天津海域海月水母分布特征及其與環境因子的關系
2023-04-06 06:43:52張博倫鄭德斌徐曉甫曾祥茜
天津農業科學 2023年3期
郭 彪,王 碩,張博倫,鄭德斌,徐曉甫,曾祥茜
(1.天津市水產研究所,天津,300457;2.天津市海洋牧場技術工程中心,天津,300457)
海月水母(Aurelia aurita)是全球廣布種,隸屬于缽水母綱(Scyphozoa)、旗口水母目(Semaeoslomeae)、洋須水母科(Ulmaridae)、海月水母屬(Aurelia),廣泛分布在70°N~40°S 的沿岸海域[1]。近年來,海月水母在我國渤海沿岸頻有暴發[2-4]。海月水母作為東亞海域常見的致災水母種類[5-7],其暴發嚴重影響海洋生態系統和社會經濟發展[4]。一方面,水母攝食浮游動物、魚卵仔稚魚,可以改變浮游動物群落結構及生態系統營養結構,水母暴發會影響多個營養級,下行控制初級生產力[8-10]。另一方面,水母暴發會直接破壞漁業捕撈網具[11]、污染養殖設施[12]、危害增養殖生物[13]、造成基礎工業設施管道堵塞[14]等。
海月水母生活史復雜,具有有性世代和無性世代的世代交替[2]。性成熟海月水母通過有性生殖釋放浮浪幼蟲,經一段浮游生活后進行附著,形成螅狀體[2,15]。螅狀體能夠通過出芽、足囊、匍匐莖等多種無性繁殖方式進行螅狀體的擴充[2,15]。在適宜的環境條件下,螅狀體通過橫裂釋放營浮游生活的碟狀體[2,15]。碟狀體在適宜的溫度、餌料供給條件下迅速生長發育為性成熟海月水母[2,15]。
天津位于渤海西岸,其近岸海域營養鹽豐富,歷史上是魚蝦的重要索餌場[16]。近年來,天津海域海月水母持續暴發,漁民生產網具破壞嚴重,漁民無法進行正常生產作業,許多漁船不得不放棄秋季對蝦捕撈生產作業,2021 年天津對蝦海捕統計產量為零[17]。目前,關于天津海域海月水母的分布特征未見報道,雖然王朋鵬等[3]于2018 年6 月份調查了整個渤海水母的分布特征,但缺少天津海域的調查站位。
全面了解海月水母在天津海域的分布特征,一方面有助于人們了解天津海域海月水母的暴發源頭及其變動規律;另一方面有助于人們制定海蜇等生物的增殖策略,避免海月水母的危害[13],盡可能提高海蜇的增殖效益。為此,本研究在2021 年1—12 月期間系統性調查了天津海域海月水母分布特征,分析海月水母種群分布特征與環境因子的關系,以期為水母防控和海蜇增殖提供科學參考。
1 材料與方法
1.1 調查時間和地點
依據海月水母的生活習性[2,15],2021 年1 月下旬從不同區域(北疆電廠、中心漁港航道、北塘、東疆、臨港、南港)海岸線開始向遠離海岸線方向推進調查,直至無海月水母碟狀體為止;同時在海洋牧場人工魚礁區設計3 個站位進行調查。2021 年2—12月,每個月下旬安排1 個航次調查海月水母分布特征。每個調查依航次據上個航次調查結果逐步擴大調查范圍。1—2 月調查站位圖見圖1,其他月份調查站位參考結果部分圖2 和圖3。
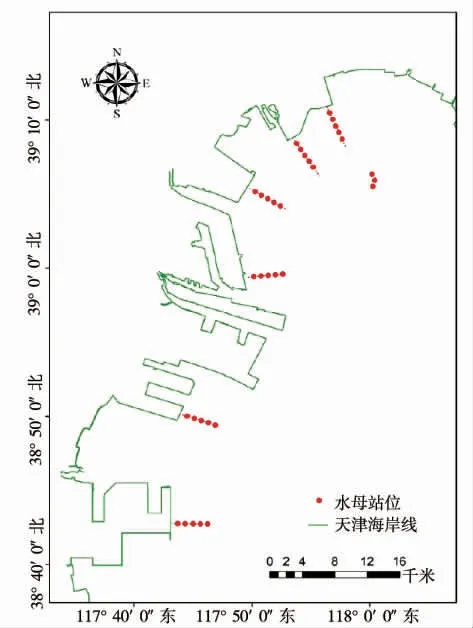
圖1 初始調查站位示意圖
1.2 調查方法
(1)海月水母蝶狀體調查:調查采用淺水Ⅰ型網由底到表垂直拖網采集,利用4%(V/V)福爾馬林海水固定保存,帶回實驗室在解剖鏡下進行全樣品鑒定并計數。2021 年1—4 月調查航次采用該方法。
(2)海月水母成體調查:采用大型浮游生物網在海水表層進行水平拖拽,拖拽船速為2 km·h-1,拖拽時間為10 min,如果拖拽過程中發現浮游生物網網兜內海月水母已滿,網兜有嚴重下沉情況需立即起網,并記錄好拖拽時間。2021 年5—12 月調查航次采用該方法。
(3)環境因子調查:水溫、鹽度、溶解氧、pH 等水質指標采用美國YSI 多參數水質分析儀(型號YSI professional plus)測定;葉綠素a 按照《海洋監測規范》(GB 17378—2007)進行樣品采集與測定;同時采用船載LOWRANCE HDS-7 聲吶探測儀測定各站位水深。
1.3 數據處理
蝶狀體密度按體積計算,其計算公式如下:
式中,Dd 為蝶狀體密度;Nd為樣品中蝶狀體的總個數;V 為垂直拖網水體體積。
由于海月水母成體基本上均漂浮在海水表面,故資源密度采用掃海面積法進行計算。其計算公式如下:
式中,Dc為海月水母成體密度;Nc為采集到的海月水母總數量;S 為采樣過程中掃海的總面積。
所獲的不同區域和不同時間海月水母資源量數據采用SPSS19.0 進行單因素方差分析,分析結果采用SigmaPlot 軟件進行制圖,不同站位的資源密度采用ArcGIS 軟件作圖。運用SPSS 19.0 軟件計算了海月水母豐度與海水溫度、鹽度、溶解氧含量、pH 和葉綠素a 之間的Pearson 相關性系數,并用Origin 2018 軟件繪制圖形。
2 結果與分析
2.1 天津近岸海域海月水母蝶狀體空間分布特征
2021 年1—4 月在天津近岸海域完成4 次海月水母蝶狀體調查,其中1 月和2 月未發現海月水母碟狀體,3 月和4 月發現海月水母蝶狀體,其空間分布特征見圖2。
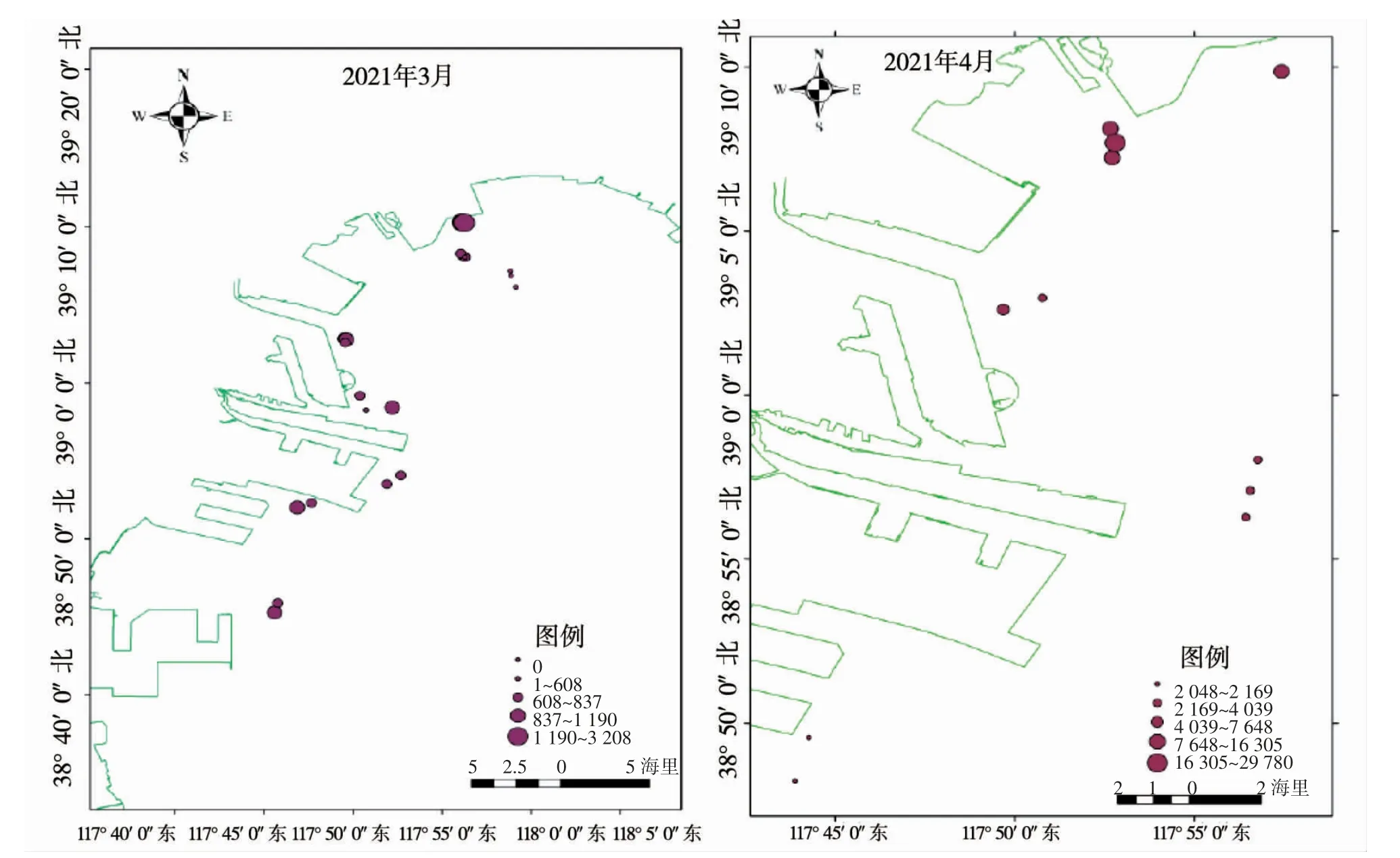
圖2 海月水母蝶狀體分布示意圖
海月水母蝶狀體在天津近岸海域出現于3月底,主要分布在近岸線50 m 附近的海域,且岸線須有可供海月水母水螅體附著的礁石或人工岸線;海月水母碟狀體的平均傘徑長為(4.56±0.63)mm,蝶狀體分布海域平均分布密度為(1 063.46±692.14)ind·m-3,電廠附近蝶狀體密度最高,遠高于其它調查海域(P<0.05)。在遠離岸線的海洋牧場人工魚礁區內未發現海月水母碟狀體。
2021 年4 月底,海月水母蝶狀體呈數量級增加,分布海域平均分布密度高達(9 165.89±8 758.09)ind·m-3,平均傘經長變化不明顯,為(5.56±1.61)mm。蝶狀體也由近岸線50 m 附近逐步擴大到近岸線5.556 km 范圍。中心漁港附近海域海月水母碟狀體的密度顯著高于其他調查海域(P<0.05),其他調查海域蝶狀體密度沒有顯著性差異(P>0.05)。
2.2 天津近岸海域海月水母成體空間分布特征
2021 年5—12 月在天津近岸海域完成8 個航次的海月水母成體調查,其中10—12 月調查未發現海月水母成體,其他月份海月水母成體空間分布特征見圖3。
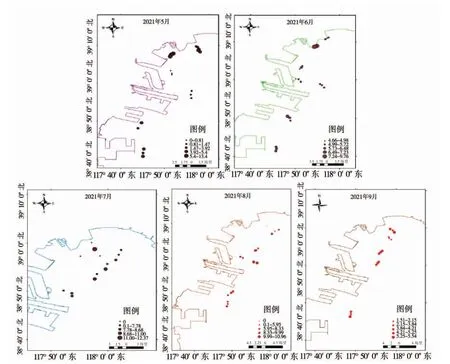
圖3 海月水母成體分布示意圖
2021 年5 月,海月水母繼續擴散分布,平均傘徑為(2.56±1.67)cm,表層水體平均分布密度為(4.48±3.81)ind·m-3。不同海域海月水母密度差異明顯,其中中心漁港附近海域的密度顯著高于其他調查海域(P<0.05),北塘和東疆附近海域的密度顯著低于其他調查海域(P<0.05)。
2021 年6 月,海月水母在全天津海域均有分布,平均傘徑為(8.23±3.18)cm,表層水體平均分布密度為(6.20±1.59)ind·m-3。不同海域海月水母密度差異正在逐步縮小,但中心漁港附近海域的密度仍然顯著高于其他調查海域(P<0.05)。
2021 年7 月,受強降雨和各河口排放淡水的影響,天津沿岸海域海水呈明顯褐黑色,水體中有大量死亡的藻類。在褐黑色水域中未發現海月水母分布。在遠離岸線海域(海水顏色正常)發現海月水母,平均傘徑為(13.23±6.11)cm,其表層水體平均分布密度為(8.63±1.75)ind·m-3。各調查海域海月水母的分布密度沒有顯著性差異(P>0.05)。
2021 年8 月,天津沿岸海域仍存在少量褐黑色海水分布區域,在此區域未發現海月水母。其他區域調查發現海月水母分布均勻,各區域海月水母密度沒有顯著性差異(P>0.05)。海月水母平均傘徑為(14.41±7.77)cm,其表層水體平均分布密度為(8.63±1.36)ind·m-3。
2021 年9 月,較大個體的海月水母消失,海月水母平均傘徑為(5.03±2.71)cm。海月水母在天津海域分布均勻,各區域海月水母密度沒有顯著性差異(P>0.05),其表層水體平均分布密度為(4.26±1.38)ind·m-3。
2.3 天津近岸海域海月水母的時間分布特征
海月水母蝶狀體密度在時間序列上波動較大,2021 年3 月底開始出現,4 月底顯著增加,4 月份海月水母蝶狀體密度顯著高于3 月份(P<0.05)(圖4)。
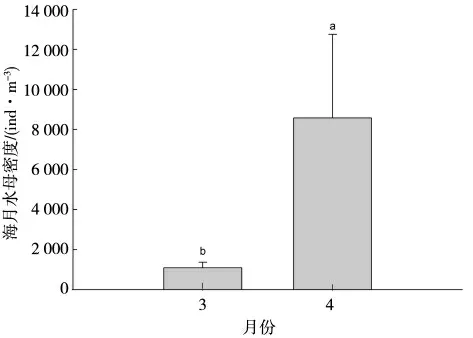
圖4 海月水母蝶狀體豐度變化
將2021 年5—9 月海月水母成體資源密度進單因素方差分析,結果顯示7 月和8 月海月水母成體資源密度均值顯著高于5 月和9 月(P<0.05)。海月水母成體資源密度的均值在5 月、6 月和9 月之間無顯著性差異(P>0.05),6 月、7 月和8 月間無顯著性差異(P>0.05)(圖5)。

圖5 海月水母成體豐度變化
2021 年3—9 月采集到海月水母傘徑均值和最大值見圖6。由圖6 可以看出,2021 年3—9 月海月水母傘徑的均值和最大值均隨著時間的推移有所增加。進入9 月份,由于大規格海月水母的消失,漁獲水母傘徑的均值和最大值均出現明顯下降。
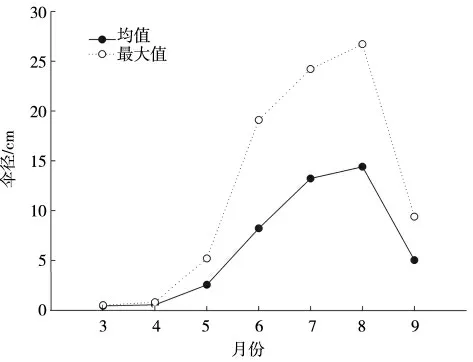
圖6 海月水母傘徑變化
2.4 海月水母分布與環境因子的關系
不同調查站位2021 年3 月和4 月海月水母蝶狀體密度與環境因子之間的關系如圖7 所示。海月水母蝶狀體密度與溫度、鹽度均具有顯著相關性(P<0.05),而與pH、溶解氧和葉綠素a 相關性不顯著(P>0.05)。其中,蝶狀體密度與溫度呈顯著正相關,相關性系數0.744;蝶狀體密度與鹽度呈顯著負相關,其相關性系數-0.693。
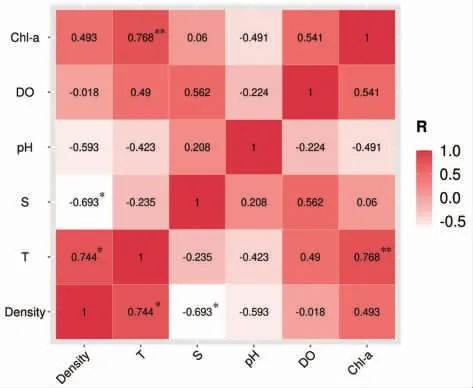
圖7 2021 年3—4 月海月水母蝶狀體分布密度與環境因子(溫度、鹽度、pH、溶解氧和葉綠素a)相關性分析結果
不同調查站位2021 年5—9 月海月水母成體密度與環境因子間的關系如圖8 所示。海月水母成體體密度與溫度呈極顯著正相關(P<0.01),其相關性系數為0.595。海月水母成體體密度與葉綠素a 同樣呈極顯著正相關(P<0.01),其相關性系數為0.506。而海月水母成體體密度與鹽度、pH 和溶解氧均沒有顯著相關性(P>0.05)。
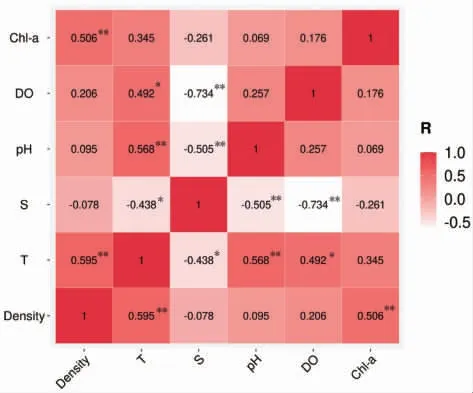
圖8 2021 年5—9 月海月水母成體分布密度與環境因子(溫度、鹽度、pH、溶解氧和葉綠素a)相關性分析結果
3 討論與結論
3.1 討論
在自然海域,當環境條件適宜時海月水母螅狀體發生橫裂釋放蝶狀體[18],因此蝶狀體可用于指示海域是否存在螅狀體[4,19]。本研究中,海月水母蝶狀體最初的出現海域為近岸50 m 有礁石或人工岸線的海域,其中中心漁港附近海域蝶狀體數量較高。這表明,天津近岸海域海月水母螅狀體棲息的主要海域為近岸有礁石或人工岸線的海域,其中中心漁港海域為主要棲息地。
2021 年3 月電廠附近海域蝶狀體數量最高,其原因可能與該海域平均水深較淺有關。據調查發現,電廠海域的平均水深僅有不足2 m,遠低于其他調查海域。較淺水深海域受太陽輻射等影響較大,其水溫日變化幅度較大[20],而溫度變化,尤其是較高的水溫可能誘導海月水母螅狀體橫裂生殖產生蝶狀體[21-22]。
海洋牧場海域存在大量人工魚礁,理論上其可以為海月水母螅狀體提供附著基[23]。但是,筆者的研究中并未發現海月水母蝶狀體出現在海洋牧場人工魚礁區。海洋牧場海域存在大量腹足綱生物[24],這些生物可能會大量攝食海月水母螅狀體,抑制了該海域海月水母螅狀體的棲息[15,25],需要進一步深入研究。
溫度通過調節海月水母機體的酶活性等,實現對海月水母碟狀體和水母體生長及代謝的影響[17]。相對溫暖水溫會刺激海月水母螅狀體發生橫裂生殖[22],10~17 ℃是膠州灣海月水母產生蝶狀體的溫度[26]。本研究中,海月水母蝶狀體的出現基本符合上述規律,3 月底當水溫接近于10 ℃時,海月水母蝶狀體開展在近岸海域出現,4 月底當水溫升至14 ℃左右時,海月水母蝶狀體大量出現。對海月水母蝶狀體密度和溫度相關性分析也顯示,在3 月和4 月期間,海月水母蝶狀體密度與溫度呈顯著正相關,這一結果與其他人的研究結果基本一致[4,21]。本研究中,海月水母成體密度和溫度呈極顯著正相關,這種相關性與海月水母成體的季節變化趨勢相吻合。Lucas等[27]研究發現,海月水母幼體一般在早春時期緩慢生長,春末夏初隨著水溫的升高呈指數增長,夏末種群數量和個體大小達到最大并停止生長,秋季隨著性成熟和浮浪幼蟲釋放、胃絲排空、生長衰退,個體萎縮變小,隨后死亡。本研究中,海月水母密度和傘徑的季節性變化規律與其完全一致。
鹽度是影響海月水母無性生殖過程的環境因子之一[28]。海月水母螅狀體發生橫裂生殖產生蝶狀體的數量隨鹽度降低(34~27)而增加[12]。本研究中,雖然3 月和4 月不同區域之間海水鹽度變化不大,但是海月水母蝶狀體密度仍展現出與鹽度顯著負相關。這說明水體鹽度與蝶狀體豐度的相關性很強,薛力園等[4]在研究秦皇島海域海月水母蝶狀體分布與環境因子關系時也發現與本研究相一致的結論。
葉綠素a 能夠反應水體中浮游植物的生物量[29],而海月水母的數量會隨著浮游植物生物量的升高而增加[27],這可能與浮游植物生物量增加,提高了海月水母食物的可利用率有關[30]。本研究中,海月水母成體密度與葉綠素a 呈極顯著正相關,說明浮游植物的生物量影響海月水母的分布。
3.2 結論
綜合上述研究結果,總結如下結論和建議:(1)天津近岸海月水母蝶狀體的來源地主要是以中心漁港為主的近岸有礁石和人工岸線海域,為本地生長型,其在每年4 月底會集中暴發。(2)海月水母成體種群數量和個體大小從5 月份開始逐步增大,到8月份達到峰值,隨后二者開始下降,進入10 月底基本消失。(3)天津近岸海月水母蝶狀體與海水溫度和鹽度顯著相關,成體與海水溫度和葉綠素a 極顯著相關。(4)結合天津近岸海月水母的分布特征,建議海蜇增殖放流盡量在5 月初進行,放流地點應選擇距海岸線11.112 km 以上海域,以降低海蜇苗種被海月水母捕食的機率,提高海蜇的存活率。
