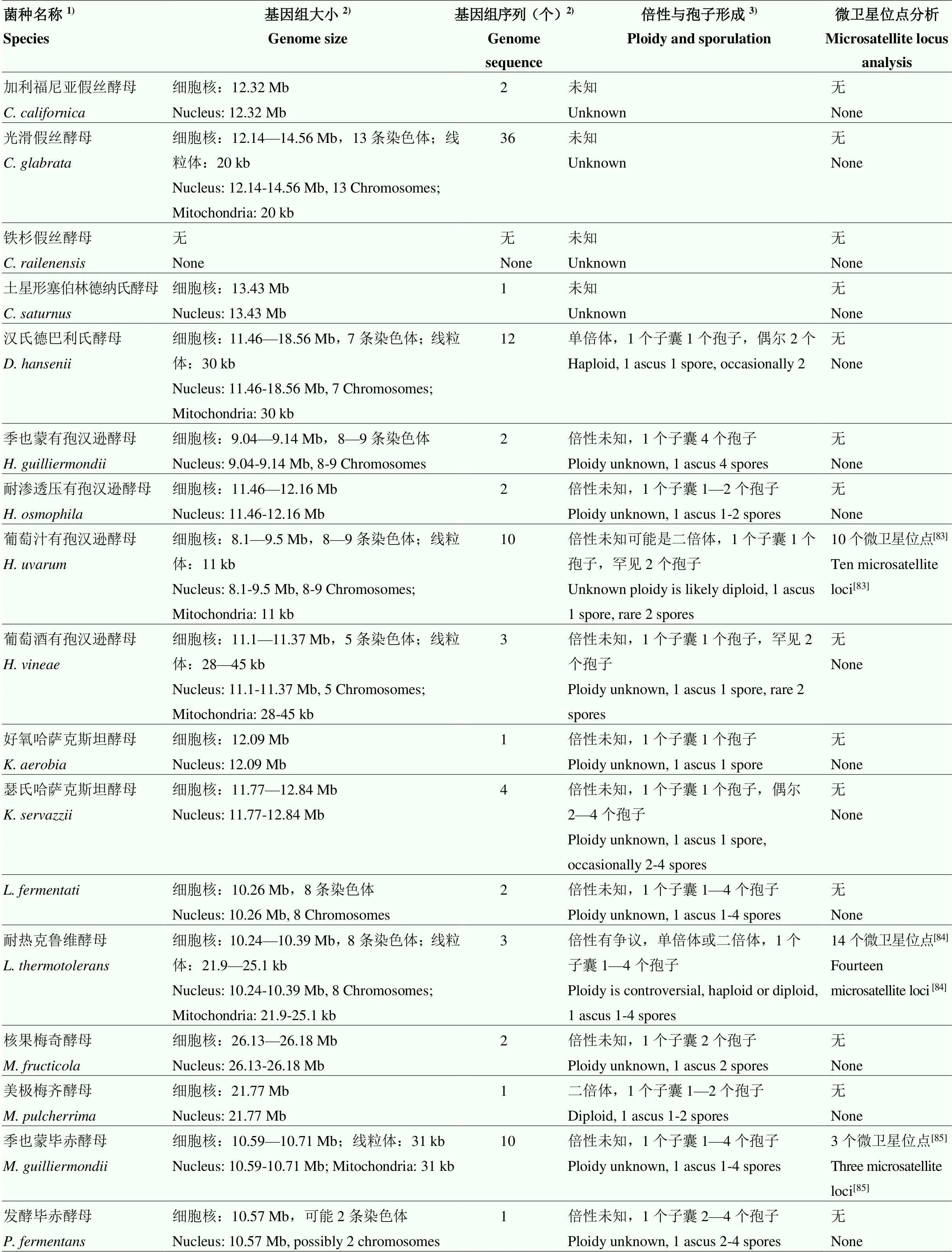
表2 非釀酒酵母屬酵母的基因組相關信息
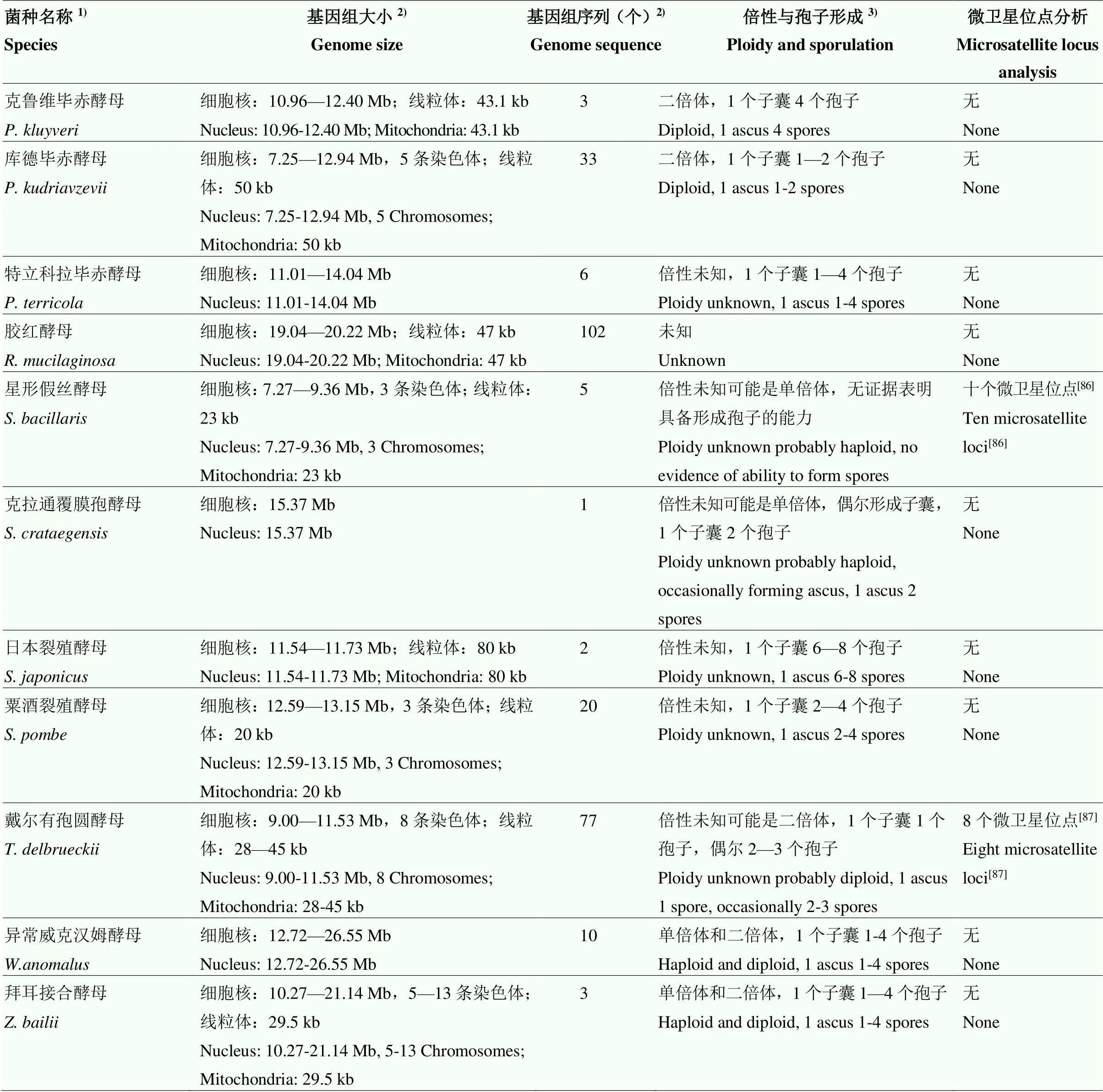
續表2 Continued table 2
1)只包含本研究列舉的具有葡萄酒發酵應用特點的非釀酒酵母屬酵母;2)基因組大小和序列信息獲取自NCBI網址(http://www.ncbi.nlm.nih.gov/ genome/)2022年6月22日檢索結果和參考文獻[81];3)倍性和孢子形成信息參考文獻[2,81]
1)This only included the non-yeast which had the trait of wine fermentation applications;2)Genome size and sequence information obtained from NCBI web site (http://www.ncbi.nlm.nih.gov/genome/) search results and references on 22 June, 2022[81];3)References for information on ploidy and spore formation[2,81]
第二,關注非釀酒酵母屬酵母基因組和種內基因型分析。如前所述,野生酵母菌可能在葡萄酒發酵過程中存在并發揮作用,因此,分析主導酵母菌的基因型將有利于控制和認識葡萄酒質量的改變,尤其是對應用多種非釀酒酵母屬酵母的葡萄酒發酵來說,基因型分析可確認發酵過程的主導菌。
第三,關注非釀酒酵母屬酵母的純種發酵和混合發酵代謝研究,增加對非釀酒酵母屬酵母貢獻的認識。非釀酒酵母屬酵母早期的研究以純種發酵為主,但其大多無法主導酒精發酵全過程,近年來,大多數研究直接以與釀酒酵母混合發酵某個葡萄品種的方式并對比釀酒酵母純種發酵來認識非釀酒酵母屬酵母的貢獻,為在葡萄酒工業中直接應用非釀酒酵母屬酵母提供了研究基礎。但此方法從理論上無法辨別葡萄酒質量的提升是否真正來自于非釀酒酵母屬酵母本身的代謝優勢。一方面,研究表明混合發酵過程中釀酒酵母與非釀酒酵母屬酵母之間的相互作用改變了二者在純種發酵下的代謝模式,如非釀酒酵母屬酵母激發釀酒酵母糖代謝相關基因的表達[89],具有嗜殺性的釀酒酵母在混合發酵中產生了更高濃度的果味酯類物質[90],非釀酒酵母屬酵母和釀酒酵母之間對營養的競爭極大促進了香氣化合物的產生[91],具有相互抑制作用的(低產甘油)和釀酒酵母(高產甘油)在混合發酵中增加了甘油產量[92]。代謝組學分析表明,共同接種導致了新化合物的生成,酵母之間的相互作用影響了戊糖磷酸和氨基酸合成途徑[93],因此,以釀酒酵母與非釀酒酵母屬酵母純種發酵和不同比例的接種發酵作為對比,將有利于正確認識非釀酒酵母屬酵母的釀造應用特點。另一方面,葡萄營養成分(碳源、氮源等)與發酵工藝(接種順序、通氧等)的不同可能會影響酵母之間的相互作用,進而影響葡萄酒的風味[94]。如銨能促進和對氨基酸的吸收,而不影響釀酒酵母對氨基酸的吸收[12],因此,順序接種混合發酵中非釀酒酵母屬酵母對氮源的利用是否會導致釀酒酵母的代謝調節值得思考。非釀酒酵母屬酵母的嗜果糖特性[95],非釀酒酵母屬酵母的脂類代謝與酒精發酵耐受性的關聯[96],發酵初期通風對酵母菌乳酸代謝的促進[41],順序接種發酵對葡萄酒品質的提升優于同時接種發酵[47]等,說明原料和工藝都會影響非釀酒酵母屬酵母在混合發酵中的表現,因此,篩選和應用非釀酒酵母屬酵母時需綜合考量這些因素的影響。
第四,關注最終成品葡萄酒質量與非釀酒酵母屬酵母代謝特點之間的關聯。過去一直認為酒精發酵過程中高產乙醛和乙酸乙酯的酵母會給葡萄酒帶來不良風味,但最近有研究發現24個月的陳釀會降低葡萄酒中乙醛和乙酸乙酯的含量[30],因此,鼓勵研究者從最終產品考量具有極端突出代謝特點的非釀酒酵母屬酵母菌株。此外,葡萄酒的顏色、香氣和口感隨著后續工藝和時間不斷發生變化,因此,非釀酒酵母屬酵母的代謝對最終成品的貢獻值得探究。
第五,關注非釀酒酵母屬酵母對葡萄與葡萄酒安全與健康的影響。非釀酒酵母屬酵母是否可能取代二氧化硫,是否可以有效去除葡萄酒中的毒性成分,是否可能促進有益成分如褪黑素等的分泌需要更多的研究證實。
第六,關注非釀酒酵母屬酵母商業菌劑的加工類型(活性干酵母、黏稠奶油狀的液體酵母、活性冷凍酵母、膠囊封裝酵母等)對非釀酒酵母屬酵母活性的影響[16]。目前,釀酒酵母主要加工成活性干酵母應用于葡萄酒工業,非釀酒酵母屬酵母對極端環境的耐受性弱于釀酒酵母,因此,需要關注加工過程對非釀酒酵母屬酵母活性的影響。
[1] JOLLY N P, VARELA C, PRETORIUS I S. Not your ordinary yeast: Non-yeasts in wine production uncovered. FEMS Yeast Research, 2014, 14(2): 215-237. doi: 10.1111/1567-1364.12111.
[2] KURTZMAN C P, FELL J W, BOEKHOUT T. The Yeasts: A Taxonomic Study. 5th ed. Amsterdam: Elsevier, 2011.
[3] DRUMONDE-NEVES J, FERNANDES T, LIMA T, PAIS C, FRANCO-DUARTE R. Learning from 80 years of studies: A comprehensive catalogue of non-yeasts associated with viticulture and winemaking. FEMS Yeast Research, 2021, 21(3): foab017. doi: 10.1093/femsyr/foab017.
[4] PADILLA B, GIL J V, MANZANARES P. Past and future of non-yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Frontiers in Microbiology, 2016, 7: 411.
[5] MAS A, PADILLA B, ESTEVE-ZARZOSO B, BELTRAN G, REGUANT C, BORDONS A. Taking advantage of natural biodiversity for wine making: The wildwine project.Agriculture and Agricultural Science Procedia, 2016, 8: 4-9.
[6] BERBEGAL C, FRAGASSO M, RUSSO P, BIMBO F, GRIECO F, SPANO G, CAPOZZI V. Climate changes and food quality: The potential of microbial activities as mitigating strategies in the wine sector. Fermentation, 2019, 5: 85.
[7] LAIRóN-PERIS M, PéREZ-TRAVéS L, MU?IZ-CALVO S, GUILLAMóN J M, HERAS J M, BARRIO E, QUEROL A. Differential contribution of the parental genomes to a×hybrid, inferred by phenomic, genomic, and transcriptomic analyses, at different industrial stress conditions. Frontiers Media, 2020, 8: 129.
[8] ALBERTIN W, ZIMMER A, MIOT-SERTIER C, BERNARD M, COULON J, MOINE V, COLONNA-CECCALDI B, BELY M, MARULLO P, MASNEUF-POMAREDE I. Combined effect of thelag phase and the non-consortium to enhance wine fruitiness and complexity. Applied Microbiology and Biotechnology, 2017, 101(20): 7603-7620.
[9] BENITO S, CALDERóN F. The influence of non-species on wine fermentation quality parameters. Fermentation, 2019, 5: 54.
[10] BORREN E, TIAN B. The important contribution of non-yeasts to the aroma complexity of wine: A review. Foods, 2020, 10: 13.
[11] TAILLANDIER P, LAI Q P, JULIEN-ORTIZ A, BRANDAM C. Interactions betweenandin wine fermentation: Influence of inoculation and nitrogen content. World Journal of Microbiology and Biotechnology, 2014, 30(7): 1959-1967.
[12] PRIOR K J, BAUER F F, DIVOL B. The utilisation of nitrogenous compounds by commercial non-yeasts associated with wine. Food Microbiology, 2019, 79: 75-84.
[13] ROUDIL L, RUSSO P, BERBEGAL C, ALBERTIN W, SPANO G, CAPOZZI V. Non-commercial starter cultures: scientific trends, recent patents and innovation in the wine sector. Recent Patents on Food, Nutrition & Agriculture, 2020, 11(1): 27-39.
[14] 唐沖, 成池芳, 許引虎, 段長青, 燕國梁. 耐熱克魯維酵母在葡萄酒發酵中的研究進展. (2022-04-15) [2022-06-25].食品科學:1-12. DOI:10.7506/spkx1002-6630-20211223-266.
TANG C, CHENG C F, XU Y H, DUAN C Q, YAN G L. Research progresses ofin wine fermentation. (2022-04-15) [2022-06-25].Food Science:1-12. doi:10.7506/spkx1002- 6630-20211223-266. (in Chinese)
[15] 董琦楠, 李瑩, 葉冬青, 劉延琳. 耐熱克魯維酵母在葡萄酒釀造中的研究進展. 微生物學通報, 2022, 49(5): 1941-1954. doi: 10.13344/j.microbiol.china.210653.
DONG Q N, LI Y, YE D Q, LIU Y L. Research progress ofin winemaking. Microbiology China, 2022, 49(5): 1941-1954. doi: 10.13344/j.microbiol.china.210653. (in Chinese)
[16] VEJARANO R, GIL-CALDERóN A. Commercially available non-yeasts for winemaking: Current market, advantages over, biocompatibility, and safety. Fermentation, 2021, 7(3): 171.
[17] LIN M M-H, BOSS P K, WALKER M E, SUMBY K M, JIRANEK V. Influence ofspp. on the chemical and sensory profile of red wines.International Journal of Food Microbiology, 2022, 362: 109496.
[18] GIANVITO P D, ENGLEZOS V, RANTSIOU K, COCOLIN L. Bioprotection strategies in winemaking. International Journal of Food Microbiology, 2022, 364: 109532.
[19] TUFARIELLO M, FRAGASSO M, PICO CARBAJO J, PANIGHEL A, CASTELLARIN S D, FLAMINI R, GRIECO F. Influence of non-on wine chemistry: A focus on aroma-related compounds. Molecules, 2021, 26: 644.
[20] CLEMENTE-JIMENEZ J M, MINGORANCE-CAZORLA L,MARTI?NEZ-RODRI?GUEZ S, HERAS-VáZQUEZ F J L, RODRI?GUEZ- VICO F. Molecular characterization and oenological properties of wine yeasts isolated during spontaneous fermentation of six varieties of grape must. Food Microbiology, 2004, 21(2): 149-155.
[21] WANG C X, ESTEVE-ZARZOSO B, MAS A. Monitoring of,, and(synonym) populations during alcoholic fermentation by fluorescence in situ hybridization. International Journal of Food Microbiology, 2014, 191: 1-9.
[22] 李華, 王華, 袁春龍. 葡萄酒工藝學. 北京: 科學出版社, 2007: 24-27.
LI H, WANG H, YUAN C L. Wine Technology. Beijing: Science Press, 2007: 24-27. (in Chinese)
[23] 王二雷, 劉彥君, 劉靜波. 吡喃型花青素類化合物的研究進展. 食品工業科技, 2018, 39(12): 325-333. doi: 10.13386/j.issn1002-0306. 2018.12.059.
WANG E L, LIU Y J, LIU J B. Research advances in pyranoanthocyanin compounds. Science and Technology of Food Industry, 2018, 39(12): 325-333. doi: 10.13386/j.issn1002-0306.2018. 12.059. (in Chinese)
[24] LI X S, TENG Z J, LUO Z Y, YUAN Y B, ZENG Y Y, HU J, SUN J X, BAI W B. Pyruvic acid stress caused color attenuation by interfering with anthocyanins metabolism during alcoholic fermentation. Food Chemistry, 2022, 372:131251.
[25] 張寧, 趙旭, 蘭義賓, 石英, 段長青, 吳廣楓. 中國東亞種紅葡萄酒的顏色特征及酚類組成研究. 中國釀造, 2022, 41(5): 34-41. doi: 10.11882/j.issn.0254-5071.2022.05.007.
ZHANG N, ZHAO X, LAN Y B, SHI Y, DUAN C Q, WU G F. Color features and phenolic composition of red wines made from East Asianspecies native to China. China Brewing, 2022, 41(5): 34-41. doi: 10.11882/j.issn.0254-5071.2022.05.007. (in Chinese)
[26] BENITO S, PALOMERO F, MORATA A, CALDERóN F, PALMERO D, SUáREZ-LEPE J A. Physiological features ofof interest in making of white wines. European Food Research and Technology, 2013, 236(1): 29-36.
[27] BENITO á, CALDERóN F, BENITO S. The combined use ofand- Effect on the anthocyanin wine composition. Molecules, 2017, 22(5): 739.
[28] MORATA A, ESCOTT C, BA?UELOS M A, LOIRA I, DEL FRESNO J M, GONZáLEZ C, SUáREZ-LEPE J A. Contribution of non-yeasts to wine freshness. A review. Biomolecules, 2020, 10(1): 34.
[29] 趙美, 田秀, 李敏, 高娉娉, 梁麗紅, 韓舜愈, 王婧. 粟酒裂殖酵母與釀酒酵母共同接種發酵對‘黑比諾’干紅葡萄酒品質的影響. 食品科學, 2021, 42(24): 108-116. doi: 10.7506/spkx1002-6630- 20201012-100.
ZHAO M, TIAN X, LI M, GAO P P, LIANG L H, HAN S Y, WANG J. Effect of mixed culture fermentation withandon the quality of ‘Pinot Noir’ dry red wine. Food Science, 2021, 42(24): 108-116. doi: 10.7506/ spkx1002-6630-20201012-100. (in Chinese)
[30] PORTARO L, MAIOLI F, CANUTI V, PICCHI M, LENCIONI L, MANNAZZU I, DOMIZIO P./mixed starter cultures: New perspectives for the improvement ofaroma, taste, and color stability. LWT, 2022, 156: 113009.
[31] VELENOSI M, CRUPI P, PERNIOLA R, MARSICO A D, SALERNO A, ALEXANDRE H, ARCHIDIACONO N, VENTURA M, CARDONE M F. Color stabilization of Apulian red wines through the sequential inoculation ofand. Molecules, 2021, 26(4): 907.
[32] 胡苑. 干紅葡萄酒陳釀衍生色素顏色穩定性的研究[D]. 煙臺: 煙臺大學, 2021.
HU Y. Study on the color stability of red wine derived pigments during aging [D]. Yantai: Yantai University, 2021. (in Chinese)
[33] HRANILOVIC A, GAMBETTA J M, SCHMIDTKE L, BOSS P K, GRBIN P R, MASNEUF-POMAREDE I, BELY M, ALBERTIN W, JIRANEK V. Oenological traits ofshow signs of domestication and allopatric differentiation. Scientific Reports, 2018, 8(1): 14812.
[34] MORATA A, ESCOTT C, LOIRA I, DEL FRESNO J M, GONZáLEZ C, SUáREZ-LEPE J A. Influence ofand non-yeasts in the formation of pyranoanthocyanins and polymeric pigments during red wine making. Molecules, 2019, 24(24): 4490.
[35] WINDHOLTZ S, REDON P, LACAMPAGNE S, FARRIS L, LYTRA G, CAMELEYRE M, BARBE J-C, COULON J, THIBON C, MASNEUF-POMARèDE I. Non-yeasts as bioprotection in the composition of red wine and in the reduction of sulfur dioxide. LWT, 2021, 149: 111781.
[36] 李婧. 冰葡萄酒發酵過程中酵母菌群落演潛規律及優良菌株的篩選[D]. 大連: 大連理工大學, 2020.
LI J. The dynamic changes of yeast populations during icewine fermentation and screening of the yeast strains [D]. Dalian: Dalian University of Technology, 2020. (in Chinese)
[37] HAN X Y, QING X, YANG S Y, LI R L, ZHAN J C, YOU Y L, HUANG W D. Study on the diversity of non-yeasts in Chinese wine regions and their potential in improving wine aroma by-glucosidase activity analyses.Food Chemistry, 2021, 360: 129886.
[38] GE Q, GUO C F, YAN Y, SUN X Y, MA T T, ZHANG J, LI C H, GOU C L, YUE T L, YUAN Y H. Contribution of non-yeasts to aroma-active compound production, phenolic composition and sensory profile in Chinese Vidal icewine. Food Bioscience, 2021, 46: 101152.
[39] BINATI R L, LEMOS JUNIOR W J F, LUZZINI G, SLAGHENAUFI D, UGLIANO M, TORRIANI S. Contribution of non-yeasts to wine volatile and sensory diversity: A study on,spp. andstrains isolated in Italy. International Journal of Food Microbiology, 2020, 318: 108470.
[40] SCANSANI S, VAN W N, NADER K B, BEISERT B, BREZINA S, FRITSCH S, SEMMLER H, PASCH L, PRETORIUS I S, VON WALLBRUNN C, SCHNELL S, RAUHUT D. The film-formingspp. in a winemaker’s toolbox: A simple isolation procedure and their performance in a mixed-culture fermentation ofL. cv. Gewürztraminer must.International Journal of Food Microbiology, 2022, 365: 109549.
[41] Yan G l, Zhang B q, Joseph L, Waterhouse A L. Effects of initial oxygenation on chemical and aromatic composition of wine in mixed starters ofand. Food Microbiology, 2020, 90:103460.
[42] HONG M N, LI J, CHEN Y w, QI B y, HUANG Y p, WU J, YUE H B, TONG Z Q, LIU Y N, WANG F. Impact of mixed non-yeast during fermentation on volatile aroma compounds of Vidal blanc icewine. LWT, 2021, 145: 111342.
[43] 孫玉霞, 趙新節. 美極梅奇酵母的代謝特性及其在葡萄酒生產中的應用前景. 食品與發酵工業, 2021, 47(4): 305-311. doi: 10.13995/j.cnki.11-1802/ts.025251.
SUN Y X, ZHAO X J. Metabolic characteristics ofand its application in wine production. Food and Fermentation Industries, 2021, 47(4): 305-311. doi: 10.13995/j.cnki. 11-1802/ts.025251. (in Chinese)
[44] LAI Y T, HSIEH C W, LO Y C, LIOU B K, LIN H W, HOU C Y, CHENG K C. Isolation and identification of aroma-producing non-yeast strains and the enological characteristic comparison in wine making. LWT, 2022, 154: 112653.
[45] BELDA I, RUIZ J, BEISERT B, NAVASCUéS E, MARQUINA D, CALDERóN F, RAUHUT D, BENITO S, SANTOS A. Influence ofin varietal thiol (3-SH and 4-MSP) release in wine sequential fermentations. International Journal of FoodMicrobiology, 2017, 257: 183-191.
[46] SEGUINOT P, BLOEM A, BRIAL P, MEUDEC E, ORTIZ-JULIEN A, CAMARASA C. Analysing the impact of the nature of the nitrogen source on the formation of volatile compounds to unravel the aroma metabolism of two non-strains. International Journal of Food Microbiology, 2020, 316: 108441.
[47] SHI W K, WANG J, CHEN F S, ZHANG X Y. Effect ofandon wine flavor and quality through simultaneous and sequential co-fermentation with. LWT, 2019, 116: 108477.
[48] 李愛華, 王星晨, 彭文婷, 李朔, 陶永勝. 膠紅酵母與釀酒酵母混合酒精發酵中酵母生長與糖苷酶活動力學. 西北農業學報, 2018, 27(6): 896-903. doi: 10.7606/j.issn.1004-1389.2018.06.018.
LI A H, WANG X C, PENG W T, LI S, TAO Y S. Kinetics of biomass and glycosidase activities during mixed alcoholic fermentation of selectedand. Acta Agriculturae Boreali-Occidentalis Sinica, 2018, 27(6): 896-903. doi: 10.7606/j.issn.1004-1389.2018.06.018. (in Chinese)
[49] WANG X C, LI A H, DIZY M, ULLAH N, SUN W X, TAO Y S. Evaluation of aroma enhancement for ‘Ecolly’ dry white wines by mixed inoculation of selectedand. Food Chemistry, 2017, 228: 550-559.
[50] 馬娜, 王星晨, 孔彩琳, 陶永勝. 膠紅酵母與釀酒酵母共發酵對干紅葡萄酒香氣與色澤的影響. 食品科學, 2021, 42(2): 97-104. doi: 10.7506/spkx1002-6630-20191112-158.
MA N, WANG X C, KONG C L, TAO Y S. Effect of mixed culture fermentation withandon the aroma and color of red wine. Food Science, 2021, 42(2): 97-104. doi: 10.7506/spkx1002-6630-20191112-158. (in Chinese)
[51] NISIOTOU A, SGOUROS G, MALLOUCHOS A, NISIOTIS C-S, MICHAELIDIS C, TASSOU C, BANILAS G. The use of indigenousandstrains as a tool to create chemical complexity in local wines.Food Research International, 2018, 111: 498-508.
[52] ENGLEZOS V, RANTSIOU K, CRAVERO F, TORCHIO F, POLLON M, FRACASSETTI D, ORTIZ-JULIEN A, GERBI V, ROLLE L, COCOLIN L. Volatile profile of white wines fermented with sequential inoculation ofand. Food Chemistry, 2018, 257: 350-360.
[53] LEE P R, SAPUTRA A, YU B, CURRAN P, LIU S Q. Effects of pure and mixed-cultures ofandon the volatile profiles of grape wine. Food Biotechnology, 2012, 26(4): 307-325. doi: 10.1080/08905436.2012.723606.
[54] GARAVAGLIA J, SCHNEIDER R DE C DE S, CAMARGO MENDES S D, WELKE J E, ZINI C A, CARAM?O E B, VALENTE P. Evaluation ofBCV 08 as a co-starter in wine fermentation for the improvement of ethyl esters production. Microbiological Research, 2015, 173: 59-65.
[55] MARTíNEZ-AVILA O, SáNCHEZ A, FONT X, BARRENA R. Bioprocesses for 2-phenylethanol and 2-phenylethyl acetate production:current state and perspectives.Applied Microbiology and Biotechnology, 2018, 102(23): 9991-10004.
[56] ZHANG B Q, TANG C, YANG D Q, LIU H, XUE J, DUAN C Q, YAN G L. Effects of three indigenous non-yeasts and their pairwise combinations in co-fermentation withon volatile compounds of Petit Manseng wines.Food Chemistry, 2022, 368: 130807.
[57] 張清安, 陳博宇. 葡萄酒中與風味相關4類含硫化合物的研究進展. 中國農業科學, 2020, 53(5): 1029-1045. doi: 10.3864/j.issn.0578- 1752.2020.05.014.
ZHANG Q A, CHEN B Y. Research progress of four sulfur compounds related to red wine flavor. Scientia Agricultura Sinica, 2020, 53(5): 1029-1045. doi: 10.3864/j.issn.0578-1752.2020.05.014. (in Chinese)
[58] ZHANG P Z, ZHANG R G, SIRISENA S, GAN R Y, FANG Z X. Beta-glucosidase activity of wine yeasts and its impacts on wine volatiles and phenolics: A mini-review.Food Microbiology, 2021, 100: 103859.
[59] 任學梅, 姚紅紅, 嚴幻汝, 祝霞, 楊學山. 高產糖苷酶非釀酒酵母菌株篩選、鑒定及其發酵過程中酶活性變化. 食品科學, 2022, 43(20): 198-206.
REN X M, YAO H H, YAN H R, ZHU X, YANG X S. Screening and identification of non-yeast strains with high glycosidase production and changes in enzyme activities during their fermentation. Food Science, 2022, 43(20): 198-206. (in Chinese)
[60] 馬延琴, 徐曉裕, 李甜, 李春燕, 蔣霞, 王斌, 史學偉. 釀酒葡萄表皮產酶非釀酒酵母的篩選及其產酶特性研究. 中國釀造, 2021, 40(12): 149-154. doi: 10.11882/j.issn.0254-5071.2021.12.027.
MA Y Q, XU X Y, LI T, LI C Y, JIANG X, WANG B, SHI X W. Screening of non-from wine grape epidermis and its enzyme-producing ability. China Brewing, 2021, 40(12): 149-154. doi: 10.11882/j.issn.0254-5071.2021.12.027. (in Chinese)
[61] ROMANO P, SUZZI G. Higher alcohol and acetoin production bywine yeasts. Journal of Applied Bacteriology, 1993, 75(6): 541-545.
[62] PUERTAS B, JIMENEZ-HIERRO M J, CANTOS-VILLAR E, MARRUFO-CURTIDO A, CARBú M, CUEVAS F J, MORENO- ROJAS J M, GONZáLEZ-RODRíGUEZ V E, CANTORAL J M, RUIZ-MORENO M J. The influence of yeast on chemical composition and sensory properties of dry white wines. Food Chemistry, 2018, 253: 227-235.
[63] Zhu L X, Wang G, Aihaiti A. Combined indigenous yeast strains produced local wine from over ripen Cabernet Sauvignon grape in Xinjiang. World Journal of Microbiology and Biotechnology, 2020, 36(8): 122.
[64] DOMIZIO P, LIU Y, BISSON L F, BARILE D. Use of non-wine yeasts as novel sources of mannoproteins in wine. Food Microbiology, 2014, 43: 5-15.
[65] DOMIZIO P, LENCIONI L, CALAMAI L, PORTARO L, BISSON L F. Evaluation of the yeastfor use in wine production. American Journal of Enology and Viticulture, 2018, 69(3): 266-277.
[66] KIM D H, HONG Y A, PARK H D. Co-fermentation of grape must byandreduces the malic acid content in wine. Biotechnology Letters, 2008, 30(9): 1633-1638.
[67] HONG S K, LEE H J, PARK H J, HONG Y A, RHEE I K, LEE W H, CHOI S W, LEE O S, PARK H D. Degradation of malic acid in wine by immobilizedcells with oriental oak charcoal and alginate. Letters in Applied Microbiology, 2010, 50(5): 522-529. doi: 10.1111/j.1472-765X.2010.02833.x.
[68] BELDA I, NAVASCUéS E, MARQUINA D, SANTOS A, CALDERON F, BENITO S. Dynamic analysis of physiological properties ofin wine fermentations and its incidence on wine quality. Applied Microbiology and Biotechnology, 2015, 99(4): 1911-1922.
[69] 白玉峰, 張文霞, 田亞楠, 張秀艷. 寧夏賀蘭山東麓降L-蘋果酸葡萄酒酵母的篩選. 中國釀造, 2021, 40(1): 49-54. doi: 10.11882/ j.issn.0254-5071.2021.01.010.
BAI Y F, ZHANG W X, TIAN Y N, ZHANG X Y. Screening of yeast strains with L-malic acid degradation ability from Eastern Foothills of Helan in Ningxia. China Brewing, 2021, 40(1): 49-54. doi: 10.11882/j.issn.0254-5071.2021.01.010. (in Chinese)
[70] MILHEIRO J, FILIPE-RIBEIRO L, VILELA A, COSME F, NUNES F M. 4-Ethylphenol, 4-ethylguaiacol and 4-ethylcatechol in red wines: Microbial formation, prevention, remediation and overview of analytical approaches. Critical Reviews in Food Science and Nutrition, 2019, 59(9): 1367-1391. doi: 10.1080/10408398.2017.1408563.
[71] FLEET G H. Wine yeasts for the future. FEMS Yeast Research, 2008, 8(7): 979-995. doi: 10.1111/j.1567-1364.2008.00427.x.
[72] KUCHEN B, MATURANO Y P, MESTRE M V, COMBINA M, TORO M E, VAZQUEZ F. Selection of native non-yeasts with biocontrol activity against spoilage yeasts in order to produce healthy regional wines. Fermentation, 2019, 5: 60.
[73] COMITINI F, AGARBATI A, CANONICO L, GALLI E, CIANI M. Purification and characterization of WA18, a new mycocin produced byactive in wine againstspoilage yeasts. Microorganisms, 2020, 9: 56.
[74] YAN W, GAO H, QIAN X J, JIANG Y J, ZHOU J, DONG W L, XIN F X, ZHANG W M, JIANG M. Biotechnological applications of the non-conventional yeast.Biotechnology Advances, 2021, 46: 107674.
[75] BENITO S. The impacts ofyeast strains on winemaking. Applied Microbiology and Biotechnology, 2018, 102(16): 6775-6790.
[76] BENITO S. The impacts ofon winemaking. Applied Microbiology and Biotechnology, 2019, 103(11): 4291-4312.
[77] WANG B S, TAN F L, CHU R C, LI G Y, LI L B, YANG T Y, ZHANG M X. The effect of non-yeasts on biogenic amines in wine. Trends in Food Science & Technology, 2021, 116: 1029-1040.
[78] GALLARDO-FERNáNDEZ M, VALLS-FONAYET J, VALERO E, HORNEDO-ORTEGA R, RICHARD T, TRONCOSO A M, GARCIA-PARRILLA M C. Isotopic labelling-based analysis elucidates biosynthesis pathways infor melatonin, serotonin and hydroxytyrosol formation. Food Chemistry, 2022, 374: 131742.
[79] MORCILLO-PARRA M á, GONZáLEZ B, BELTRAN G, MAS A, TORIJA M J. Melatonin and glycolytic protein interactions are related to yeast fermentative capacity.Food Microbiology, 2020, 87: 103398.
[80] WANG C X, WU C, QIU S Y. Yeast diversity investigation ofF?ex during spontaneous fermentations using culture- dependent and high-throughput sequencing approaches. Food Research International, 2019, 126: 108582.
[81] MASNEUF-POMAREDE I, BELY M, MARULLO P, ALBERTIN W. The genetics of non-conventional wine yeasts: Current knowledge and future challenges. Frontiers in Microbiology, 2016, 6: 1563.
[82] 梁樹英, 王春曉. 基于倍性的葡萄酒相關酵母釀造特性研究進展. 食品與發酵科技, 2022, 58(1): 121-130. doi: 10.3969/j.issn.1674- 506X.2022.01-017.
LIANG S Y, WANG C X. Research progress in wine-making characteristic of wine-related yeast based on ploidy. Sichuan Food and Fermentation, 2022, 58(1): 121-130. doi: 10.3969/j.issn.1674-506X. 2022.01-017. (in Chinese)
[83] ALBERTIN W, SETATI M E, MIOT-SERTIER C, MOSTERT T T, COLONNA-CECCALDI B, COULON J, GIRARD P, MOINE V, PILLET M, SALIN F, BELY M, DIVOL B, MASNEUF- POMAREDE I.from winemaking environments show spatial and temporal genetic clustering.Frontiers in Microbiology, 2016, 6: 1569.
[84] HRANILOVIC A, BELY M, MASNEUF-POMAREDE I, JIRANEK V, ALBERTIN W, FAIRHEAD C. The evolution ofis driven by geographical determination, anthropisation and flux between different ecosystems. PLoS ONE, 2017, 12(9): e0184652.
[85] WRENT P, RIVAS E M, PEINADO J M, DE SILóNIZ M I. Development of an affordable typing method forusing microsatellite markers. International Journal of Food Microbiology, 2016, 217: 1-6.
[86] MASNEUF-POMAREDE I, JUQUIN E, MIOT-SERTIER C, RENAULT P, LAIZET Y, SALIN F, ALEXANDRE H, CAPOZZI V, COCOLIN L, COLONNA-CECCALDI B, ENGLEZOS V, GIRARD P, GONZALEZ B, LUCAS P, MAS A, NISIOTOU A, SIPICZKI M, SPANO G, TASSOU C, BELY M, ALBERTIN W. The yeast(synonym) shows high genetic diversity in winemaking environments. FEMS Yeast Research, 2015, 15(5): fov045. doi: 10.1093/femsyr/fov045.
[87] ALBERTIN W, CHASSERIAUD L, COMTE G, PANFILI A, DELCAMP A, SALIN F, MARULLO P, BELY M, SCHACHERER J. Winemaking and bioprocesses strongly shaped the genetic diversity of the ubiquitous yeast. PLoS ONE, 2014, 9(4): e94246.
[88] 田進, 吳成, 楊金仙, 王春曉. 葡萄酒有孢漢遜酵母屬分子指紋圖譜分析. 菌物學報, 2020, 39(4): 755-765. doi: 10.13346/j. mycosystema.190404.
TIAN J, WU C, YANG J X, WANG C X. The molecular fingerprinting analysis ofin wine. Mycosystema, 2020, 39(4): 755-765. doi: 10.13346/j.mycosystema.190404. (in Chinese)
[89] MENCHER A, MORALES P, CURIEL J A, GONZALEZ R, TRONCHONI J.represses aerobic respiration insuggesting a direct response to co-cultivation. Food Microbiology, 2021, 94: 103670.
[90] LI Y Q, HU K, XU Y H, MEI W C, TAO Y S. Biomass suppression ofby killerhighly increased fruity esters in mixed culture fermentation. LWT, 2020, 132: 109839.
[91] ROLLERO S, BLOEM A, BRAND J, ORTIZ-JULIEN A, CAMARASA C, DIVOL B. Nitrogen metabolism in three non-conventional wine yeast species: A tool to modulate wine aroma profiles. Food Microbiology, 2021, 94: 103650.
[92] TANGULER H. Evaluation ofinoculum level on fermentation and flavor compounds of white wines made from emir (L) grown in anatolia. Food Biotechnology, 2012, 26(4): 351-368.
[93] ROULLIER-GALL C, BORDET F, DAVID V, SCHMITT-KOPPLIN P, ALEXANDRE H. Yeast interaction on Chardonnay wine composition: Impact of strain and inoculation time. Food Chemistry, 2022, 374: 131732.
[94] 戰吉宬, 曹夢竹, 游義琳, 黃衛東. 非釀酒酵母在葡萄酒釀造中的應用. 中國農業科學, 2020, 53(19): 4057-4069. doi: 10.3864/j.issn. 0578-1752.2020.19.018.
ZHAN J C, CAO M Z, YOU Y L, HUANG W D. Research advance on the application of non-in winemaking. Scientia Agricultura Sinica, 2020, 53(19): 4057-4069. doi: 10.3864/j.issn. 0578-1752.2020.19.018. (in Chinese)
[95] LIU H, LI X, DENG J Z, DAI L Y, LIU W, PAN B L, WANG C T, ZHANG D J, LI Z J. Molecular mechanism of the response ofto D-fructose stress by the glutathione metabolism pathway. FEMS Yeast Research, 2020, 20(5): foaa034. doi: 10.1093/femsyr/foaa034.
[96] MBUYANE L L, BAUER F F, DIVOL B. The metabolism of lipids in yeasts and applications in oenology. Food Research International, 2021, 141: 110142.
Research Progress on the Application of Non-During Wine Fermentation
WANG ChunXiao1, YU JunZhu1, ZHOU WenYa1, XU YinHu2
1School of Liquor and Food Engineering, Guizhou University/Key Laboratory of Fermentation Engineering and Biological Pharmacy of Guizhou Province, Guiyang 550025;2National Center for Yeast Technology Research and Promotion, Yichang 443003, Hubei
Non-yeast are naturally present on grape skin, which mainly play their roles in the maceration and initial stage of fermentation during winemaking process. More and more concerns have been recently focused on the application of non-yeast in wine fermentation. Non-yeast often have weaker fermentation ability thanand can convert the reducing sugar into ethanol and other fermentation by-products. Therefore, non-are the potential excellent yeast for producing wine with complex flavor traits and lower ethanol degree. Different non-yeast species have different metabolic characteristics in wine fermentation applications. The selection of non-yeasts with certain characteristics for application in fermentation can improve the specialized quality of wine. This review was based on the initial summary on the species, winemaking traits, and application ways of commercial non-yeast, and emphasized on the positive role of non-in wine color, aroma, taste, safety and health, their metabolic mechanisms and corresponding research hotspot. The positive role of non-in wine color included the promotion of high acid production, high polysaccharide production, high extracellular pyruvate production and low sorption properties. Different non-yeast could promote the fruity aroma enhancement in wine by low production of ethanol, acetaldehyde, and volatile phenols, and high production of ethyl acetate, acetic acid ester, ethyl ester, higher alcohols, and enzymes related with the release of terpene and mercaptan, and therefore improve the aroma complexity of wine. Non-yeast could regulate wine mouthfeel traits by high production of glycerol, polysaccharide, and lactic acid, and degradation of malic acid. In addition, non-yeast could improve the safety quality of wine by decreasing the SO2dosage as biological control agent, and reducing the content of toxic compounds through metabolic degradation during alcoholic fermentation. This review further analyzed the current research status on genome and microsatellite loci analysis. The main inoculation strategies were discussed on application of non-yeast during wine fermentation, and six aspects were posed for further research concern. This review provided theoretical references for the research on application of the non-yeasts in wine alcoholic fermentation.
mixed fermentation; color stability; aroma complexity; genotype; commercialization

10.3864/j.issn.0578-1752.2023.03.011
2022-04-30;
2022-07-22
國家自然科學基金青年科學基金(32160544,31801523)、貴州省科技計劃(黔科合支撐[2022]重點006號)、貴州省教育廳青年科技人才成長項目(黔教合KY字[2018]120)、貴州大學引進人才科研項目(貴大人基合字[2017]44號,貴大培育[2020]50號)、貴州省發酵工程與白酒釀造人才基地(黔人領發[2018]3號)、遵義市創新人才團隊培養項目(遵義科人才[2020]9)
通信作者王春曉,Tel:18111908483;E-mail:cxwang@gzu.edu.cn
(責任編輯 趙伶俐)
猜你喜歡
現代畜牧科技(2021年6期)2021-07-16 05:50:06
河北農機(2020年10期)2020-12-14 03:13:30
今日農業(2019年15期)2019-01-03 12:11:33
新農業(2017年22期)2018-01-03 05:46:00
中國農業文摘-農業工程(2016年5期)2016-04-12 05:38:15
現代農業(2016年5期)2016-02-28 18:42:33
現代農業(2016年5期)2016-02-28 18:42:33
現代農業(2016年4期)2016-02-28 18:42:19
現代農業(2015年5期)2015-02-28 18:40:54
作物研究(2014年6期)2014-03-01 03:39:10
