云南金鐵鎖根腐病病原菌的分離及鑒定
2023-03-13 07:41:22楊文婷張曉梅李國棟李俊楠楊承凡張愛麗
廣西植物 2023年1期
楊文婷, 張曉梅, 李國棟, 李俊楠, 楊承凡, 張愛麗
( 云南中醫藥大學 云南省高校中醫藥學分子生物學重點實驗室/中藥學院暨云南省南藥可持續利用研究重點實驗室, 昆明 650500 )
根腐病是一類具有容易傳染、發病率高、防治困難等特點的根部病害,在根和根莖類藥用植物中的發病率較高(陳茂婷等,2020)。據報道,引起植物根腐病的病原菌主要為鐮刀菌屬真菌(穆向榮等,2014;伍曉麗等,2020;曹敏,2022)。鐮刀菌是一類世界性分布的真菌,可通過破壞植物體結構,侵染多種植物根、莖及果實等部位,引起根腐病、莖腐病和穗腐病等多種病害(謝安娜等,2020)。由鐮刀菌引起的多種土傳病害也正威脅我國三七、人參、黃芪、半夏、川芎、麻黃等重要中藥材的種植生產(朱春雨等,2003;李佳穗等,2015;文增葉等,2019;張靜等,2019;姚天明等,2020;Li et al., 2021)。目前,國內外研究關于鐮刀菌分類地位、形態學與分子學特征、致病性與致病機理及生物防治等方面均有報道(高芬等,2018;梁嘉俊等,2019)。由于鐮刀菌屬形態復雜,易受外界環境影響而發生變異,因此,通過形態學特征很難準確地鑒定鐮刀菌種類,影響了后續防治等工作的開展。隨著分子生物學技術的發展,形態學結合DNA分子片段分析等手段能更加有效地提高鐮刀菌種類鑒定的準確性(王世偉等,2018)。內轉錄間隔區(internal transcribed spacer,ITS)、線粒體小亞基核糖體(mitochondrial small subunit rDNA,mtSSU)、β-微管蛋白(β-tubulin)及翻譯延伸因子(nuclear translation elongation factor-1α,TEF-1α)等rDNA片段已被廣泛地用于鐮刀菌種類的鑒定(Sampietro et al., 2010)。其中,核糖體DNA (rDNA)和TEF-lα延伸因子兩類基因位點常用于進行鐮刀菌屬的種屬鑒定(Tapani et al., 2002)。rDNA是鑒定鐮刀菌的理想方法,其最大優點是既具有保守性,又存在變異性。作為真核生物多肽鏈延伸因子,TEF-1α在種水平上具豐富信息量,較ITS序列有更多的種間變異,具有高分辨力。這種方法與ITS序列聯合分析,可提高對鐮刀菌鑒別的準確性。
金鐵鎖(Psammosilenetunicoides)是石竹科(Caryophyllaceae)金鐵鎖屬(Psammosilene)植物,首載于《滇南本草》(蘭茂,2004),主要分布在中國云南、貴州、西藏等省區(朱常成等,2007)。金鐵鎖以根入藥,主治跌打損傷、風濕、癰疽瘡癤、創傷出血等(胡成剛等,2002)。在苗族、彝族、白族等少數民族中有著悠久的用藥歷史,其有效成分為齊墩果烷型三萜皂苷類化合物,在鎮痛抗炎、調節免疫、殺菌抑菌等方面具有較好的作用(王學勇等,2006;周欣等,2013),是“云南白藥”等多種著名中成藥的主要原料藥物之一(周祖英等,2021)。近年來,由于長期的采挖,金鐵鎖野生資源急劇減少(朱常成和尹子麗,2017),現已被列為國家二級保護植物,被《中國植物紅皮書》收錄(傅立國,1991)。
目前,云南大部分地區都有種植金鐵鎖(呂金富等,2018;羅映菊,2018)。課題組前期對曲靖馬龍、玉溪華寧、大理劍川及麗江華坪等云南各地區金鐵鎖栽培基地進行了走訪調查,發現二年生、三年生的金鐵鎖在7—8月雨季時極易發生根腐病,多年的連作方式使土壤中的病原菌積累,導致土壤環境復雜,根腐病害的發生逐年加重,農戶因怕遭受病害導致損失而采挖一年生的金鐵鎖,但一年生金鐵鎖因栽培年限不夠又導致藥材浸出物檢驗不合格(楊麗云等,2020),因此,極大地影響了金鐵鎖藥材品質,很大程度上限制了云南白藥等企業對金鐵鎖資源的開發及利用。然而,目前未見金鐵鎖根腐病的相關報道,嚴重影響了其防治工作的開展。鑒于此,本研究結合形態學觀察及基因序列分析系統地鑒定了金鐵鎖根腐病的致病菌,首次發現尖孢鐮刀菌是引起金鐵鎖根腐病的主要病原菌之一,研究結果可為下一步開展金鐵鎖根腐病生物防治功能菌株資源的挖掘奠定基礎,并為金鐵鎖的病害診斷和防治提供科學理論依據。
1 材料與方法
1.1 材料
植物材料:金鐵鎖病害樣品及健康植株(健康樣品和病害樣品都為二年生金鐵鎖植株的根)于2020年11月采集于云南馬龍,該地年均氣溫10~21 ℃,年均降水量979.6~1 001.8 mm。該金鐵鎖栽培基地的土質疏松肥沃,為金鐵鎖提供了良好的生長條件。金鐵鎖組培苗來自云南中醫藥大學中藥材優良種苗繁育工程中心。
主要試劑和儀器:馬鈴薯葡萄糖瓊脂培養基(PDA)(Solarbio,生產批號為428G031),馬鈴薯葡萄糖肉湯(PDB) (Solarbio,生產批號為601Y031),萘啶酮酸(Solarbio,生產批號為Y77408),Taq酶(Takara,生產批號為AL52014A),真菌DNA小量提取試劑盒(Magen,生產批號為DKI09-01)。立式高壓滅菌鍋(Uniclave,FD50A型號),超凈工作臺(蘇凈安泰,SW-CJ-FD型號),生化培養箱(Bluepard,LRH-70型號),生物顯微鏡(Nikon,ECLIPSE E100型號)。測序引物由擎科生物(上海)科技有限公司合成。
1.2 方法
1.2.1 病原菌的分離純化 取有根腐病癥狀的根的病健交界處組織,在5%NaClO中表面消毒1.5 min,然后在75%酒精中表面消毒1.5 min,無菌水沖洗2~3次,無菌濾紙吸干水分,用無菌剪刀在根樣品的病健交接處剪1/3指甲蓋大小的組織,將有切口的一面貼到含50 mg·L-1萘啶酮酸的PDA培養基上(50 mg·L-1萘啶酮酸可選擇性抑制細菌生長),并在28 °C下培養2~3 d,待染病組織塊周圍長出菌絲時,用接種針挑取菌落邊緣尖端菌絲進行純化,并觀察菌落形態。多次純化后的菌株根據菌落大小、形態、顏色等特征去重復后接種于PDA斜面-4 ℃保存。
1.2.2 致病性測定
1.2.2.1 病原菌分生孢子懸浮液的制備 將保留的菌種在PDA平板上活化后轉接至PDB培養基中,放在搖床上28 °C培養7 d,然后將液體培養基中培養好的菌株用4層紗布過濾除去菌絲,5 000 r·min-1離心10 min后,棄上清液,取沉淀物加入適量的無菌水搖勻,制備成濃度約為1.0×106cfu·mL-1的孢子懸浮液備用。孢子懸浮液的濃度采用血球計數板法計算(張繼忠和王薇,2018)。
1.2.2.2 病原菌的回接 組培苗接種病原菌:提前準備好金鐵鎖組培苗,每個組培瓶里應具有10個以上的金鐵鎖根。使用注射器往金鐵鎖組培苗的須根周圍接種5 mL分生孢子懸浮液,重復接種6瓶;對照組采用同樣方法接種5 mL無菌水,重復接種6瓶,然后室溫中培養,觀察組培瓶內金鐵鎖組培苗的變化,連續觀察1個月。
盆栽接種病原菌:前期把采回的二年生金鐵鎖移栽置塑料花盆(直徑19 cm,高19 cm)內。采用浸泡法接種,將健康金鐵鎖的根置于培養皿中,往培養皿中注入分生孢子懸浮液,浸泡0.5~1 h后倒出分生孢子懸浮液,處理后的根立即種于裝有滅菌土的塑料花盆內,覆土。對照處理用無菌水浸泡相同時間后立即栽種。重復用分生孢子懸浮液接種60株金鐵鎖,然后正常的管理培養。培養期間,觀察盆栽中金鐵鎖苗的生長狀況,待葉片枯黃脫落后挖出金鐵鎖根,觀察其是否爛根。從發病植株的病健交界處再次進行病原菌分離,驗證病原菌的致病性。
1.2.3 病原菌的鑒定 形態學鑒定:將菌株接種于PDA培養基上進行活化,期間觀察PDA培養基上的菌落的形態特征(大小、顏色、形狀的變化),并用接種針挑取少量菌絲放置于載玻片上,加入幾滴無菌水,蓋上蓋玻片制成臨時裝片,采用Nikon生物顯微鏡,在1 000倍鏡下觀察分生孢子的形狀、大小、厚垣孢子的有無及著生方式。分子生物學鑒定:將病原菌接種到PDA培養基上,待菌絲長至旺盛,挑取適量菌絲體于1.5 mL無菌管中,采用真菌DNA小量提取試劑盒進行病原菌DNA提取。對病原菌DNA進行內轉錄間隔區(internal transcribed spacer,ITS)和轉錄延伸因子-1α(transcription elongation factor-1α,TEF-1α)的擴增。引物名稱及序列:ITS1 (TCCGTAGGTGAACCTGCGG)/ITS4 (TCCTCCGCTTATTGATATGC);EF1α-EF (GCTCCYGGHCAYCGTGAYTTYAT)/EF1α-ER (ATGACACCRACRGCRACRGTYTG)。反應體系(50 μL):Takara Premix Taq擴增酶 45 μL,正向引物(10 μmol·L-1) 2 μL,反向引物(10 μmol·L-1) 2 μL,DNA模板1 μL。反應條件:98 ℃ 預變性2 min;98 ℃變性10 s,ITS 56 ℃ ( TEF-1α 54 ℃) 退火10 s,72 ℃延伸 (根據片段長度按照10 s·kb-1進行設置),30個循環;72 ℃ 延伸5 min。PCR產物經凝膠電泳檢測后委托擎科生物(上海)科技有限公司進行測序。將測序結果與NCBI數據庫中的已知序列進行BLAST比對,使用MEGA 5.0 (Tamura, et al., 2011)將序列拼接起來,采用貝葉斯法(Bayesian Inference,BI)對病原菌進行多基因(TEF-1α和ITS)聯合建樹的方法構建系統發育樹(Ronquist & Huelsenbeck, 2003; Maharachchi-kumbura, et al., 2012)。
2 結果與分析
2.1 金鐵鎖根腐病癥狀
前期我們對金鐵鎖栽培基地的調研發現,金鐵鎖根腐病的葉片癥狀很難見到,一般根腐病發生后,植株葉片早已經完全枯萎脫落,導致我們采樣時很難在田間見到金鐵鎖感染根腐病后的地上部分癥狀。實驗室進行金鐵鎖染菌試驗后,發現感染根腐病后的金鐵鎖葉片會脫綠,逐漸黃化,直至枯萎、脫落、死亡。金鐵鎖根部變化首先是根表皮萎縮,暗黃色加深直至變為黑褐色,之后軟化,根部開始從根尖慢慢向根上部開始腐爛,表面有時會長出白色絲狀物,可能是病原菌的菌絲(圖1)。

A. 田間完全腐爛的根; B. 田間部分腐爛的根; C. 根部病健交接處的組織。A. Completely rotten roots in the field; B. Partially rotten roots in the field; C. Tissues at the junctions of disease and health of the roots.圖 1 金鐵鎖根腐病的癥狀Fig. 1 Symptoms of Psammosilene tunicoides root rot
2.2 金鐵鎖根腐病病原菌的分離及純化
對金鐵鎖根腐病的根系組織進行分離及純化,純化后共得到85株真菌,85株真菌的菌落在PDA培養基上總共呈現三種形態(圖2):第一種絨毛蓬松,呈輪紋狀,布滿整個培養基,白色(圖2:A);第二種呈絨毛狀較貼培養基,形狀不規則,白色(圖2:B);第三種有輪紋狀,內圈為黃褐色絨毛,較貼培養基,外圈為灰白色蓬松絨毛(圖2:C)。

圖 2 分離純化后的真菌菌落形態特征Fig. 2 Morphological characteristics of fungal colonies after isolation and purification
2.3 金鐵鎖根腐病病原菌的致病性測定
我們對分離純化后的真菌去重復后進行了致病性測定,金鐵鎖組培苗分別接種了病原菌PSD-1、PSD-2、PSD-3的分生孢子懸浮液,3 d后長出菌絲,1周后組培苗從根部開始,朝著莖葉方向發黃,2周后組培苗全部變黃,死亡,而對照組仍無癥狀。重復試驗后獲得相同的結果。
金鐵鎖盆栽苗分別接種了病原菌PSD-1、PSD-2、PSD-3的分生孢子懸浮液,1個月后,葉子開始萎蔫但沒有褪色,在中午前后光照強、蒸發量大時,植株上部葉片出現萎蔫,但夜間又能恢復。繼續觀察一段時間后,葉片萎蔫狀況夜間也不能再恢復,葉片的綠色逐漸褪去,直至變成黃白色,葉片開始慢慢脫落。2個月后,待葉片完全枯萎脫落后挖出金鐵鎖的根,發現根部顏色比栽種之前更深,根部軟化、腐爛,與田間患病植株根的癥狀相同,而對照組仍無癥狀。重復試驗后獲得相同的結果。發病率分別為60%、61.7%、71.7% (發病率=發病植株/植株總數×100%)。并且,從發病金鐵鎖根上均能再分離到與初接菌株相同的分離物,確定為金鐵鎖根腐病病原菌,因此判斷病原菌PSD-1、PSD-2、PSD-3對金鐵鎖的致病力較強。
2.4 金鐵鎖根腐病病原菌的形態鑒定
PSD-1接種于PDA培養基上,28 ℃培養5 d后,菌落呈近圓形,菌絲體較細,呈疏松絨毛狀較貼培養基,菌落有輪轉紋,內圈呈淡黃褐色,外圈邊緣白色,隨時間的推移內圈顏色逐漸朝外圈變深,菌落背面邊緣呈白色,中間呈黃褐色,菌落直徑約為3.4 cm;小型分生孢子為卵圓形,著生于單出的分生孢子梗上;分生孢子梗細長;大型分生孢子較多,5~6分隔,多為新月型;厚垣孢子近球形,多生于菌絲的頂端或中間,壁表光滑(圖3)。
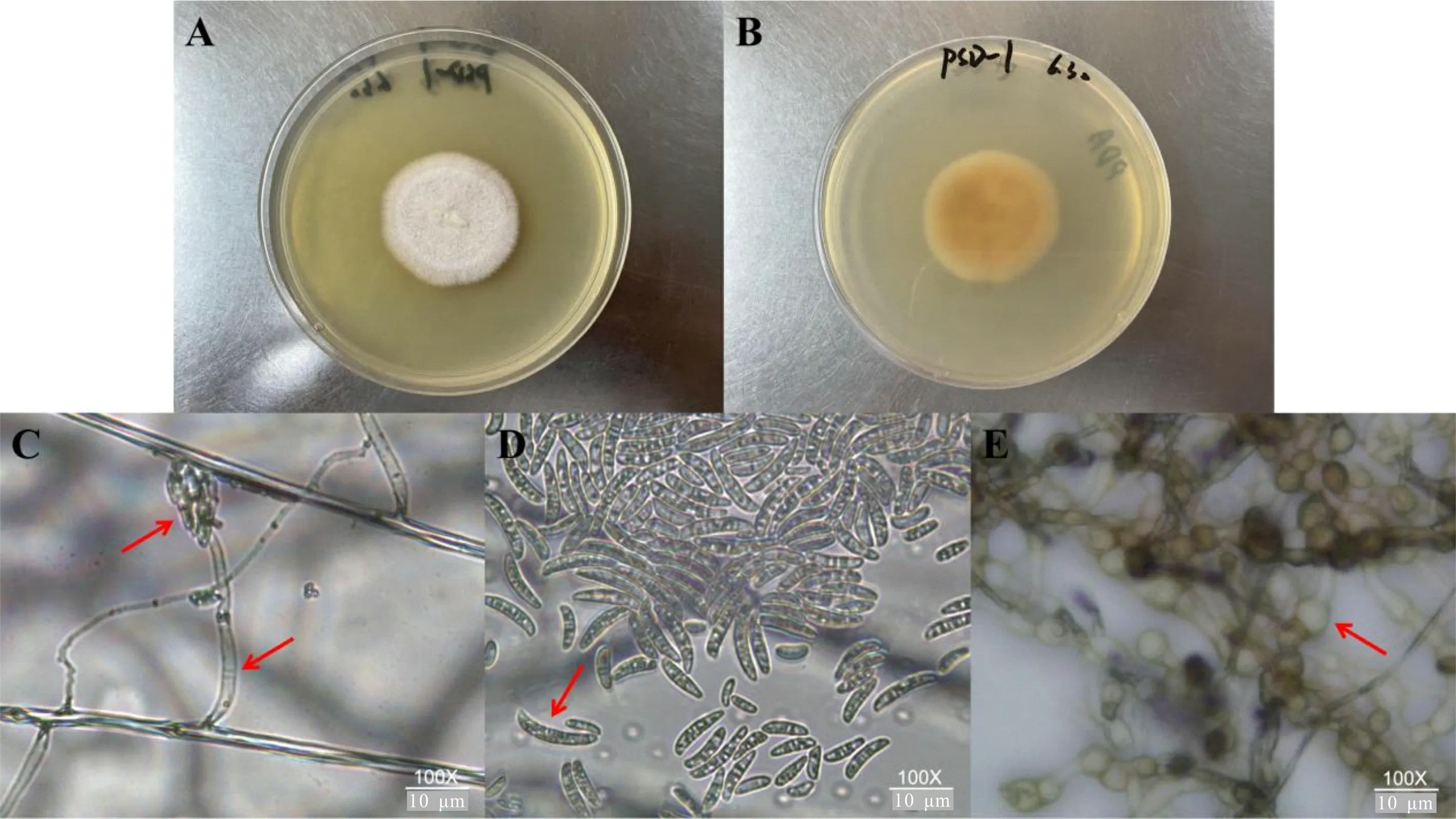
A, B. PSD-1的菌落形態; C. 小型分生孢子及分生孢子梗; D. 大型分生孢子; E. 厚垣孢子。B-D標尺: 100×10 μm。A, B. Bacterial colony morphology of PSD-1; C. Microconidia and conidiophores; D. Macroconidia; E. Chlamydospores. B-D scale bars: 100 × 10 μm.圖 3 以PSD-1為代表的菌落和菌體形態Fig. 3 Colony and morphology with the represent of PSD-1
培養5 d后,PSD-2菌落呈不規則橢圓形,邊緣呈鋸齒狀,菌絲較長,絨毛狀,質地棉絮狀,白色,菌落背面大部分呈白色,中間偶見黃色細輪紋,菌落長軸為約3.7 cm,短軸為約2.6 cm;小型分生孢子為腎形,偶見2~5分隔,著生于分生孢子梗上;分生孢子梗細長,呈樹枝狀分生;大型分生孢子4~7分隔,呈鐮刀型;厚垣孢子橢圓形,壁表光滑或粗糙。PSD-3菌落呈類橢圓形,邊緣不整齊,菌絲濃密,質地棉絮狀,白色,菌落背面顏色呈白色,中間有黃色小點,菌落長軸為約3.3 cm,短軸為約2.8 cm;小型分生孢子呈腎型至卵圓形,著生于分生孢子梗上;分生孢子梗細長、分枝較少;大型分生孢子多為1~3分隔,為卵形;厚垣孢子呈不規則球形,多生于菌絲的頂端,壁表光滑。
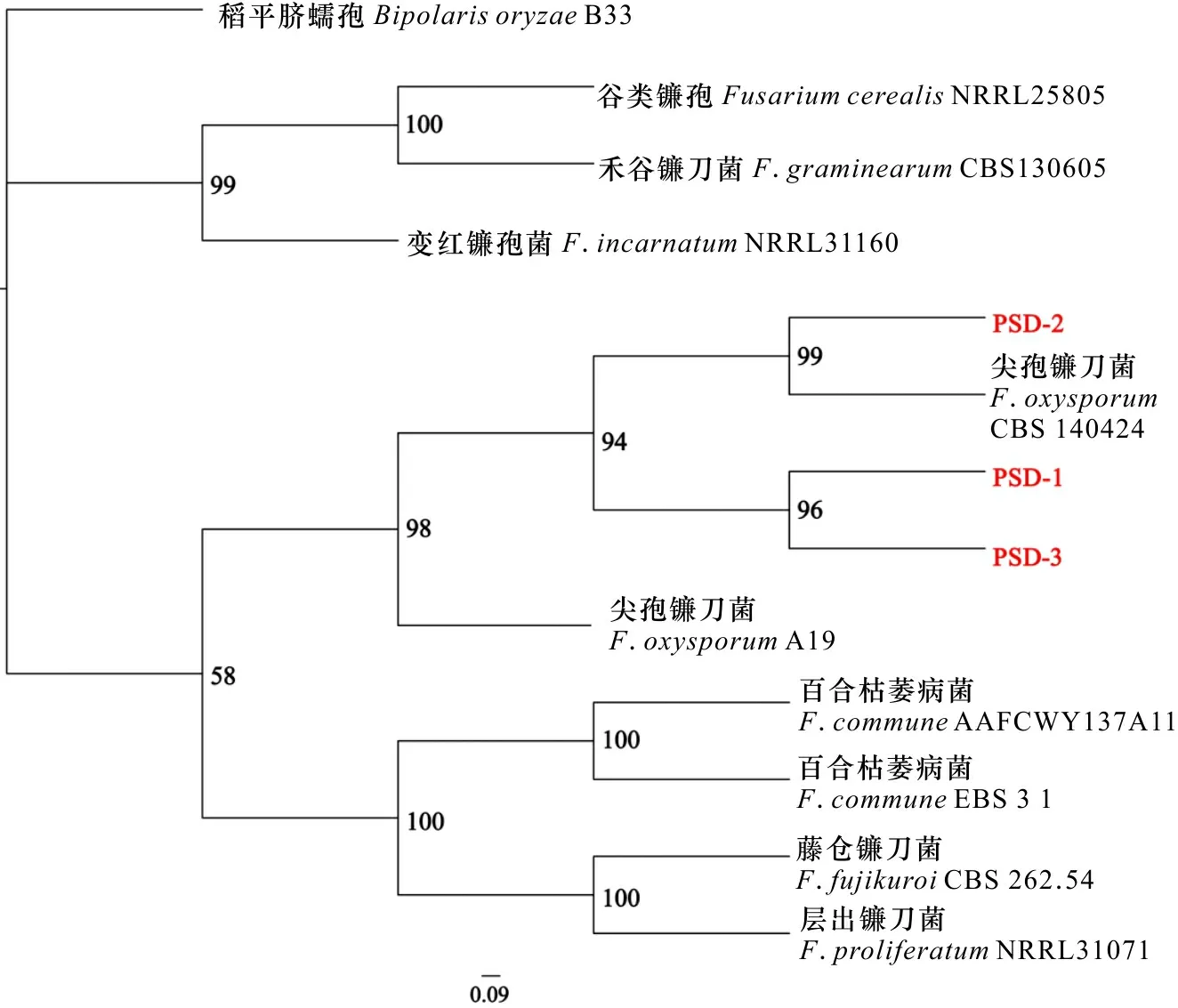
編號PSD-1、PSD-2、PSD-3的菌株是從根腐病的金鐵鎖中分離而得到。The strains numbered PSD-1, PSD-2 and PSD-3 were isolated from Psammosilene tunicoides with root rot.圖 4 基于TEF-1α和ITS序列構建的Fusarium oxysporum的貝葉斯(BI)系統發育樹Fig. 4 Bayesian inference phylogenetic tree of Fusarium oxysporum based on TEF-1α and ITS sequences
2.5 金鐵鎖根腐病病原菌的分子生物學鑒定
經真菌ITS和TEF-1α序列分析表明,PSD-1、PSD-2、PSD-3與尖孢鐮刀菌(Fusariumoxysporium)相似性在99%以上。基于ITS和TEF-1α構建的系統發育樹發現,菌株PSD-1、PSD-2、PSD-3都與尖孢鐮刀菌A19聚為一支,支持值達98%,并與鐮刀屬的其他菌株分支明顯,其中菌株PSD-2與模式菌株尖孢鐮刀菌CBS 140424聚到一起,支持值達99%。基于上述多基因聯合構建的供試菌株與尖孢鐮刀菌系統發育樹結果和形態學特征,將PSD-1、PSD-2、PSD-3鑒定為尖孢鐮刀菌(圖4)。
3 討論與結論
根腐病是一種常見的土傳病害,對中草藥根和莖具有毀滅性的危害(廖長宏等,2017)。目前,大量研究表明,一些鐮刀屬真菌是引起藥用植物根腐病的主要病原菌之一(陳茂婷等,2020)。本研究通過病原菌分離、柯赫氏法則驗證(黃幸鴿等,2021)及病原菌形態學鑒定和分子生物學鑒定,首次分離鑒定了金鐵鎖根腐病的一種主要致病真菌為尖孢鐮刀菌。將分離得到的病原菌分別接種至健康的組培苗和盆栽苗,結果發現健康植株產生了與大田植株根腐病相似的癥狀,發病率分別為60%、61.7%、71.7%。這說明尖孢鐮刀菌是引起金鐵鎖根腐病的主要病原菌之一。Guo等(2009)研究發現,引起三七根腐病的病原菌主要為腐皮鐮刀菌(F.solani)、尖孢鐮刀菌、惡疫霉病菌(Phytophthoracactorum)和草莖點霉 (Phomaherbarum)等。張禮維(2015)結合形態及分子鑒定,分離得到的61株白術的根腐病病原菌均為尖孢鐮刀菌。由此可見,尖孢鐮刀菌是引起植物根腐病中較為常見的一種鐮刀菌屬的病菌。我們對云南馬龍栽培基地的金鐵鎖病原菌進行鑒定,分離得到的主要病原菌也為尖孢鐮刀菌,這與他人的研究結果一致。然而,前人的研究中,許多植物的根腐病多由多種鐮刀菌復合侵染所致,本研究中分離得到的85株真菌,均為尖孢鐮刀菌,未見其他種類的病原菌。主要原因有兩個:一是可能本研究采集病害的樣本不夠廣泛,僅分離了云南馬龍栽培基地的病害金鐵鎖,導致分離獲得的病原菌比較單一;二是尖孢鐮刀菌是大多植物根腐病致病菌中的優勢病原菌種,相對分離頻率較高。陳思杰等(2021)研究亦發現,分離得到寧夏枸杞的致病菌中,尖孢鐮刀菌在分離出的鐮刀菌中占據優勢,其他鐮刀菌分離頻率相對較低。
同時,本研究基于ITS和TEF-1α分子片段構建的系統發育樹,結果發現菌株PSD-1、PSD-2、PSD-3都與尖孢鐮刀菌A19聚為一支,并與鐮刀屬的其他菌株分支明顯,其中菌株PSD-2與模式菌株尖孢鐮刀菌CBS 140424聚到一起,支持值達99%。鐮刀菌屬是真菌中最難鑒定的種屬之一,其由于形態復雜,在生長過程中形態受環境的影響變異較大,單純依據形態學很難準確鑒定到種(邢安,2017)。在本研究中,亦發現菌株PSD-1、PSD-2、PSD-3雖然都為尖孢鐮刀菌,但菌落形態還存在一定的差異。近年來利用分子標記技術輔助鑒定鐮刀菌的方法已被廣泛應用,可以彌補形態學鑒定的不足。曹瑱艷等(2020)使用鐮刀菌種特異性引物并結合ITS和TEF序列分析及形態學鑒定確定了鐵皮石斛根腐病的4種病原真菌。遲惠榮(2019)根據形態學、rDNA-ITS和TEF-1α序列同源性分析,確定了導致多花黃精葉枯病的病原菌為尖孢鐮刀菌。由于鐮刀菌種類繁多,形態變異復雜,包括很多親緣關系較近的種及復合種,所以單一片段很難對其準確鑒定。本研究在形態學鑒定的基礎上,結合ITS和TEF-1α序列進行聯合分析,首次鑒定引發金鐵鎖根腐病的主要病原菌為尖孢鐮刀菌,并得到了可靠的分類鑒定結果。
綜上所述,本研究調查了云南省金鐵鎖根腐病的發生狀況,首次分離并鑒定引起金鐵鎖根腐病的主要病原菌為尖孢鐮刀菌。金鐵鎖根腐病的防治是下一步工作的重點,其中生物防治菌株的挖掘和篩選是金鐵鎖根腐病防治的重要途徑。生防菌較化學農藥而言,具有能改善環境污染、降低農藥殘留、保護生態環境等優點。近年來,基于高通量組學測序技術挖掘與驗證藥用植物的生防菌已成為科學研究的熱點。通過微生物組學和宏基因組學分析,探究金鐵鎖根腐病與根際土壤微生物群落的關系,篩選出用于生物防治劑的候選菌株,并通過純培養手段獲得并富集拮抗活性菌株,為金鐵鎖根腐病的防治提供了新思路。同時,基于現在云南省金鐵鎖的栽培現狀,結合栽培措施(選育優良抗病品種,脫毒中藥種苗的規模化應用,對土壤進行消毒處理,使用土壤改良劑,進行合理的輪作)、化學防控(使用甲霜靈等殺菌劑)和生物防控(引入拮抗菌和接種有益微生物)等多種措施聯合使用,能更有效地防治金鐵鎖的根腐病。
