高寒草地植物多樣性與Godron 群落穩定性關系及其環境驅動因素
2023-03-21 09:18:06雷石龍廖李容王杰張路葉振城劉國彬張超
草業學報 2023年3期
雷石龍,廖李容,王杰,張路,葉振城,劉國彬,張超
(1. 西北農林科技大學水土保持研究所,陜西 楊凌 712100;2. 中國科學院水利部水土保持研究所,陜西 楊凌 712100;3. 中國科學院大學,北京 100049)
作為生物多樣性重要組成部分,植物多樣性在維持群落穩定性和生態系統功能方面起著至關重要的作用[1]。近幾十年來,植物多樣性與群落穩定性之間關系及驅動機制一直是生態學的研究熱點[2]。研究表明,植物多樣性高的群落,其群落穩定性也較高[3],也有研究發現群落穩定性與植物多樣性在一定環境下呈負相關關系[4],一些學者也在不同生境中證明了兩者之間并非簡單的相關關系[5-6]。目前植物多樣性與群落穩定性之間的關系仍存在爭議。因此有必要開展不同環境梯度下植物多樣性與群落穩定性相關關系的研究,揭示植物群落穩定性的維持機制。
草地是陸地上分布最廣的生態系統,在生物多樣性保護、全球變化應對、碳平衡維持,以及食物的供應中發揮著重要作用[7]。作為“地球第三極”,青藏高原擁有全球最大的高寒草地,面積約16.5×105km2,占中國草地面積的42%[8]。高寒草地作為青藏高原主要的植被類型,其生長季短[9]、自我調節能力弱[10]、群落結構簡單但類型復雜[11],是我國最脆弱的生態系統類型之一[12]。由于其在氣候調節、水源涵養和多樣性維持中發揮著重要作用[13],高寒草地近年來受到眾多學者的關注。其中,多樣性沿環境梯度的空間變化及與群落穩定性之間的關系是植物群落研究的關鍵問題[14-15]。例如,吳紅寶等[16]發現隨著海拔的升高,高寒草地Shannon-Wiener 指數和物種豐富度先增加后降低,而Pielou 均勻度指數先降低后增加;武建雙等[17]研究發現高寒草地植物多樣性隨著生長季累積降水呈現指數增加趨勢;薛晨陽等[18]研究發現群落穩定性與多樣性無顯著關系。這些研究闡明了植物多樣性和群落穩定性隨海拔、降水量等環境梯度的變化趨勢,對理解植物多樣性和群落穩定性有重要意義。但目前關于生物多樣性與群落穩定性的關系尚存在爭議,仍需深入研究[19]。本研究以青藏高原的高寒草甸、高寒濕地草甸、高寒草原和高寒荒漠草原為研究對象,探究不同草地類型植物多樣性與穩定性之間關系及其對土壤因子的響應,揭示高寒草地植物多樣性和群落穩定性關系,為青藏高原高寒草地生態系統可持續發展提供理論依據。
1 材料與方法
1.1 研究區域概況
研究區位于念青唐古拉山和岡底斯山之間,是青藏高原的核心地區,東西橫跨1400 km(79.71°-94.54° E),南北縱跨570 km(28.74°-33.72° N),總面積約45.57 萬km2。該區域氣候屬于典型的高寒大陸性季風氣候,寒冷干燥,年平均降水為88.9~526.1 mm,年平均氣溫為4.1~7.6 ℃,海拔3810~5020 m[20]。研究區域內包含高寒濕地草甸、高寒草甸、高寒草原和高寒荒漠草原4 種草地類型,主要物種為藏北蒿草(Kobresia littledalei),高山蒿草(Kobresia pygmaea),紫花針茅(Stipa purpurea)和沙生針茅(Stipa glareosa)。
1.2 植被調查與樣品采集
于2020年7 月在研究區域內選取高寒濕地草甸、高寒草甸、高寒草原和高寒荒漠草原4 種草地類型各20 個,共計80 個樣點,每兩個樣點之間間隔20 km。在每個樣點選取遠離道路、受人為干擾較少的、地勢相同的草地,設置100 m×100 m 的樣地,每個樣地沿對角線設置6 個1 m×1 m 的觀測樣方,相鄰兩個樣方之間間隔20 m,共480個樣方。在每個樣方內用直徑為7 cm 的土鉆采用5 點法取0~20 cm 土層,充分混合,共采集480 個土樣。研究區域及樣點分布見圖1。

圖1 研究區域及樣點分布Fig.1 Study area and sample point distribution
采用刈割法將植物按照物種齊根割掉,每割掉一個物種均對刈割前后的樣方進行數碼相機拍照,將照片圖像用CAN-EYE-V6313 軟件進行分析,獲取樣方總蓋度和每種植物的分蓋度[21-22]。植株高度使用鋼卷尺測量。割掉的植物按照物種進行分類標號并裝在信封中,放入65 ℃烘箱中烘至恒重,樣方內所有物種的生物量相加即為總生物量。以科為單位將每個樣方內植物分為莎草、禾草、豆科和雜類草4 個功能群,樣方內各科植物的生物量相加即為各功能群的總生物量。pH 采取玻璃電極法測定;土壤有機碳(soil organic carbon,SOC)用重鉻酸鹽法測定;全磷(total phosphorus,TP)含量采用NaOH 熔融-鉬銻抗比色法測定;硝態氮(nitrate nitrogen,NO3--N)用紫外分光光度法測定;速效磷(available phosphorus,AP)和銨態氮(ammonium nitrogen,NH4+-N)用分光光度計法測定[23-25]。
1.3 指標計算
選 取 物 種 豐 富 度 指 數(species richness index,S)、Shannon-Wiener 多 樣 性 指 數(Shannon-Wiener biodiversity index,H)和Pielou 均勻度指數(Pielou evenness index,J)來表征植物多樣性,計算公式如下:
式中:C為樣方內的物種數,Pi為相對重要值。
Godron 穩定性(G)計算方法是目前生態學應用較為廣泛的測定群落穩定性的方法[18,26-27]。植物蓋度能夠表征植物的橫向分布、影響土壤淺層水分[28],是表征高寒草地退化程度的重要指標。因此,本研究采用鄭元潤[29]和王鮮鮮等[30]改進的穩定性計算方法,首先把樣方內植物的蓋度轉換成相對蓋度;其次,將植物的蓋度與種對應,按照蓋度值從大到小排列;然后將群落內相對蓋度與物種總數的倒數按順序分別累加;最后以累積相對蓋度為y,以累積物種數為x,建立一條公式為y=ax2+bx+c 的平滑曲線,繪制出y=100-x直線,兩條線的交點與穩定點(20,80)之間距離越近則表示群落越穩定,即兩點之間歐氏距離與穩定性呈反比。本研究取歐式距離的倒數并乘以100%作為Godron 指數來表征群落穩定狀態。
1.4 數據處理
在國家氣象科學數據中心(https://data.cma.cn/)下載研究區域2001-2020年的國家氣象站數據,在軟件Arcmap 10.2 中,運用普通克里金插值[31]的方法對年均降水量(mean annual precipitation,MAP)和年均溫度(mean annual temperature,MAT)進行插值分析,通過柵格提取獲得采樣點的氣象數據。采用單因素方差和最小顯著差數法(LSD)分析4 種類型草地的植被因子和土壤因子之間的差異以及不同草地類型不同功能群地上生物量和物種數的差異;運用回歸分析揭示不同草地類型群落穩定性與多樣性之間的線性關系;選用結構方程模型分析氣候、土壤、植物多樣性與群落穩定性之間的關系,選用比較擬合指數[compare fitted indices(CFI)>0.9]、漸進殘差均方和平方根[root mean square error of approximation(RMSEA)<0.08]、標準均方根殘差[standardized root mean square residual(SRMR)<0.05]、顯著性概率值(P>0.05)等指標對模型優度進行評價,選出最優模型。最終選用均勻度指數、穩定性指數、MAP、pH、NH4+-N、SOC、TP 構建結構方程。結構方程模型在Amos 21.0 中建立,其他的分析與繪圖均在R 3.6.2 中進行。
2 結果與分析
2.1 不同草地類型植物群落結構特征
不同草地類型植被群落特征存在顯著差異(圖2)。地上生物量和群落總蓋度由高寒濕地草甸、高寒草甸、高寒草原到高寒荒漠草原顯著下降(P<0.05)。高寒荒漠草原物種豐富度指數和Shannon-Wiener 多樣性指數均顯著小于其他3 種草地類型(P<0.05),而其均勻度指數則相反。高寒草原3 種多樣性指標均處在最高水平,但其穩定性指數較低。高寒濕地草甸和高寒草甸3 種多樣性指數均沒有顯著差異。高寒草甸豐富度最高,穩定性顯著高于其他3 種草地類型(P<0.05)。

圖2 不同草地類型植被群落特征、多樣性和穩定性Fig.2 Vegetation community,diversity and stability characteristics of different grassland types不同字母表示不同草地類型間差異顯著(P<0.05),下同。Different letters indicate significant differences among different regions(P<0.05),the same below.
由圖3 可知,4 種草地類型不同功能群地上生物量和物種數差異顯著(P<0.05)。莎草科地上生物量和物種數從高寒濕地草甸、高寒草甸、高寒草原到高寒荒漠草原顯著下降(P<0.05);高寒草原禾本科的地上生物量和物種數以及豆科的物種數均顯著大于高寒濕地草甸和高寒荒漠草原(P<0.05)。高寒草甸雜類草地上生物量和物種數均顯著大于高寒草原和高寒荒漠草原(P<0.05)。由圖4 可知,由高寒濕地草甸和高寒草甸到高寒荒漠草原和高寒草原莎草科相對地上生物量和物種數均減少,禾本科和豆科均增加;雜類草相對地上生物量增加,相對物種數大小排序為高寒草甸>高寒草原>高寒荒漠草原>高寒濕地草甸。綜上所述,由高寒濕地草甸和高寒草甸到高寒草原和高寒荒漠草原,主要功能群由莎草科逐漸轉變為雜類草和禾本科。
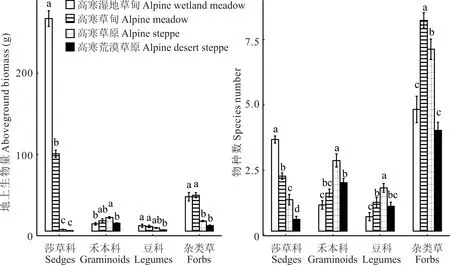
圖3 不同草地植被群落功能群地上生物量和物種數Fig.3 Aboveground biomass and species number of functional communities of different grassland vegetation communities
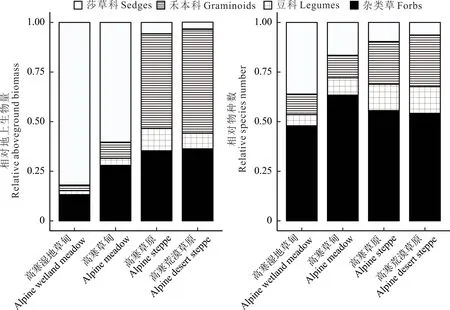
圖4 不同草地植被群落功能群地上生物量和物種數相對比例Fig. 4 Relative proportions of aboveground biomass and species number in functional communities of different grassland vegetation communities
2.2 不同草地類型環境因子變化
由圖5 可知,各環境因子在4 種草地類型間均有顯著差異。MAP 表現為高寒草甸>高寒濕地草甸>高寒草原>高寒荒漠草原(P<0.05),變化范圍為88.95~540.22 mm;pH 在4 種草地類型間顯著下降(P<0.05),變化范圍為5.80~10.00;2 種草甸的NH4+-N、TP 和SOC 均顯著低于2 種草原(P<0.05),其變化范圍分別為5.64~27.54 mg·kg-1、0.24~8.53 g·kg-1和2.27~92.90 g·kg-1;高寒濕地草甸的AP 和NO3--N 均顯著低于2 種草地類型,其變化范圍分別為1.26~10.62 mg·kg-1和1.81~34.08 mg·kg-1。
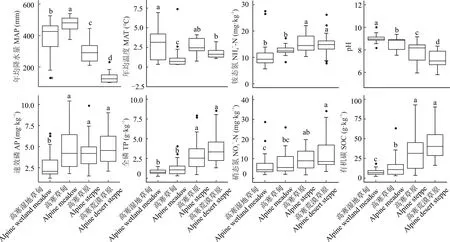
圖5 不同草地類型間環境因子的差異Fig.5 Differences of environmental factors among different grassland types·代表異常值Present abnormal value. MAP:年均降水量Mean annual precipitation;NH4+-N:銨態氮Ammonium nitrogen;TP:全磷Total phosphorus;SOC:土壤有機碳Soil organic carbon.下同The same below.
2.3 高寒草地植物多樣性與群落穩定性的關系及其影響因素
回歸分析結果表明(圖6),群落穩定性與Pielou 均勻度指數在4 種草地類型中均呈現顯著負相關(P<0.05),擬合優度分別為0.900、0.087、0.859 和0.621;在高寒草甸和高寒荒漠草原中與Shannon-Wiener 多樣性指數呈現顯著負相關(P<0.05);在高寒濕地草甸和高寒草原中與物種豐富度指數呈顯著正相關(P<0.05)。綜上所述,不同草地類型植物多樣性與群落穩定性關系不同,但大體表現為提高物種豐富度能夠增強群落的穩定性,提高物種均勻度和多樣性能減弱群落的穩定性。

圖6 不同草地類型植物多樣性與群落穩定性線性回歸分析Fig.6 Linear regression analysis of plant diversity and community stability of different grassland types
高寒草地植物多樣性與群落穩定性的驅動機制如圖7 所示。該模型中,MAP 和NH4+-N 對植物多樣性有顯著的直接影響,其直接效應分別為-0.457、0.129;pH 和SOC 通過NH4+-N 對植物多樣性有顯著間接影響。此外,MAP 和Pielou 指數對群落穩定性有顯著直接影響,其直接效應為0.317 和-0.262;NH4+-N 和MAP 通 過 植物多樣性對群落穩定性有顯著的間接影響。綜上所述,高寒草地植物多樣性的主要影響因子為MAP 和NH4+-N,MAP 和植物多樣性是群落穩定性的主要影響因子。

圖7 高寒草地植物多樣性和群落穩定性驅動因素Fig.7 Driving mechanism of plant diversity and community stability in alpine grassland實線和虛線分別表示正作用和負作用,箭頭的寬度與作用強度呈正比,箭頭上的數字是標準化路徑系數,***,P<0.001;**,P<0.01;*,P<0.05。J:Pielou 均勻度指數Pielou evenness index;G:Godron 穩定性指數Godron stability index.The solid line and the dotted line represent the positive and negative effects respectively. The width of the arrow is proportional to the action intensity. The number on the arrow is the standardized path coefficient.
3 討論
3.1 不同草地類型下植物群落的差異
有研究表明[32],高寒草地地上生物量呈現出高寒草甸>高寒草原>高寒荒漠草原的趨勢。本研究結果與其相符(圖2)。然而本研究中高寒濕地草甸年均降水量小于高寒草甸,地上生物量卻大于高寒草甸。這可能是由于高寒濕地草甸多處在河流湖泊周圍,由于河流湖泊水分的補充,濕地土壤含水量較高,因而高寒濕地草甸植物的可利用水分高于高寒草甸,植被的地上生物量也高于高寒草甸,這說明土壤水分也是影響高寒草地地上生物量的重要的環境因子[28]。
本研究發現,不同草地類型物種豐富度呈現高寒荒漠草原顯著小于其他3 種草地類型(圖2)。這可能是因為不同類型生境的草地中,生境不同的植物所占比例不同導致的[33]。高寒荒漠草原絕大部分為旱生植物,中生植物少于其他3 種草地類型。此外,本研究發現植物的均勻度指數表現為草甸低于草原,這可能是因為土壤養分對物種均勻度的分布有著重要的影響。土壤作為維持陸地生態系統的重要載體,在轉移營養和水分運轉、儲存、調控與外界氣體的交換等方面發揮著重要作用[34-35]。曾思金等[36]研究發現土壤有機碳與Pielou 物種均勻度指數顯著正相關,本研究結果與其相符(圖7)。本研究中,由于高寒草甸和高寒濕地草甸地上生物量較大,植被茂盛,對土壤營養物質的消耗較多,土壤氮素和有機碳的含量較低,這導致了其物種均勻度低于高寒草原和高寒荒漠草原。
本研究中,高寒草甸群落穩定性大于其他3 種草地(圖2),這可能是由于不同草地類型物種的優勢度不一樣導致的。優勢度效應認為優勢種的穩定性對群落穩定性有重要作用[37]。群落組成物種的波動性越小,則群落的方差之和就越少,進而降低了群落的波動性,使群落穩定性更高[38]。研究表明優勢種比其他物種穩定性更高,優勢物種的優勢度增加會使群落更加穩定[39]。此外,本研究發現從高寒濕地草甸到高寒荒漠草原,占優勢地位的功能群從莎草科逐漸變為雜類草和禾本科。這可能與土壤水分有關。水分含量較高時,莎草科占據了大量的生態位,在對光和養分的競爭中處于優勢地位,抑制了雜類草和禾本科植物的生長[40];而水分較少時,耐旱的禾本科和雜類草植株發展壯大。
3.2 植物多樣性與群落穩定性關系
本研究中,不同草地類型植物多樣性與穩定性關系不同,同一草地類型不同多樣性指數對群落穩定性影響也不同,總體表現出均勻度的提高降低了群落穩定性(圖6),這可能與群落抗干擾能力有關。Hillebrand 等[41]研究表明均勻度的提高會提高群落的恢復力,降低抗干擾能力。而本研究的樣地處于生態環境脆弱、對外界刺激響應敏感的高寒地區,群落演替時間較長,一旦被破壞短時間內難以恢復,群落的抗干擾能力對穩定性的影響更大,因而均勻度與穩定性呈現負相關關系。另外,本研究發現物種豐富度指數與群落穩定性呈正相關關系,這與胡冬等[19]的研究結果一致。由于受到物種間相互作用對環境波動的放大效應和物種對環境波動的敏感性的影響,生物多樣性能夠增加物種對環境波動響應的多樣性,從而使群落的穩定性增加。多樣性指數是把物種豐富度與均勻度結合起來的統計量,多樣性信息度量、概率度量和幾何度量都是以不同的方式表達物種豐富度與均勻度的結合[42]。本研究中Shannon-Wiener 多樣性與群落穩定性呈負相關關系,這可能是因為高寒草甸中均勻度與群落穩定性的擬合程度更好,對穩定性的影響高于物種豐富度。
3.3 環境因子對植物多樣性和群落穩定性的影響
降水量能夠在不同程度上影響植物群落結構、生物多樣性和穩定性。高寒草地生態系統對氣候變化敏感,尤其是降水的變化會引起植物水分可利用性在時間空間上的差異,導致植物種內、種間關系的變化[43],進而引起群落穩定性的變化。本研究發現降水能夠通過直接和間接作用促進高寒草地植物多樣性和群落穩定性提高,其原因可能是降水能夠促進土壤中種子的萌發、加速土壤有效養分的溶解和流動[44]、提高植物對養分的吸收效率[45],提高物種多樣性從而提高群落的穩定性。土壤養分對于植物多樣性起著至關重要的作用[11],本研究發現土壤中的NH4
+-N 對植物多樣性和群落穩定性有顯著影響。這是由于高寒草地氮素含量較高,導致其他營養元素的相對不足,抑制植物豐富度的增加[44],進而降低了群落的穩定性。另外,有研究表明SOC 和pH 對植物多樣性有直接影響[46],而本研究發現SOC 和pH 通過影響NH4+-N 間接影響植物多樣性。這可能是因為本研究地域跨度大,各草地類型之間植物組成不同,對所處生境資源的需求、能力、方式等均有差異,導致環境因子與植物多樣性并未產生直接效應[47]。
4 結論
高寒草地不同草地類型植被群落特征差異顯著;高寒草原植物多樣性顯著高于其他3 種草地類型,高寒草甸群落穩定性最高;植物多樣性和群落穩定性的關系并不相同,但大體表現為提高物種豐富度能夠增強群落的穩定性,提高物種均勻度和多樣性能減弱群落的穩定性。降水量、土壤有機碳、pH 以及銨態氮含量是影響高寒草地植物多樣性和群落穩定性的主要因素。在高寒草地生態系統的管理過程中,應關注氣候和土壤因子變化,根據不同草地類型制定不同的管理措施。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
