我國棱果花天然主要分布區葉片多樣性分析
2023-04-11 03:34:30徐斌易禮江林子湛陳新宇廖煥琴潘文楊會肖
熱帶亞熱帶植物學報 2023年2期
徐斌, 易禮江, 林子湛, 陳新宇, 廖煥琴, 潘文, 楊會肖*
我國棱果花天然主要分布區葉片多樣性分析
徐斌1, 易禮江2, 林子湛3, 陳新宇1, 廖煥琴1, 潘文1, 楊會肖1*
(1. 廣東省森林培育與保護利用重點實驗室/廣東省林業科學研究院,廣州 510520;2. 廣東省沙頭角林場, 廣東 深圳 518081;3. 廣東陽春鵝凰嶂省級自然保護區管理處, 廣東 陽春 529631)
為了解棱果花()種質資源葉片表型的多樣性,對棱果花6個種群的139株的葉片性狀進行方差分析、相關性分析、主成分分析及聚類分析。結果表明,棱果花種群間和個體間的葉片表型性狀存在顯著差異,變異系數為0.14~0.84,平均為0.32,說明棱果花葉片性狀變異較大。葉面積、葉周長與葉長間的相關系數為0.75~0.99。葉片性狀可提取為5個主成分,累計貢獻率達88%,其中葉片的面積、周長、寬度、長/寬和葉形是表型變異的主要因素。聚類分析將139株棱果花分為6組,第Ⅰ組包含34株材料,第Ⅱ組7株,第Ⅲ組1株,第Ⅳ組15株,第Ⅴ組37株和第Ⅵ組45株,說明來自不同種群的棱果花植株分散在不同的組中,棱果花葉片表型與來源無顯著關系。因此,棱果花葉片表型多樣性豐富,其中種群間和單株間具有豐富的多樣性,可為棱果花遺傳改良和新品種選育提供基礎。
棱果花;種質資源;葉片性狀;相關性分析
種質資源群體多樣性是長期進化的產物,群體多樣性越高,表明對環境變化的適應能力越強[1]。植物通過葉片將光能轉換成化學能,因此葉片是植物物質積累的重要場所,是決定植物生物量大小的重要器官之一。不同植物的葉片在葉形、顏色、質地、葉緣等存在明顯的差異,而同種植物不同種質資源的葉片表型性狀也存在較大的差異[2–4]。通過對植物葉片性狀變異規律進行研究,可為優質種質篩選提供依據。
棱果花()為野牡丹科(Melasto- mataceae)棱果花屬灌木,單屬單種,聚傘狀花序, 花瓣長圓狀、橢圓形或近倒卵形,花白色至粉紅色或紫紅色,分布于我國廣東、湖南、廣西、福建、臺灣等地。棱果花花期為10月到翌年4月,比野牡丹科其他多數植物花期(多為6—9月)較晚,花期長、抗逆性強,適生范圍廣。目前已對廣東、海南、福建和湖南等地區野牡丹科觀賞植物進行了資源調查[5–9],并對其觀賞性及園林應用價值進行了評價分析[10–12],但對棱果花種質資源的表型性狀多樣性研究未見報道。本研究利用139株棱果花對種質資源進行葉片表型性狀分析,結合相關分析、主成分分析和聚類分析研究棱果花種質資源的多樣性,為棱果花種質資源的改良利用、新種質的創制及雜交親本選擇提供依據。
1 材料和方法
1.1 材料
根據現存棱果花天然資源分布,在分布相對集中的6個天然種群,選取間距在30 m以上的植株,于2021年春季采集樹冠外側東、南、西、北4個方位的成熟葉片,各種群的來源和編號見表1。

表1 棱果花資源
1.2 葉片表型性狀測定
從東、南、西、北4個方位選取植株中部外側成熟葉片各2片,將葉子平展后用掃描儀掃描,采用萬深LA-S植物圖像分析儀(TMA1600)檢測葉面積(mm2)、葉柄面積(mm2)、葉周長(mm)、葉片長(mm)、葉柄長(mm)、葉片寬(mm)、葉柄寬(mm)、葉柄長/寬、葉長/寬、葉綠素和葉形數據。用游標卡尺測量葉片厚度;用電子天平先稱取葉片鮮質量, 然后將葉片置于烘箱中烘干,再用電子天平稱取葉片干質量,數據取平均值。
1.3 數據分析
利用Excel對原始數據進行整理,數據的基本統計分析則采用R軟件[13]。用“psych”和“cluster”包分別進行主成分分析和聚類分析,其中聚類分析基于系統聚類中離差平方和進行聚類;用ASReml軟件分析棱果花葉片形狀的性狀變異[14],混合線性模型為Y=u+P+I+e,其中是第種群第單株的觀測值,為總葉片平均值,P為第個種群的隨機效應值,I為第個種群內第個單株的隨機效應值,e為隨機誤差。
2 結果和分析
2.1 葉片表型性狀變異分析
對139株棱果花葉片表型性狀的變異分析表明,葉面積、葉柄面積分別為2 000.00和19.00 mm2; 葉周長、葉長、葉柄長、葉寬、葉柄寬分別為220.00、82.00、11.00、39.00和2.50 mm;葉柄長/寬、葉長/寬、葉綠素、葉形分別為4.90、2.20、4.50、2.00; 葉厚、葉鮮質量和葉干質量分別為0.31 mm、9.10 g和6.46 g。葉片表型性狀的變異系數為14%~84%, 說明棱果花種質資源的葉片表型性狀差異較大, 性狀表現不穩定。其中變異系數較大的是葉柄面積和葉干質量,變異系數分別為61%和84%。除葉形的峰度系數為5.80外,其余性狀的偏度系數和峰度系數均約為1,說明這些葉形性狀符合近似正態分布,可用于相關性、主成分分析和聚類分析(表2)。
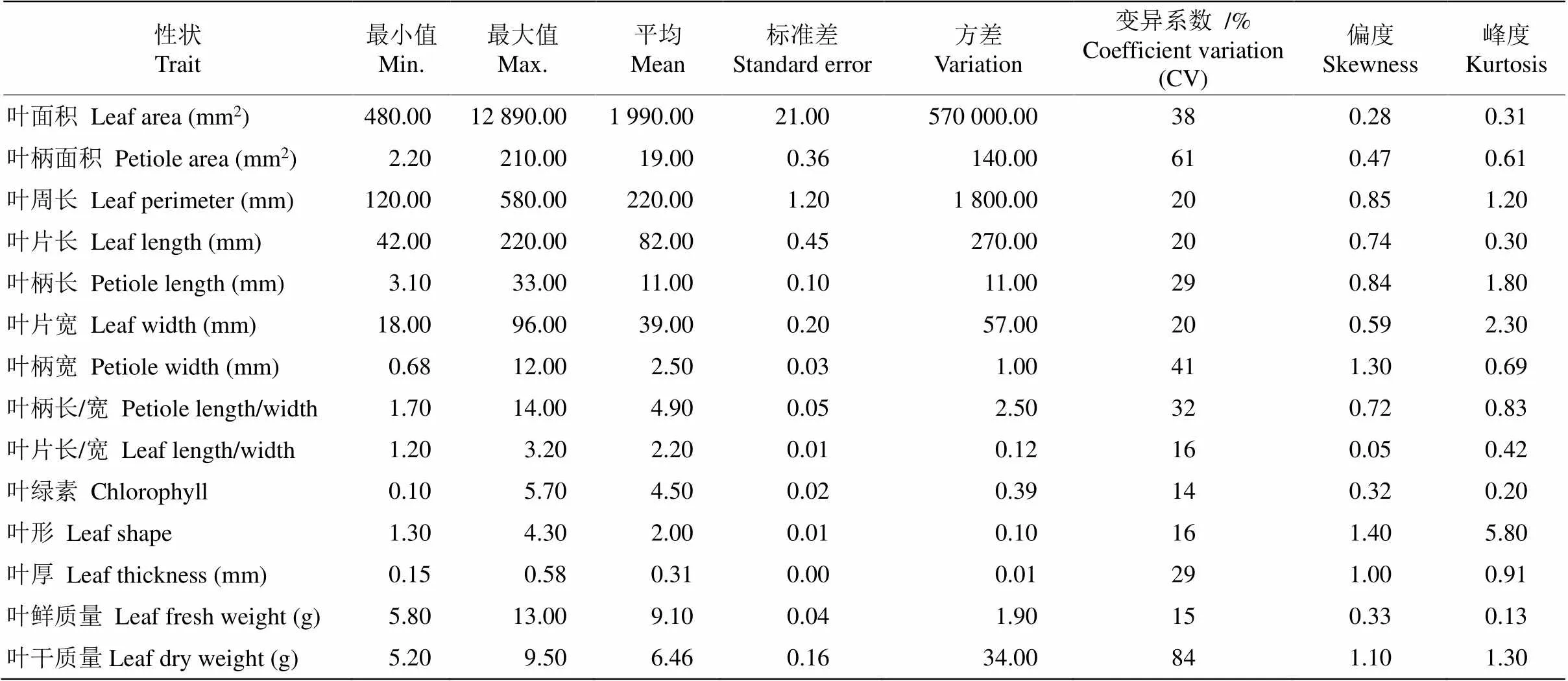
表2 棱果花葉片表型性狀的變異
對139株棱果花葉片表型性狀進行方差分析表明,不同種群棱果花的葉面積、葉柄面積、葉周長、葉長、葉柄長、葉寬、葉柄寬、葉柄長/寬、葉長/寬、葉綠素、葉形、葉厚度、葉鮮質量和葉干質量的差異達極顯著水平(表3,<0.05)。其中,葉綠素在種群間的值最大,為131.53,葉寬在種群內的值最大,為9.18,說明139株棱果花的葉片性狀在種群間和個體間存在豐富的表型變異,為棱果花進一步評價研究提供材料。
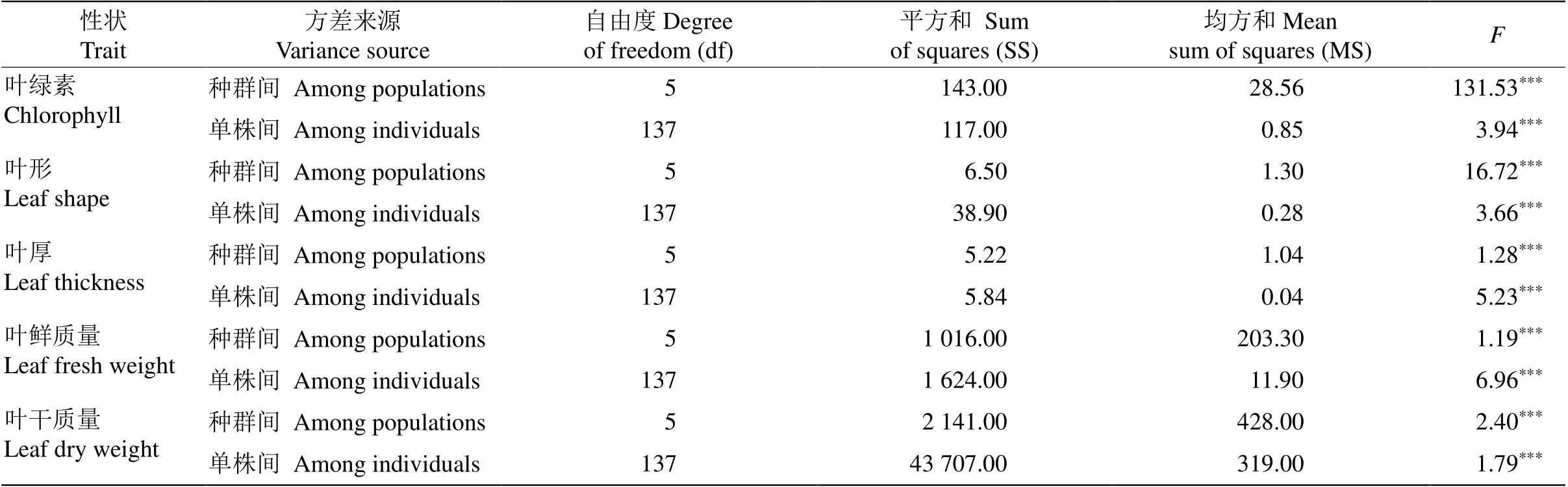
續表(Continued)
***:<0.001
2.2 葉片表型性狀相關分析
棱果花葉片表型性狀的相關分析結果表明(表4),除葉綠素和葉厚度外,其他葉片表型性狀間的相關系數較高,其中葉面積、葉周長、葉長和葉寬間存在極顯著正相關,相關系數為0.660~0.997。葉鮮質量與葉面積、葉周長、葉長和葉寬存在極顯著正相關,相關系數為0.360~0.600。葉形與葉長/寬呈極顯著正相關,相關系數為0.682。葉厚度與葉長/寬呈極顯著負相關,相關系數為0.369。
2.3 葉片表型性狀的主成分分析
植物不同性狀間存在一定的關聯,而單個性狀的作用較難分析,通過主成分分析對多個性狀原有信息進行抽提和合并,進而簡化成數量較少的獨立性狀。通過對棱果花14個葉片表型性狀進行主成分分析,結果見表5。以特征值為1進行抽提,共獲得5個主成分,累計貢獻率為88%,說明139株樣品的14個葉片性狀的大部分信息可以用這5個主成分提取。
第1主成分對葉片表型變異的貢獻率最大,為46%,包含葉面積、葉寬、葉周長、葉長、葉鮮質量、葉柄面積、葉柄寬、葉柄長、葉厚、葉綠素、葉干質量、葉長/寬、葉柄長/寬和葉形,說明第1主成分是表示葉面積的綜合因子。第2主成分的貢獻率為20%,主要為葉長/寬、葉柄長/寬和葉形, 說明第2主成分是表示葉形的綜合因子。第3主成分的貢獻率為10%,貢獻較大的性狀為葉綠素、葉柄面積和葉柄長,說明第3主成分是表示葉綠素值和葉柄性狀的綜合因子。第4個主成分的貢獻率為6%,貢獻較大的性狀為葉干質量,說明第4主成分是表示葉干質量的綜合因子。第5個主成分的貢獻率為5%,貢獻較大的性狀為葉厚度,說明第5主成分是表示葉厚度的綜合因子。因此,棱果花葉片表型的差異主要表現在葉面積、葉周長和葉寬上。
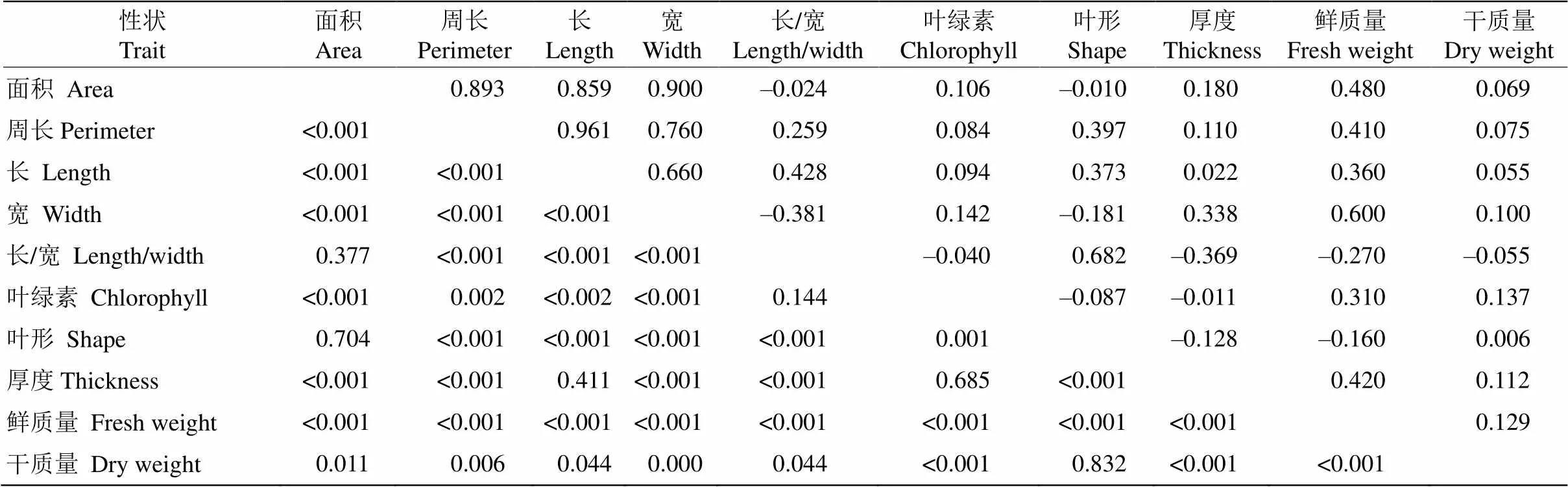
表4 棱果花葉片表型性狀相關系數(右上)和顯著性檢驗(左下)
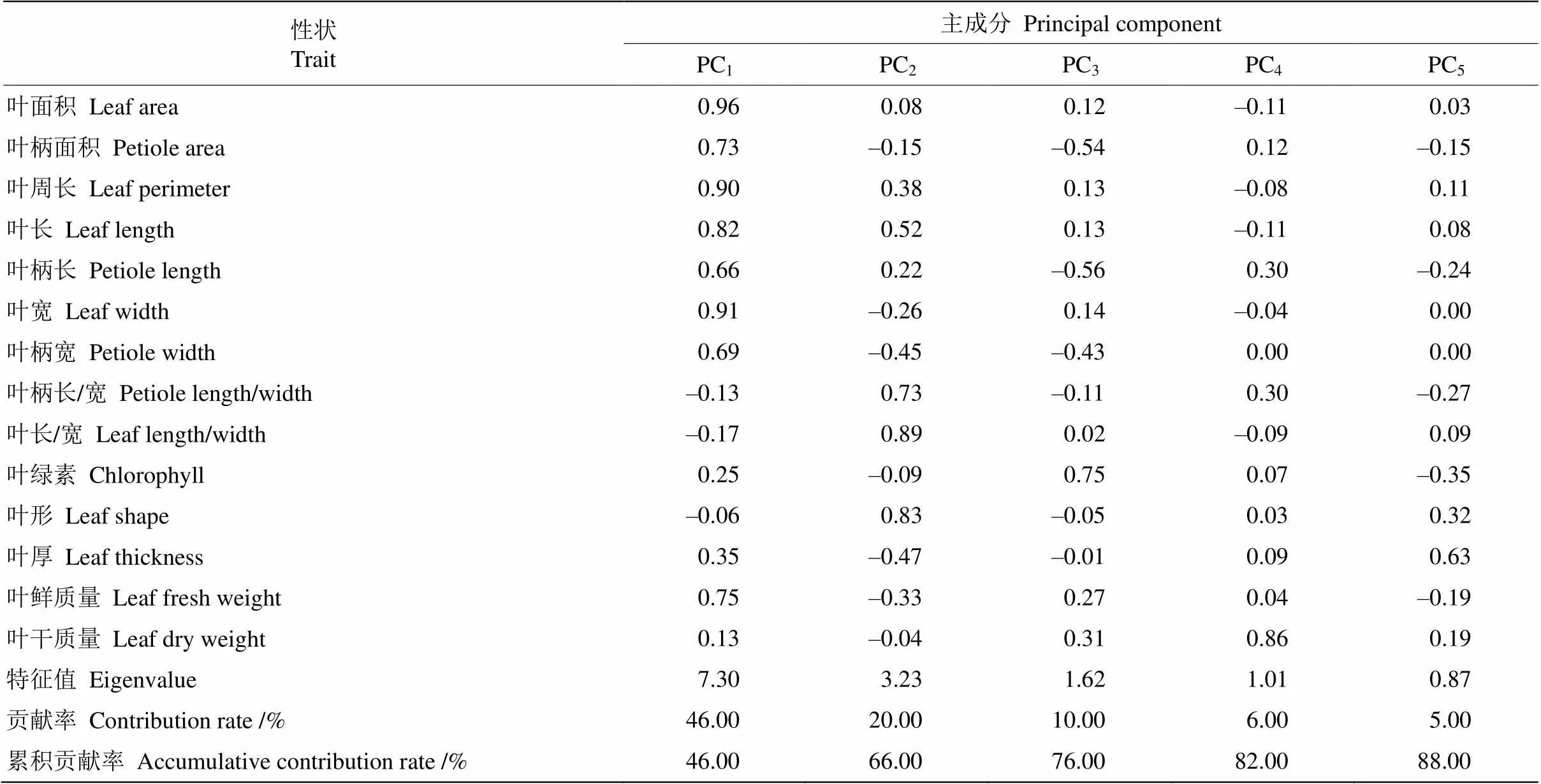
表5 棱果花葉片表型性狀的主成分分析
2.4 葉片表型性狀的聚類分析
利用R軟件中的“psych”包對139株棱果花的14個葉表型性狀進行聚類分析,并構建相關聚類圖(圖1)。由圖1和表6可見,當距離=1時,139株棱果花可劃分為6組,第Ⅰ組包含34株,占樣本總數的24.46%,葉柄長和葉柄寬最大,分別為12.12、2.65 mm;第Ⅱ組包含7株,約占樣本總數的5.04%,葉面積、葉柄面積、葉周長、葉長、葉柄長較大,分別為3274.8、3254.65 mm2和305.2、279.89、106.44、14.19 mm;第Ⅲ組包含1株,約占0.72%,葉面積、葉柄面積、葉周長和葉寬最小,分別為681.38、678.70 mm2和133.04、20.82 mm;第Ⅳ組含15株,約占10.79%,葉柄長和寬較大,分別為12.94、2.98 mm;第Ⅴ組包含37株,約占26.62%;第Ⅵ組包含45株,約占32.37%,葉綠素較小,為4.28。這表明來自不同種群的樣品分散在不同的組中,說明葉片表型聚類與樣品來源無顯著關系,進一步說明了棱果花種群內部具有豐富的多樣性,為開展棱果花種內雜交育種的親本選擇奠定基礎。
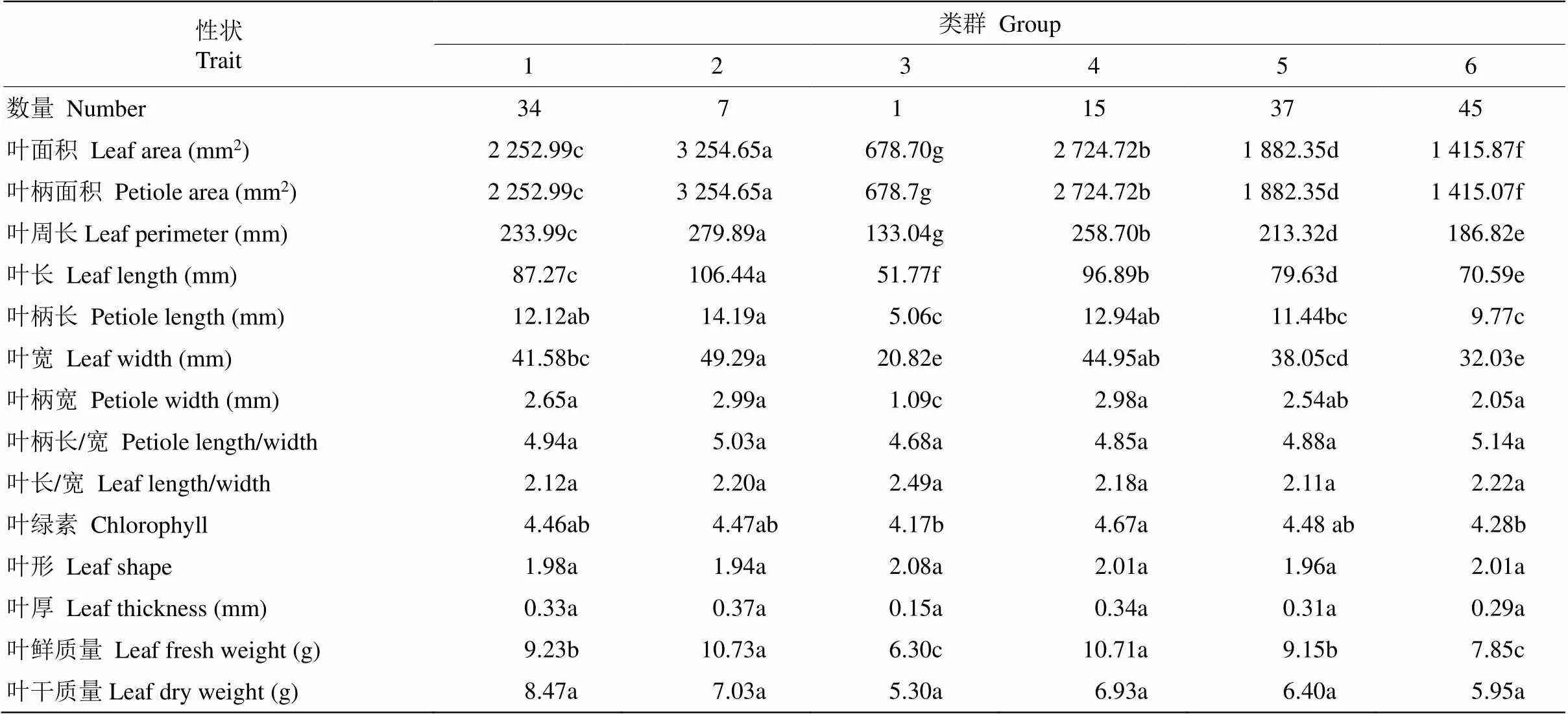
表6 棱果花種質資源各類群葉片性狀特征
同行數據后不同字母表示差異顯著(<0.05)。
Data followed different letters at the same line indicate significant difference at 0.05 level.
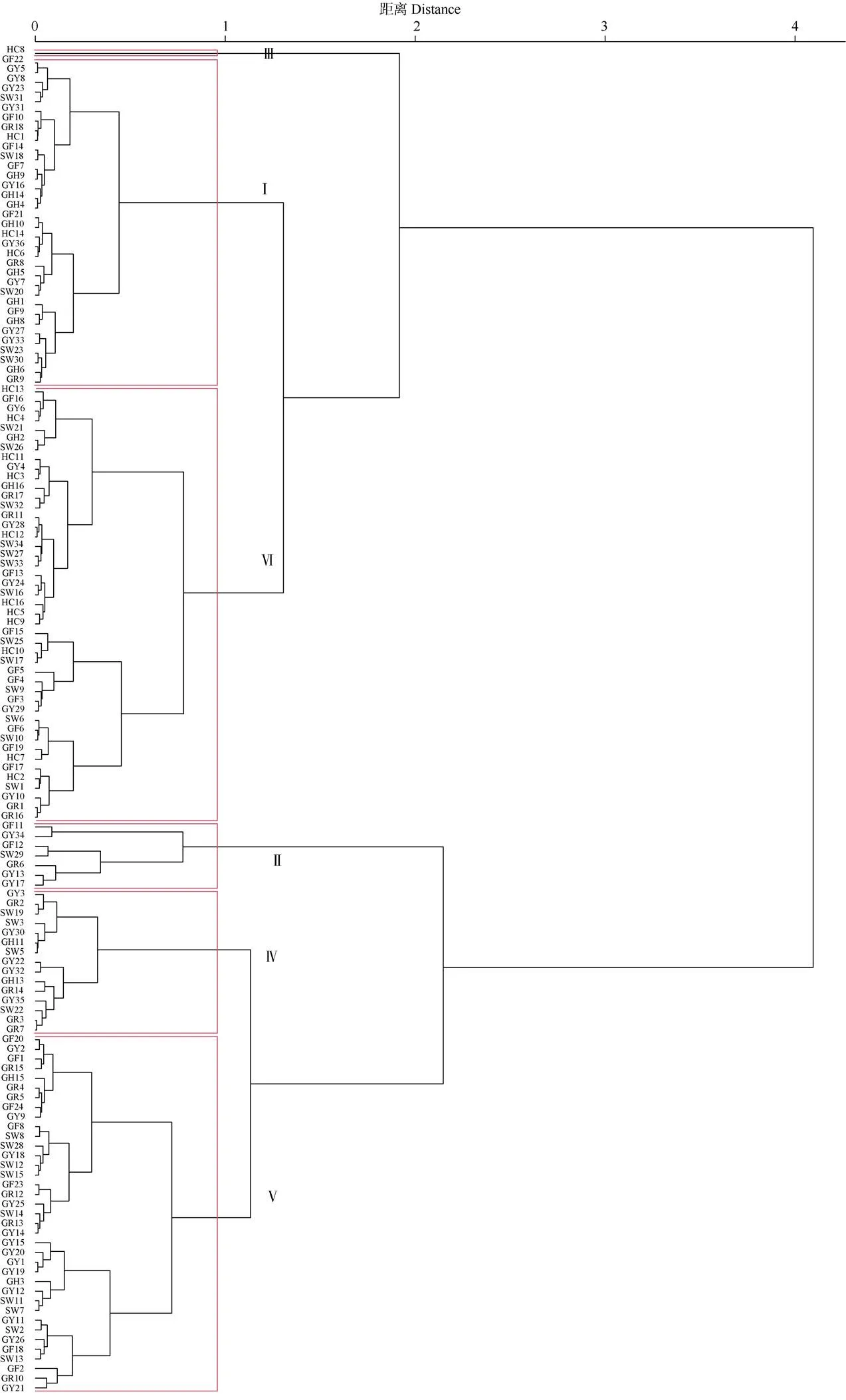
圖1 139株棱果花葉片表型性狀聚類圖。GY1~GY36、SW1~SW3、SW5~SW23、SW25~SW34、GR1~GR18、GH1~GH6、GH8~GH11、GH13~GH16、GF1~GF24、HC1~HC14和HC16見表1。
3 結論和討論
群體變異系數在一定程度上反映出群體性狀的差異性及多樣性,變異系數值越大,說明群體離散程度越大,相應的性狀越不穩定;變異系數值越小,說明群體相對穩定,一致性較好[15–17]。目前, 已有較多利用葉片表型性狀對種質資源進行遺傳多樣性分析的報道[2–4,18–19]。文珊娜等[19]對12個灰木蓮()群體的葉片性狀進行了表型多樣性分析;唐慶蘭等[18]對10個群體斑皮檸檬桉(subsp.)葉片性狀進行變異分析;徐斌等[2]對37個杜鵑紅山茶()無性系的葉片主要性狀進行了遺傳多樣性分析。本研究表明139個棱果花種質資源的葉片表型性狀變異系數為14%~84%,平均為32%,說明這些葉片性狀在棱果花種質資源中變異較大,表型多樣性水平豐富,可以選出葉片觀賞性狀較高的優良品種。下一步將對資源的鑒定和創新進行研究, 利用高通量和分子生物學手段評價各個資源的遺傳背景及其特性,通過常規雜交或現代生物技術手段對高觀賞性狀進行改良利用研究,研發具有自主知識產權的新品種,對華南地區花卉產業健康發展具有重要的意義。
葉片性狀相關性和主成分分析是植物表型多樣性分析的重要方法,在植物表型多樣性及其進化研究方面發揮了重要作用。相關性分析可了解不同性狀間相互作用關系,如果2個性狀間為顯著的正相關關系,說明一個性狀的改良將有助于另一個性狀的應用,表現為協同效應,反之則表現為拮抗作用[20]。本研究分析的棱果花不同葉片表型性狀間的相關值較高,如葉面積、葉周長與葉長間的相關系數為0.75~0.99。主成分分析通過降維處理將多個變量轉成為少量能代表大部分變量信息的方法。通過對葉片表型性狀進行主成分分析,共提取了5個主成分,累計貢獻率達到88%,表明各性狀的累計貢獻率增長比較迅速,說明在棱果花的性狀變異具有多樣性。綜上所述,根據葉片各主成分貢獻率選出了影響較大的性狀指標,葉片面積、葉片周長、葉片寬、葉片長/寬和葉形參數是棱果花分類最主要的標準和依據。
聚類分析能夠將性狀相近的種質資源聚為一類,每一類都有各自的表型特征[21–22]。育種學家可根據不同的選育目標選擇親緣關系較遠的親本配置雜交組合,進而篩選出具有雜種優勢的新品種。離差平方和法能夠將139個單株完全區分開,并在閾值為1時將棱果花種質資源劃分為6類,每個類型內棱果花種質資源表型性狀類似,而不同類型中存在明顯的差異。通過對棱果花各類群的分析發現,部分來自同一個種群的材料分布在不同的類群,該結果表明聚類與地理來源無明顯的關聯。因此,在后續育種材料選擇上不僅考慮種群,還要注重材料的遺傳多樣性分析,才能更有利于棱果花種質資源收集和利用。
[1] MOU H X, HOU X C, LIU Q Z. Study on the phenotype diversity of woody energy plant[J]. For Res, 2007, 20(3): 350–355. [牟洪香, 侯新村, 劉巧哲. 木本能源植物文冠果的表型多樣性研究 [J]. 林業科學研究, 2007, 20(3): 350–355. doi: 10.3321/j. issn:1001-1498.2007.03.010.]
[2] XU B, PENG L X, YANG H X, et al. Genetic diversity analysis for leaf main traits of[J]. Bull Bot Res, 2015, 35(5): 730–734. [徐斌, 彭莉霞, 楊會肖, 等. 杜鵑紅山茶葉片主要性狀的遺傳多樣性分析 [J]. 植物研究, 2015, 35(5): 730–734. doi: 10.7525/j.issn. 1673-5102.2015.05.015.]
[3] CHEN W W, XIA T F, SUN X X, et al. The phenotypic diversity analysis ofgermplasm leaves [J]. Mol Plant Breed, 2021, 19(11): 3800–3809. [陳偉文, 夏騰飛, 孫秀秀, 等. 文昌錐種質資源葉片表型多樣性分析 [J]. 分子植物育種, 2021, 19 (11): 3800–3809. doi: 10.13271/j.mpb.019.003800.]
[4] QIU Q Q, FENG Y F, WU C Y. Genetic diversity of leaf phenotypic traits in Jujube germplasm resources [J]. Xinjiang Agric Sci, 2021, 58(2): 282–293. [仇倩倩, 馮一峰, 吳翠云. 棗種質資源葉表型性狀遺傳多樣性分析 [J]. 新疆農業科學, 2021, 58(2): 282–293. doi: 10. 6048/j.issn.1001-4330.2021.02.010.]
[5] YIN J M, WANG Z N, YANG G S, et al. Germplasm resources of wild Melastomataceae ornamental plants and their development and utili- zation in Hainan Island [J]. Chin J Trop Agric, 2006, 26(6): 63–66. [尹俊梅, 王祝年, 楊光穗, 等. 海南野牡丹科野生觀賞植物種質資源及其開發利用 [J]. 熱帶農業科學, 2006, 26(6): 63–66. doi: 10.3969/j. issn.1009-2196.2006.06.018.]
[6] PENG D H, ZHANG Q X, HUANG J T. Melastomataceae ornamental plant germplasm resources in China and the distribution investigation in Fujian Province [J]. Chin Landsc Archit, 2007, 23(11): 83–88. [彭東輝, 張啟翔, 黃俊婷. 中國野牡丹科觀賞植物種質資源及其在福建省的分布初步調查 [J]. 中國園林, 2007, 23(11): 83–88. doi: 10. 3969/j.issn.1000-6664.2007.11.022.]
[7] CAO Y, PENG Z H. Preliminary studies on ornamental plant germ- plasm resources of Melastomataceae in Hunan Province [J]. Hunan For Sci Technol, 2010, 37(1): 30–33. [曹瑜, 彭重華. 湖南省野牡丹科野生觀賞植物初步研究 [J]. 湖南林業科技, 2010, 37(1): 30–33. doi: 10.3969/j.issn.1003-5710.2010.01.009.]
[8] DAI S P, LIU L H, LIU H, et al. Investigation and evaluation on the wild ornamental resource of Melastomataceae in Guangdong [J]. J Fujian For Sci Technol, 2012, 39(4): 121–126. [代色平, 劉連海, 劉慧, 等. 廣東省野牡丹科植物資源調查與評價 [J]. 福建林業科技, 2012, 39(4): 121–126.]
[9] ZHAO J, QIU S, LI X J, et al. The germplasm resource of Melasto- mataceae ornamental plants in Guangxi and its development prospect [J]. J Hebei Agric Sci, 2008, 12(7): 80–82. [趙健, 仇碩, 李秀娟, 等. 廣西野牡丹科觀賞植物種質資源及開發前景 [J]. 河北農業科學, 2008, 12(7): 80–82. doi: 10.3969/j.issn.1088-1631.2008.07.032.]
[10] LIN Q J, LIN X X, SU J Q, et al. Comprehensive appraisal on ornamental characteristics and adaptability of plants in family Melasto- mateae [J]. J SW For Univ, 2010, 30(5): 33–37. [林秋金, 林秀香, 蘇金強, 等. 16種野牡丹科植物觀賞性及適應性綜合評價 [J]. 西南林學院學報, 2010, 30(5): 33–37. doi: 10.3969/j.issn.2095-1914.2010. 05.008.]
[11] SU C L, ZHANG S K, HU Q Y, et al. The comprehensive appraisal of Melastomataceae species on cultivation characteristics [J]. For Environ Sci, 2020, 36(2): 94–100. [蘇純蘭, 張尚坤, 胡秋艷, 等. 16種野牡丹科植物栽培性狀綜合評價 [J]. 林業與環境科學, 2020, 36(2): 94– 100. doi: 10.3969/j.issn.1006-4427.2020.02.014.]
[12] XIE D S, ZHANG W W, LI S Y, et al. Introduction and comprehensive evaluation of 23 species of wild Paeoniaceae [J]. Jiangsu Agric Sci, 2019, 47(14): 164–168. [謝東升, 張彎彎, 李仕裕, 等. 23種野生野牡丹科植物引種和綜合評價 [J]. 江蘇農業科學, 2019, 47(14): 164– 168. doi: 10.15889/j.issn.1002-1302.2019.14.038.]
[13] R Core Team. A Language and Environment for Statistical Computing [M]. Vienna: R for Statistical Computing, 2019.
[14] GILMOUR A, CULLIS B, WELHAM S, et al. An efficient computing strategy for prediction in mixed linear models [J]. Comput Stat Data Anal, 2004, 44(4): 571–586. doi: 10.1016/S0167-9473(02)00258-X.
[15] OU H B, LIN J Y, LI J, et al. Analysis of variation of leaf phenotypic traits in different provenances of[J]. SW China J Agric Sci, 2020, 33(3): 637–644. [歐漢彪, 林建勇, 李娟, 等. 不同種源楠木葉片表型性狀變異分析 [J]. 西南農業學報, 2020, 33(3): 637–644. doi: 10.16213/j.cnki.scjas.2020.3.026.]
[16] LI Y C, FU N F, SUN J Z, et al. Intraspecific phenotypic diversity in[J]. Bull Bot Res, 2021, 41(5): 775–788. [李雁瓷, 付乃峰, 孫加芝, 等. 秋海棠()的種內表型多樣性 [J]. 植物研究, 2021, 41(5): 775–788. doi: 10.7525/j.issn.1673-5102.2021. 05.016.]
[17] UPADHYAYA H D, YADAV D, DRONAVALLI N, et al. Mini core germplasm collections for infusing genetic diversity in plant breeding programs [J]. Electr J Plant Breed, 2010, 1(4): 1294–1309.
[18] TANG Q L, LIANG L H, DENG Z Y, et al. Variation analysis on leaf morphologic characters of different provenance ofssp.[J]. Mol Plant Breed, 2022, 20(16): 5454–5462. [唐慶蘭,梁麗華, 鄧紫宇, 等. 不同種源斑皮檸檬桉葉片形態變異分析[J]. 分子植物育種, 2022, 20(16): 5454–5462. doi: 10.13271/j.mpb.020. 005454.]
[19] WEN S N, ZHONG C L, JIANG Q B, et al. Phenotypic diversity analysis of seedling leaf traits ofDandy [J]. Bull Bot Res, 2017, 37(2): 288–297. [文珊娜, 仲崇祿, 姜清彬, 等. 灰木蓮種源幼苗葉片性狀表型多樣性分析 [J]. 植物研究, 2017, 37(2): 288–297. doi: 10.7525/j. issn.1673-5102.2017.02.018.]
[20] PENG Y H, LU G, HAO H K, et al. Analysis on leaf nutrient elements of differentprovenances [J]. J SW For Univ, 2012, 32(5): 37–41. [彭玉華, 路剛, 郝海坤, 等. 不同種源紅錐葉片營養元素分析 [J]. 西南林業大學學報, 2012, 32(5): 37–41. doi: 10.3969/j. issn.2095-1914.2012.05.008.]
[21] PENG F, ZHAO M L, XU C X, et al. Genetic diversity analysis of spinach germplasm resources based on phenotypic traits [J]. Mol Plant Breed, 2021, 19(5): 1698–1708. [彭楓, 趙孟良, 徐晨曦, 等. 基于表型性狀的菠菜種質資源遺傳多樣性分析 [J]. 分子植物育種, 2021, 19(5): 1698–1708. doi: 10.13271/j.mpb.019.001698.]
[22] RUIZ D, EGEA J. Phenotypic diversity and relationships of fruit quality traits in apricot (L.) germplasm [J]. Euphytica, 2008, 163(1): 143–158. doi: 10.1007/s10681-007-9640-y.
Phenotypic Diversity Analysis ofGermplasm Resources in Main Natural Distribution Area
XU Bin1, YI Lijiang2, LIN Zizhan3, CHEN Xinyu1, LIAO Huanqin1, PAN Wen1, YANG Huixiao1*
(1. Guangdong Provincial Key Laboratory of Silviculture, Protection and Utilization/Guangdong Academy of Forestry,Guangzhou 510520, China; 2. Guangdong Shaotoujiao Forest Station,Shenzhen 518081, Guangdong, China; 3. Management Department of Ehuangzhang Provincial Nature Reserve, Yangchun 529631, Guangdong, China)
In order to understand the diversity of leaf phenotypic traits ingermplasm resources, the variance analysis, correlation analysis, principal component analysis and cluster analysis were conducted based on leaf traits of 139 individuals from 6 populations. The results showed that there were significant differences in leaf phenotypic traits among population groups and individuals. The variation coefficients of leaf traits ranged from 0.14 to 0.84 with average of 0.32, suggesting that the leaf traits ofvaried greatlyLeaf traits could be extracted into five principal components, with a cumulative contribution rate of 88%, in which leaf area, leaf perimeter, leaf width, leaf length/width and leaf shape parameters were the main factors caused phenotype difference.Cluster analysis showed that 139 individuals ofresources could be divided into 6 groups, there were 34 individuals in group Ⅰ, 7 in group Ⅱ, 1 in group Ⅲ, 15 in group Ⅳ, 37 in group Ⅴ, and 45 in group Ⅵ, which showed that the individuals from different population were scattered in different groups, indicating that there were no significant relationship between leaf phenotype and source. Therefore, the leaf phenotypic diversity was abundant among different populations and individuals, which could provide basis for genetic improvement and new variety breeding.
; Germplasm; Leaf trait; Correlation analysis
10.11926/jtsb.4574
2021-11-22
2022-03-14
廣東省林業科技創新項目(2020KJCX002)資助
This work was supported by the Project for Forestry Science Technology Innovation in Guangdong (Grant No. 2020KJCX002).
徐斌(1974年生),男,博士,教授級高級工程師,主要從事木本花卉遺傳育種方面的研究。E-mail: xubin@sinogaf.cn
通訊作者 Corresponding author.E-mail: hxyang@sinogaf.cn
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
電子制作(2018年18期)2018-11-14 01:48:24
資源再生(2017年3期)2017-06-01 12:20:59
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
