普通菜豆遺傳育種研究進展
2023-03-14 11:44:12羅安紅李美樂董佳文
蔬菜 2023年3期
關鍵詞:資源
羅安紅,李美樂,劉 芳,符 瓊,董佳文
(湖南應用技術學院,湖南 常德 415000)
普通菜豆(Phaseolus vulgarisL.,2n=2 x=2 2)又名四季豆、蕓豆、飯豆,為豆科(Leguminosae)、蝶形花亞科(Papilionoideae)、菜豆屬(Phaseolus)的1年生草本植物。普通菜豆是世界上種植面積最大的食用豆類,據報道,全世界種植菜豆的國家和地區(qū)共有90多個,總種植面積達2 878.04萬hm2,占食用豆類的40%,總產量達2 314.03萬t,占全部食用豆類總產量的30%。截至2018年底,我國普通菜豆種植面積達1 000萬k m2,總產量達180萬t[1]。
在19世紀末期之前,人們認為普通菜豆起源于亞洲,如Linnaeus曾提出普通菜豆發(fā)源地為印度,而科研工作者多方面研究表明,普通菜豆起源于中、南美洲。經種子貯藏蛋白標記、分子標記、形態(tài)學研究發(fā)現,野生菜豆集中分布于2個區(qū)域:即位于秘魯南部、玻利維亞和阿根廷的安第斯中心和包括墨西哥、危地馬拉、巴拿馬、洪都拉斯、尼加拉瓜、哥斯達黎加和哥倫比亞在內的中美洲區(qū)域。2個中心交匯于哥倫比亞與委內瑞拉地區(qū),該區(qū)域許多馴化材料的蛋白類型為雜合體[2]。
普通菜豆在起源中心完成馴化后陸續(xù)傳入世界各地。根據種子貯藏蛋白標記,可推斷普通菜豆的傳播路徑。菜豆于16世紀初傳播到歐洲,之后,再經歐洲傳入中東、西亞及世界上其他國家和地區(qū)。普通菜豆是在15世紀由美洲直接傳入中國的,之后于1654年由隱元禪師傳入日本[3]。我國普通菜豆種質資源豐富,但針對現有種質資源的利用及創(chuàng)新仍較為缺乏,且育種工作與其他主要蔬菜相比相對滯后,本文將介紹我國普通菜豆種質資源研究及創(chuàng)新情況,并對未來發(fā)展方向作出展望,以期為普通菜豆種質資源研究提供參考。
1 我國普通菜豆種質資源的收集鑒定
1.1 我國普通菜豆種質資源的分布及收集保存
中國作為菜豆的次級起源中心,從15世紀直接由美洲引入菜豆。我國普通菜豆種質資源分布比較廣泛,但分別以東北、西北和云貴高原為主要分布中心。目前中國栽培的普通菜豆大多為地方品種,主要分布于云南、貴州、山西、陜西、甘肅、內蒙古、黑龍江、吉林等省區(qū)[4]。編入種質資源目錄的資源占有量比較多的省份依次為:貴州1 003份、山西594份、云南461份、黑龍江381份、陜西379份、四川336份、內蒙古238份、河北151份、吉林106份和湖北101份[5]。
1.2 我國普通菜豆種質資源的鑒定評價
王述民等[5]對已入庫的3 328份普通菜豆種質材料的39%進行了抗病蟲鑒定,18%進行了抗逆性評價,最終篩選出21個抗旱品種、16個抗寒品種及39個抗病蟲品種。雷蕾等[6]對332份入庫菜豆資源進行炭疽病和枯萎病的苗期抗病性鑒定,對328份菜豆嫩秧作營養(yǎng)成分分析,并對“七五”期間通過抗病性鑒定和營養(yǎng)養(yǎng)分分析的218份材料進行重復鑒定,最終篩選出12份優(yōu)良普通菜豆種質資源。薛仁風[7]、徐新新[8]分別采用下胚軸雙孔注射和接種體蘸根2種病原菌接種方法,對159份普通菜豆資源進行了初步的細菌性疫病抗病性篩選,其中前者篩選出6個高抗品種、11個抗病品種、55個中抗品種,而后者篩選出3個高抗品種、5個抗病品種、10個中抗品種;另外,薛仁風[7]研究表明,我國擁有十分豐富的抗鐮孢枯萎病普通菜豆資源,通過選育抗病新品種來控制菜豆枯萎病前景廣闊。徐新新[8]通過改良梅花針接種法,對602份菜豆種質資源進行抗普通細菌性疫病鑒定,按1.0~10.0級病級評價標準,共篩選出10份1.1~3.0級資源,占總數的2%;192份3.1~5.0級資源,占總數的32%;306份5.1~7.0級資源,占總數的51%;94份7.1~10.0級資源,占總數的15%,其中未發(fā)現免疫或高抗品種。李龍[9]利用隸屬函數分析法,對121份種質的芽期抗旱性進行綜合評價,篩選出躍進豆、白扁豆等芽期抗旱性種質,為普通菜豆抗旱生理與機制研究、抗旱育種奠定了基礎。
2 普通菜豆性狀遺傳規(guī)律研究
2.1 普通菜豆主要質量性狀遺傳規(guī)律研究
普通菜豆質量性狀主要有花色、莢形、莢色等,均被1對等位基因控制,同一性狀、不同表現型的純合親本雜交,其F1代均表現顯性性狀(蔓生、紫色、白花等),且F2代顯性性狀與隱性性狀呈現3∶1的分離比例,符合基因分離規(guī)律。具體遺傳規(guī)律見表1[10],質量性狀遺傳規(guī)律研究對親本選擇、后代分離性狀預測及雜種鑒定具有十分重要的意義。

表1 菜豆主要質量性狀遺傳規(guī)律
2.2 普通菜豆抗病遺傳規(guī)律研究
普通菜豆主要病害有細菌性疫病(Xanthomonas phaseoli)、炭疽病(Colletotrichum lindemuthianum)、根腐病(Fusarium solani)、銹病(Uromyces appendiculatus)、枯萎病(Fusariumf.sp.Phaseoli oxysporum)等。細菌性疫病是影響普通菜豆生產的最主要的病害之一。Silva等[11]研究表明,普通菜豆對細菌性疫病的抗性為受多基因控制的數量遺傳。Coyne等[12]研究表明,菜豆對普通細菌性疫病的抗性為數量遺傳,具有中低遺傳力,其抗性是由一些未知基因所控制,且與植株的生長習性及晚熟程度等有關。Musaana等[13]研究發(fā)現,普通菜豆細菌性疫病抗性是由1~5個加性效應基因控制,與其抗性相關的基因數量與寄主親本及寄主評價時期有關。截至目前,在普通菜豆的11個連鎖群上至少發(fā)現了25個與普通菜豆細菌性疫病抗性相關的QTL標記[14]。在已發(fā)現的QTL中有3個最重要的SCAR標記,即SAP6、SU91、BC420。利用這3種SCAR標記對10份普通菜豆種質資源進行了遺傳分析,研究表明含有這3種SCAR標記的種質資源均表現出較高的抗性,而感病資源則不含這3種標記;因而,可以根據這些SCAR標記分析普通菜豆基因型,從而提高抗病鑒定的效率[15]。
目前對菜豆抗炭疽病的研究比較深入,已經發(fā)現了17個菜豆抗炭疽病基因,其中16個抗病基因是由國外研究發(fā)現[16]。有研究表明,在國內菜豆高抗材料F2533中發(fā)現1個抗炭疽病基因,但沒有對該基因進行嚴格的等位性鑒定[17-20](表2)。
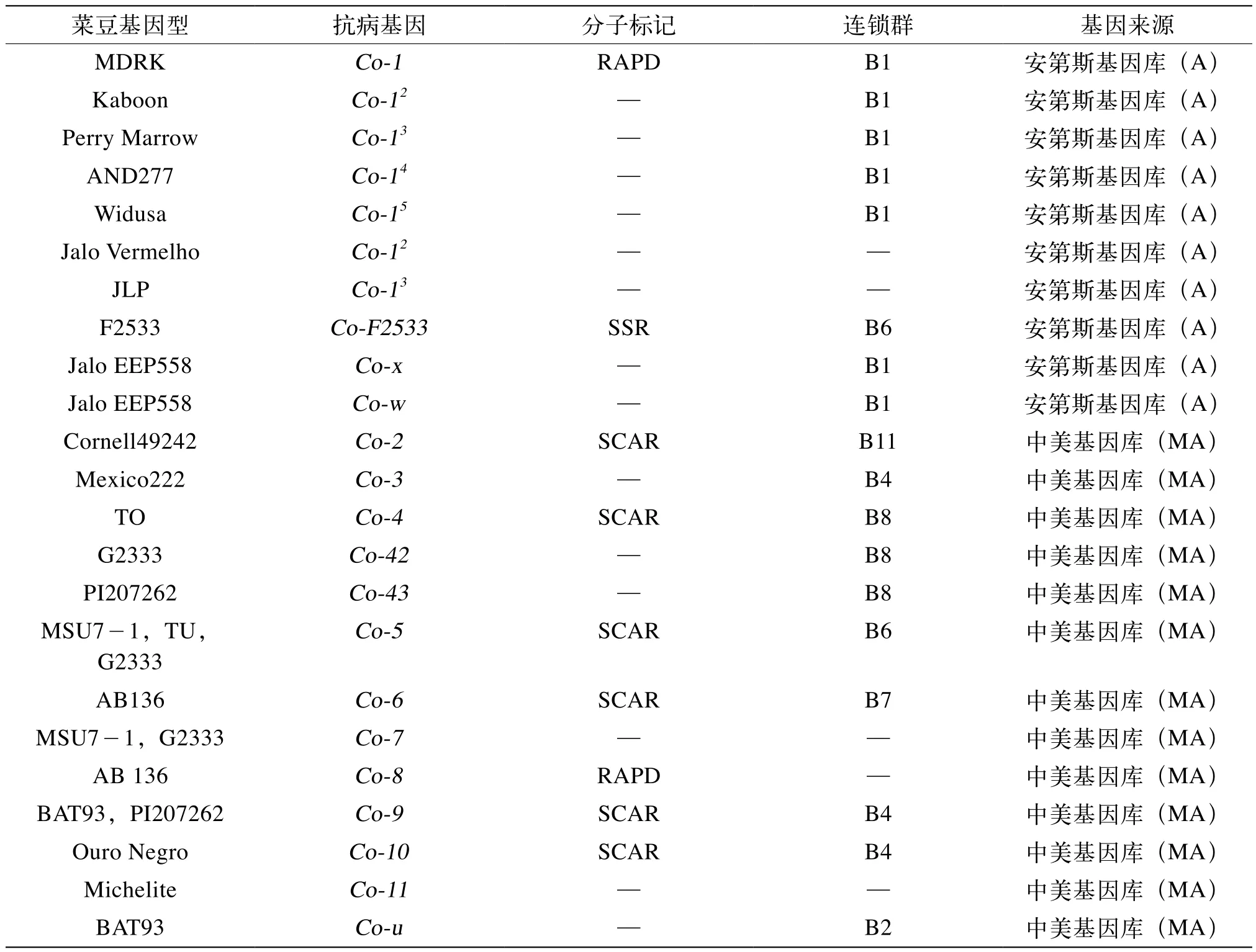
表2 普通菜豆抗炭疽病基因
根腐病也是普通菜豆的常見病害,研究其抗性遺傳規(guī)律對制定培育抗性品種的策略尤為重要。Mukankus等[21]研究表明,菜豆根腐病的抗性由加性效應和非加性效應基因共同控制,將抗性親本用作雜交母本時,后代明顯更具抗性。此外,MLB-49-89A、MLB-48-89、RWR719和Vuninkingi在雜交中表現出較高的抗性水平,可應用于今后的普通菜豆根腐病抗性育種。
銹病是一種破壞性較大的病害,可導致普通菜豆種子產量和豆莢質量顯著下降。普通菜豆的銹病抗性由單一顯性基因Ur決定。迄今為止,已有10個基因被命名和標記,大多采用RAPD或SCAR分子標記,其中5個基因(Ur-3、Ur-5、Ur-7、Ur-11和Ur-14)屬于中美洲基因庫,另外5個基因(Ur-4、Ur-6、Ur-9、Ur-12和Ur-13)屬于安第斯基因庫[22]。
普通菜豆對枯萎病不同生理小種的抗性類型既存在單基因控制的質量性狀,也存在多基因控制的數量性狀。生理小種不同,其抗性基因也不同,目前有12個基因被命名標記(Fop1、Fop2、Fop3、Fop4、Fop6、Fop7、Fop8、Fop11、Fop28、Fop30、Fop31及Fop32)[23]。
3 品種選育研究進展
3.1 引種
引種簡便易行且見效快,是常見的育種手段,而引種成功率的高低取決于引種地區(qū)與原產地區(qū)的生態(tài)條件的差異程度,表現為差異程度越高,則引種成功率越高。中國農業(yè)科學院蔬菜花卉研究所和北京市農林科學院蔬菜研究所于1979年從荷蘭引進菜豆新品種碧豐,品種原名為Selka,經2年的區(qū)域試驗,其表現高產、優(yōu)質且抗逆性強,超過了種植面積較大的蕓豐、豐收1號和雙季豆等[24]。潘仁恩等[25]從日本引進金時菜豆、明石菜豆等7個菜豆品種進行比較試驗,最終篩選出高產、優(yōu)質且生長周期短的蛋白菜豆與四季1751菜豆新品種。李江等[26]從委內瑞拉引進了矮秧材料A2007-1,將其田間變異單株通過系統(tǒng)選育,育成了矮秧型菜豆新品種青菜豆3號。
3.2 選擇育種
選擇育種為改良現有品種和創(chuàng)造新品種提供了一種簡便、有效的育種途徑,是獨立培育優(yōu)良品種的手段,也是其他多種育種方法中必不可少的一環(huán)。揚州市蔬菜研究所從當地菜豆變異株中經6個世代系統(tǒng)選育,于1984年培育出高產、抗逆性強的早熟蔓生型菜豆新品種揚白313,1990年4月通過江蘇省品種審定委員會審定[27]。紀效云等[28]從雙豐2號變異后代中經系統(tǒng)選育,育出耐熱菜豆新品種翠蕓2號。韓玉珠等[29]通過系統(tǒng)選育,育成菜豆新品種吉農油豆1號,該品種高產、抗病且品質良好,適于吉林省露地栽培,2011年2月通過吉林省農作物品種審定委員會審定。張合龍等[30]由從河北引進的唐山菜豆經單株系統(tǒng)選育,培育出早熟、豐產且纖維少、品質好的菜豆新品種豐旺。宿遷農業(yè)科學院由農家品種變異單株經系統(tǒng)選育,培育出高產、抗病的優(yōu)良菜豆新品種遷菜豆1號,并于2015年通過江蘇省農作物品種審定委員會鑒定[31]。
3.3 雜交育種
雜交育種可以有預見性地將親本的優(yōu)良性狀集中于雜種后代上,因而成為菜豆育種的主要途徑之一。經過多年的努力,我國育種工作者培育出了一系列性狀優(yōu)良的菜豆新品種,為我國菜豆產業(yè)的發(fā)展作出了巨大貢獻。吉林市農業(yè)科學院以優(yōu)質、寬莢的081自交系為母本,以豐產、抗病的早熟57號為父本進行組合雜交,經過7個世代的系統(tǒng)選擇,最終選育出高產、優(yōu)質且抗病性強的園豐905,又在此基礎上陸續(xù)選育出園豐906、園豐908等性狀優(yōu)良的菜豆新品種。哈爾濱市農業(yè)科學院以商品性好、抗逆性強的將軍油豆為母本,以當地主栽品種紫花油豆為父本,經連續(xù)6代系譜選育,培育出商品性好、纖維少且高抗炭疽病的哈菜豆9號及哈菜豆10號,并以此為基礎陸續(xù)選育出哈菜豆15號、哈菜豆17號等一系列高產、抗病的優(yōu)良品種[32]。中國農業(yè)科學院蔬菜花卉研究所以中蔓生、晚熟、抗銹病品種82-3與蔓生、早熟品種85-1為親本進行雜交,將所得抗銹病單株與國際抗炭疽病鑒定品種Cornell49-242雜交,并導入其抗炭疽病基因,最后將所得抗病株系與品質優(yōu)良的87-B雜交,經室內接種篩選及定向選擇,培育出高產、優(yōu)質、抗銹病及炭疽病的品種連農架豆36號[33]。表3展現了菜豆雜交育種部分品種情況。
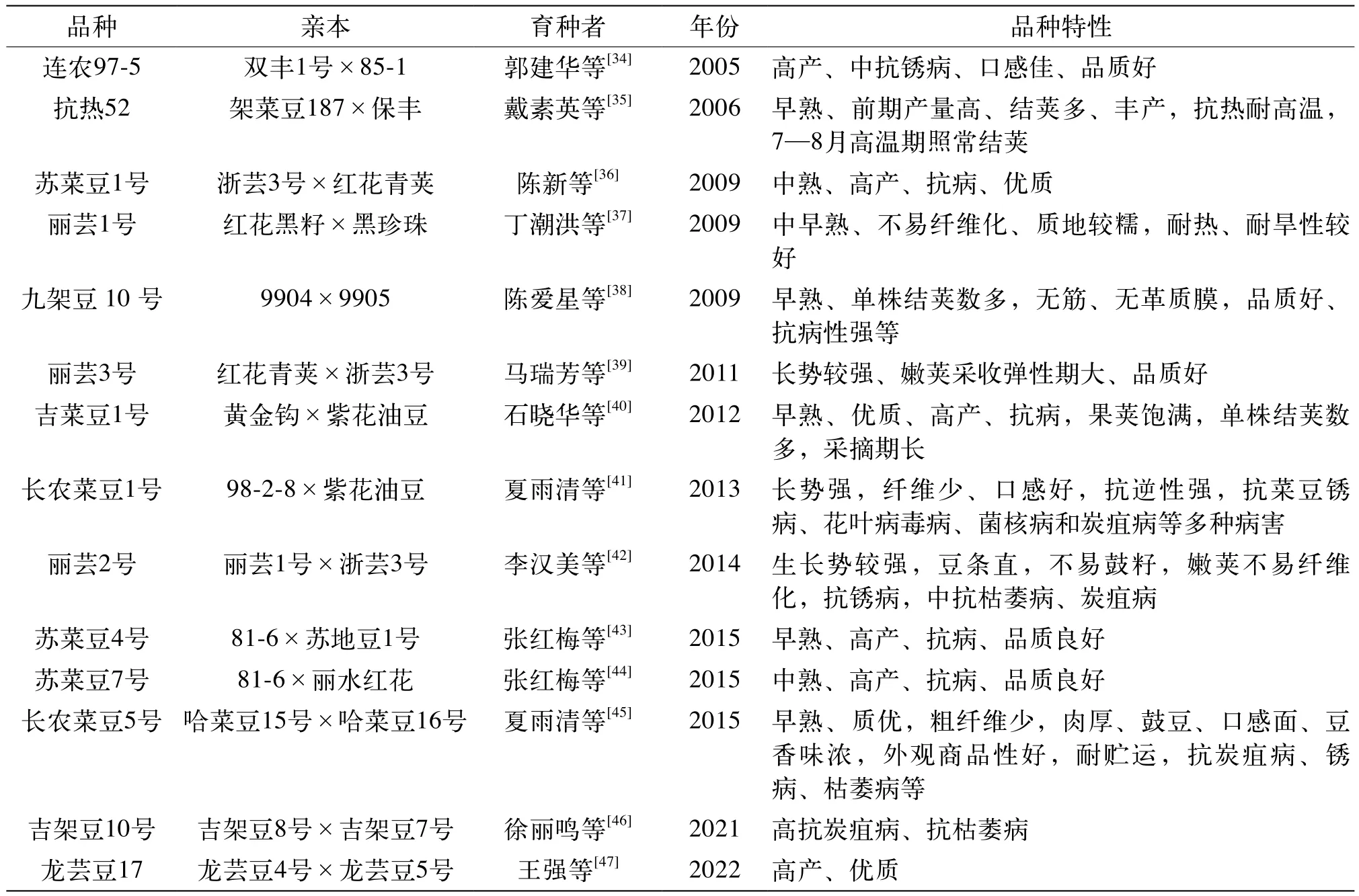
表3 菜豆雜交育種部分品種情況
3.4 其他育種方法
人工誘變育種,即人為利用物理(X射線、γ射線、α射線、β射線、中子、離子束、激光、空間誘變等)或化學(烷化劑、核酸堿類似物、無機化合物、秋水仙素誘變等)等手段誘發(fā)遺傳變異,在短時間內獲得優(yōu)良突變體以育成新品種的育種方法,人工誘變育種具有雜交育種難以替代的優(yōu)勢。張健等[48]將菜豆種子經衛(wèi)星搭載飛行15 d,回收種植后,利用RAPD分子標記分析5個葉片形態(tài)有變異的突變體的群體及親本材料,結果發(fā)現該突變品系與原始對照在DNA水平上確實已發(fā)生了明顯變異。
基因工程育種,即以植物組織、細胞或原生質體為受體系統(tǒng),導入目的基因,改良作物農藝性狀的一種育種方法,使育種目標更明確、操作更直接、程序更精簡。目前,基因工程育種在菜豆上的應用較少。蔣向輝等[49]對菜豆屬和豇豆屬18個栽培種的IST序列進行分析,結果發(fā)現與豇豆相比,菜豆IST序列較長。王全偉等[50]通過RT-PCR得到1個菜豆幾丁質酶基因家族基因Bchi,并通過發(fā)根農桿菌R1000介導轉化煙草,經PCR和Southern blot鑒定獲得了4株轉基因煙草株系。結果發(fā)現Bchi基因在煙草中高水平的表達顯著提高了煙草對真菌性病害的抗性。
隨著現代分子生物學技術的迅速發(fā)展,分子標記輔助育種也被逐步應用于菜豆育種工作中,其可以在分子水平上快速準確地分析個體的遺傳組成,進而實現對基因型的直接選擇。欒非時[51]研究表明,菜豆種內的遺傳多樣性主要分布于蔓生種群內,在所研究的蔓生、矮生、半野生種群中,蔓生種群的遺傳多樣性水平最高,矮生種群其次,半野生種群第3。張赤紅[52]基于SSR標記結果,將377份總樣品聚為6組,其中29份野生菜豆聚在第1組群,與其他樣品無任何交叉;國外16份材料中的11份與我國25份材料聚在第6組。
4 存在問題及展望
我國菜豆種質資源豐富,但育種工作與其他主要蔬菜相比還相對滯后,近幾年在科研人員的不懈努力下,已經選育和引進了一批高產、優(yōu)質且抗性強的特色品種,但從另一個角度看,隨著人民群眾對菜豆品種需求的日益多樣化,新品種選育也存在亟待解決的問題。我國近幾年雖相繼選育出一部分菜豆新品種,但仍很單一,品種更新速度慢,特別是在針對現有種質資源的利用及創(chuàng)新方面仍與國外有顯著差距。
在未來的育種工作中,還應不斷引進新的菜豆種質資源,豐富育種材料,并對搜集來的材料進行深入研究,為菜豆育種提供明確的優(yōu)良種質;以菜豆的全基因組測序完成為契機,結合基因組學、代謝組學及蛋白質組學等技術,對菜豆種質資源進行創(chuàng)新創(chuàng)制,培育出能滿足新時期消費者需求的菜豆新品種,無論對于滿足國內對菜豆花色品種的需求,還是出口創(chuàng)匯都將具有深遠意義。
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
資源節(jié)約與環(huán)保(2022年8期)2022-09-20 02:25:22
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
藝術品鑒(2020年7期)2020-09-11 08:04:44
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
當代貴州(2018年28期)2018-09-19 06:39:04
資源再生(2017年3期)2017-06-01 12:20:59
決策(2015年9期)2015-09-10 07:22:44
