杜仲莖和子葉的含膠細胞形態(tài)學研究
2023-04-29 00:44:03張明鑒岳婭東高順凱甘敏董益陽張繼川
林業(yè)科學研究 2023年3期
張明鑒 岳婭東 高順凱 甘敏 董益陽 張繼川
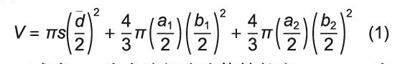
摘要:[目的]進一步認識杜仲成熟含膠細胞形態(tài)特征,并為解釋杜仲含膠細胞起源、發(fā)育和成熟過程提供細胞形態(tài)學證據(jù)。[方法]利用整體觀察法和組織切片法對杜仲莖、種子萌芽期子葉和幼苗期子葉進行研究,分析和比對杜仲含膠細胞形態(tài)。[結(jié)果]取得杜仲莖皮層中14個成熟的含腔細胞形態(tài)圖像、46個含膠細胞膨大端圖像和2個成熟中含膠細胞圖像;取得杜仲種子萌芽期子葉中原始細胞形態(tài)和成熟中含膠細胞圖像、杜仲幼苗期子葉中成熟含膠細胞圖像以及杜仲含膠細胞透射電子顯微鏡圖像;杜仲莖中含膠細胞軸長度范圍為981~3 167 um,體積范圍為1 x10-4~3 x 10-4 mm3;膨大端橢球體短軸長度范圍為8.6~22.4um,橢圓形的長軸與短軸之比范圍為1.08~1.77。[結(jié)論](1)杜仲莖皮層中的成熟含膠細胞是一種由細長的圓柱狀胞體和近似為橢球體的2個膨大端構(gòu)成的無分支一非鉸接乳管細胞。(2)透明后的含膠細胞大部分無內(nèi)部結(jié)構(gòu),小部分存在低折光率的內(nèi)部結(jié)構(gòu)。(3)杜仲子葉中含膠細胞起源于一連串啞鈴狀的原始細胞,杜仲子葉中含膠細胞生成過程存在一系列細胞形態(tài)變化。
關(guān)鍵詞:杜仲;含膠細胞;細胞形態(tài)學
中圖分類號:Q944.6,Q949.751.5 文獻標識碼:A 文章編號:1001-1498(2023)03-0050-12
杜仲(Eucommia ulmoides Oliv.)屬于杜仲科杜仲屬,進化上相對保守,是一種古老的樹種。杜仲在我國已有2000多年栽培歷史,廣泛分布于我國多省區(qū)。杜仲組織中存在一定數(shù)量的含膠細胞,含膠細胞中儲存有天然橡膠的異構(gòu)體杜仲膠(反式+1,4-聚異戊二烯),故以杜仲葉片和果實作為主要原料提取杜仲膠。天然橡膠作為一種重要的戰(zhàn)略物資,由于國際橡膠市場供需關(guān)系日趨緊張,嚴重影響我國天然橡膠資源安全,因此,大力發(fā)展杜仲膠產(chǎn)業(yè)對于保障我國的天然橡膠資源安全具有重要意義。大力開發(fā)杜仲膠產(chǎn)業(yè),需要發(fā)展杜仲種植業(yè)、提升杜仲種質(zhì)資源品質(zhì),這就要求研究人員從植株、細胞和分子等水平系統(tǒng)地研究杜仲產(chǎn)膠過程,為杜仲膠生成機制研究提供支撐,從而能夠在根本上提升杜仲的膠產(chǎn)量和品質(zhì)。目前,對于杜仲產(chǎn)膠機制的相關(guān)研究主要集中在基因、蛋白質(zhì)、代謝組和植株水平,細胞形態(tài)學水平研究較為缺乏。
杜仲含膠細胞主要分布在莖、葉、根、果實及雄蕊組織中。早在古代,我國的勞動人民就已經(jīng)發(fā)現(xiàn)杜仲莖皮存在“折之內(nèi)有白絲相連”的現(xiàn)象。史自強等采用溴-碘-冰醋酸變性試劑處理后進行石蠟切片的方法觀察到了棕色變性橡膠絲。田蘭馨等采用石蠟切片法在杜仲1年生莖初生韌皮部觀察到含膠細胞,采用薄切片法對杜仲1年生莖中含膠細胞發(fā)生和發(fā)育進行了研究,并描述了莖中含膠細胞自原始細胞發(fā)育至成熟過程的細胞內(nèi)部結(jié)構(gòu)變化。崔躍華等、申延等也在1年生莖和當年生果實中觀察到了類似細胞內(nèi)部結(jié)構(gòu)變化。崔躍華等還采用堿溶液離析組織的方法對果皮、葉片和葉柄的含膠細胞長度進行了離柝測量;周莉英等采用溴-碘-冰醋酸試劑變性處理后以堿溶液離析葉片中含膠細胞束的方法,對不同月份葉片中含膠細胞的長度進行了動態(tài)分析。Nakazawa等采用激光共聚焦顯微技術(shù)對杜仲莖組織切片進行熒光染色分析,認為杜仲含膠細胞內(nèi)部橡膠粒子的融合過程和反式-1,4-聚異戊二烯的積累同時進行,并提出了一種含膠細胞反式-1,4-聚異戊二烯合成及積累過程的模型,分為細胞分化和伸長、橡膠粒子堆積、橡膠粒子充滿乳管并融合三個階段;利用掃描電子顯微鏡技術(shù)對杜仲莖中含膠細胞斷面進行研究,認為反式-1,4-聚異戊二烯最初以棒狀粒子形式在含膠細胞中合成,并隨著細胞成熟而轉(zhuǎn)變?yōu)槔w維狀結(jié)構(gòu),最終充滿細胞腔。趙喜源也通過斷面表征觀察到含膠細胞內(nèi)部的橡膠粒子。杜仲含膠細胞的內(nèi)部橡膠粒子是積累天然高分子聚異戊二烯的細胞器,其發(fā)育機制尚未有相關(guān)文獻報道。橡膠粒子的基本結(jié)構(gòu)與脂質(zhì)體一致,內(nèi)部為疏水橡膠球芯,外部為單層生物膜。單層生物膜作為相界面防止顆粒聚集。杜仲含膠細胞在乳管分類中屬無分支-非鉸接型乳管。無分支-非鉸接型乳管在生長中原基伸長成一個不分枝的管道,通過侵入性生長方式生長。前人對杜仲含膠細胞的解剖結(jié)構(gòu)、起源分化和杜仲膠積累機制進行了一定的研究,但關(guān)于杜仲含膠細胞的起源、發(fā)育和成熟過程的細胞形態(tài)特征變化過程尚且不足。本研究通過利用整體觀察法和組織切片法對杜仲莖、種子萌芽期子葉和幼苗期子葉的含膠綱胞的起源、發(fā)育和成熟過程的細胞形態(tài)特征進行了更深一步的研究,以期為解釋含膠細胞起源、發(fā)育和成熟過程提供細胞形態(tài)學證據(jù)。
1材料與方法
1.1材料與儀器
實驗材料:2年生杜仲莖(2021年5月15日取自北京市杜仲公園內(nèi)野生杜仲戒樹,取樣后于FFA固定液中固定7 d);當年生杜仲嫩莖(取自播種204 d的杜仲幼苗,取樣后于FFA固定液中固定7d);杜仲種子(杜仲果實于2021年10月采集于北京市杜仲公園,去殼后備用);杜仲葉片(2020年7月29日取材于北京大學內(nèi)野生杜仲成樹)。
實驗儀器:Leica CM1950冰凍切片機(徠卡公司,德國),徠卡DMil倒置數(shù)碼顯微鏡(徠卡公司,德國)。
實驗試劑:次氯酸鈉溶液·鹽酸變性試劑(等體積的23.2 mmol.L-1 NaClO溶液和2.0 mol·L-1HCI溶液,使用時混勻),蘇丹III染色液(3.4mmol.L-1的蘇丹III的乙醇溶液),F(xiàn)FA固定液(福爾馬林:冰醋酸:70%酒精:丙三醇=5:5:90:5)。
1.2方法
1.2.1杜仲莖皮層樣品制備與圖像提取 取直徑3 mm左右杜仲莖,切下節(jié)間并完整環(huán)剝杜仲莖皮,以去離子水清洗;加入次氯酸鈉溶液.鹽酸變性試劑,混勻后靜置3d完成變性;變性后莖皮去除木栓層保留皮層,使用去離子水清洗2次;使用蘇丹III染色液對變性后皮層染色12 h,以8.6mol·L-1乙醇溶液洗去浮色,制作臨時裝片;染色后的杜仲莖皮層壓平并干燥后,使用二甲苯(天津大茂)透明30 min,后采用中性樹膠封片劑(上海懿洋)封固制成永久裝片;采用數(shù)碼生物顯微鏡對裝片中的含膠細胞結(jié)構(gòu)進行拍照,后用Adobe Photoshop CC 2019對提取的圖像進行拼合。
1.2.2含膠細胞形態(tài)數(shù)據(jù)提取和體積計算 含膠細胞直線長度l為含膠細胞2個膨大端最遠點之間距離,代表含膠細胞在組織中占據(jù)的空間位置;含膠細胞軸長度為2個膨大端的長軸長a1、a2和為胞體切線垂線被胞體所截線段中點連線s(以下簡稱中線)長度之和共同構(gòu)成,代表著胞體實際生長所占據(jù)的內(nèi)部空間長度;直線長度與軸長度之比代表著含膠細胞胞體的曲折程度。基于以上定義對1.2.1過程提取的含膠細胞圖像進行形態(tài)數(shù)據(jù)提取。
杜仲含膠細胞是一種“十分細長、兩端膨大、內(nèi)部充滿硬橡膠顆粒的絲狀單細胞”,故將含膠細胞2個膨大端近似為橢球體,脆體近似為圓柱體,含膠細胞中體積V估算式為式(1)。對1.2.1過程提取的含膠細胞圖像的參考圖1D并按照式(1)進行體積估算。
式中:s為含膠細胞胞體軸長度,a1、a2為2個橢球體的長軸長,b1、b2為2個橢球體的短軸長,a為胞體平均直徑。
同時對含膠細胞體積和膨大端體積占比進行計算。采用Adobe Photoshop CC 2019軟件對含膠細胞胞體像素進行統(tǒng)計并換算為X-Y平面的投影面積S。將X-Y平面的投影面積S除以含膠細胞胞體軸長度s得到胞體平均直徑d。
1.2.3當年生杜仲莖冷凍切片的制作與圖像提取 取當年生杜仲莖樣品,以去離子水清洗;加人次氯酸鈉溶液一鹽酸變性試劑,混勻后靜置24 h完成變性;變性后樣品使用去離子水清洗后,以蘇丹III染色液整體染色12h,使用8.6 mol·L-1乙醇溶液洗去浮色;使用冷凍包埋劑垂直包埋變性整體染色后的樣品,采用冷凍切片機對樣品進行切片,在濕潤的載玻片上展開切片,用數(shù)字顯微成像系統(tǒng)對杜仲組織進行拍照。
1.2.4杜仲種子萌芽期子葉樣品制備與圖像提取 恒溫培養(yǎng)箱中萌發(fā)的杜仲種子以FFA周定液固定2d;取固定后種子萌芽期子葉,以去離子水清洗,加入11.4 mmol·L-1 NaClO溶液,靜置2d至子葉至透明無色完成去除脂質(zhì)并變性;將處理后子葉樣品徒手剝離上表皮,制作臨時裝片;用數(shù)碼生物顯微鏡對含膠細胞結(jié)構(gòu)進行拍照,Adobe Photoshop CC 2019對提取的圖像進行拼合及數(shù)據(jù)測量。
取固定后種子萌芽期子葉,以去離子水清洗;加入次氯酸鈉溶液.鹽酸變性試劑,混勻后靜置2d完成變性;使用去離子水清洗,以蘇丹III染色液染色12 h,使用8.6 mol·L-1乙醇溶液浸泡1h洗去浮色;將整體染色后子葉移至培養(yǎng)皿內(nèi)的載玻片上,滴加1.31 mol·L-1 NaOH溶液覆蓋子葉表面處理1h,之后吸去NaOH溶液;使用棉簽破壞子葉表皮細胞并用去離子水洗去組織碎片,留下染色后的絲狀物制作臨時裝片;用數(shù)碼生物顯微鏡對含膠細胞結(jié)構(gòu)進行拍照,Adobe Photoshop CC 2019對提取的圖像進行拼合及數(shù)據(jù)測量。
1.2.5杜仲幼苗期子葉樣品制備與圖像提取 萌發(fā)的杜仲種子栽種于土壤中,1個月后得到杜仲幼苗。杜仲幼苗期子葉以FFA固定液固定2d;取固定后幼苗期子葉樣品,以去離子水清洗;加入次氯酸鈉溶液-鹽酸變性試劑,混勻后靜置1d完成變性;使用去離子水清洗,以蘇丹III染色液染色12 h,使用8.6 mol·L-1乙醇溶液洗去浮色,制作臨時裝片;用數(shù)碼生物顯微鏡對含膠細胞結(jié)構(gòu)進行拍照,Adobe Photoshop CC 2019對提取的圖像進行拼合及數(shù)據(jù)測量。
1.2.6杜仲含膠細胞的透射電子顯微鏡檢測 新鮮杜仲葉片組織投入固定液后,送樣武漢賽維爾生物科技有限公司對葉片主脈制作切片并以生物透射電鏡進行檢測。
2結(jié)果與分析
2.1莖皮層中含膠細胞的形態(tài)結(jié)果分析
含膠細胞在杜仲莖皮層內(nèi)大量存在,部分含膠細胞單獨生長,與其他含膠細胞無糾纏。取得的3個單獨生長的含膠細胞外部形態(tài)的完整高分辨率圖像見圖1。杜仲含膠細胞南寬度均勻的細長胞體和近似為橢球體的2個膨大端構(gòu)成;胞體基本與杜仲莖的軸方向平行,結(jié)構(gòu)呈近似直線狀或折線狀;自然狀態(tài)下的含膠細胞整體無色透明,經(jīng)變性整體染色法處理后細胞整體被蘇丹III染成紅色。含膠細胞形態(tài)結(jié)果符合以往文獻中對成熟含膠細胞形態(tài)的敘述,故其為含膠細胞的成熟形態(tài)。
對取得的14個成熟含膠細胞照片進行統(tǒng)計分析。統(tǒng)計結(jié)果顯示:莖皮層中杜仲成熟的含膠細胞軸長度范圍為981~3 167 um,直線長度范圍為813~3149 um,直線長度與軸長度之比范圍為1:1.00~1:1.21.平均軸長度為2 086.9 Um。
在杜仲莖皮層中觀察到與含膠細胞具有相同染色的特殊結(jié)構(gòu)見圖2、圖3。圖2表明:結(jié)構(gòu)具有兩端膨大、胞體細長的特征,與成熟的含膠細胞特征相同;直線長度為1196.8 um,達到成熟含膠細胞直線長度,故這種特殊結(jié)構(gòu)屬于含膠細胞;含膠細胞在其細長的胞體上具有多個膨大節(jié),成熟含膠細胞并不具備,因此,圖2屬于含膠細胞的成熟過程中間形態(tài),是成熟中的含膠細胞。圖3顯示:該結(jié)構(gòu)存在10個較為密集的膨大節(jié),相鄰膨大節(jié)之間直接相連或由一段長度較短的胞體相連,因此,是另一個成熟中含膠細胞的局部圖像。圖3中虛線標注的亞結(jié)構(gòu)呈啞鈴狀,由一段28.9 um的細長胞體和2個膨大節(jié)構(gòu)成,結(jié)構(gòu)兩端最遠處距離為61.6 um;啞鈴狀結(jié)構(gòu)右端膨大節(jié)與另一個膨大節(jié)直接相連;左端膨大節(jié)與膨大節(jié)同樣直接相連,但在2個膨大節(jié)間出現(xiàn)了長度為18 um的胞體結(jié)構(gòu)。圖3中啞鈴狀亞結(jié)構(gòu)與其直接相連的2個膨大節(jié)間存在無胞體結(jié)構(gòu)和有胞體結(jié)構(gòu)2種情況,代表相鄰亞結(jié)構(gòu)的膨大節(jié)之間的相連過程先于相鄰的啞鈴狀結(jié)構(gòu)之間胞體出現(xiàn)的過程。對比圖2和圖3結(jié)果,圖2含膠細胞膨大節(jié)更少、分布更稀疏,膨大節(jié)間胞體長度更長。
對46個含膠細胞膨大端照片進行統(tǒng)計分析,結(jié)果顯示:膨大端橢球體短軸長度(膨大端寬度)范圍為8.6~22.4 um,橢圓形的長軸與短軸之比范圍為1.08~1.77。
莖皮層中含膠細胞局部圖像見圖4所示。含膠細胞周同有一定的空隙(圖4A);經(jīng)二甲苯透明后含膠細胞膨大端大部分胞質(zhì)均勻無內(nèi)部結(jié)構(gòu)(圖4B),少部分在橢球體中心處有球狀結(jié)構(gòu),在顯微鏡光源照射下顯得十分明亮(圖4C);大多數(shù)胞體無內(nèi)部結(jié)構(gòu)(圖4D黑色箭頭處),少部分在中線處有連續(xù)的內(nèi)部結(jié)構(gòu),在顯微鏡光源照射下顯得十分明亮(圖4D白色箭頭處),其余部分胞質(zhì)染色均勻。
圖4C和圖4D中含膠細胞內(nèi)部存在光照下顯得明亮的結(jié)構(gòu)現(xiàn)象,類似于玻璃經(jīng)光照后夾雜的氣泡呈明亮狀的現(xiàn)象。分析光在含膠細胞內(nèi)外的傳播情況:顯微鏡光源發(fā)出的光由于受到周圍環(huán)境影響,到達含膠細胞表面時光線方向各異。將含膠細胞胞體近似為圓柱體,當光于細胞表面進入細胞質(zhì)時,此時含膠細胞細胞質(zhì)的亮度與周圍組織未有明顯差異,代表永久裝片中覆蓋在樣品表面的中性樹膠封片劑(n0.20:1.521 6~1.524 0)和二甲苯(nD.25:1.493 25~1.502 95)透明處理后的周嗣組織與二甲苯透明處理后的含膠細胞細胞質(zhì)的折射率差異較小,光只發(fā)生了曲面折射。當光由含膠細胞細胞質(zhì)進入內(nèi)部結(jié)構(gòu)時,此時觀察到含膠細胞胞體中線處具有呈明亮內(nèi)部結(jié)構(gòu),代表相當一部分的光在二甲苯透明后的細胞質(zhì)與內(nèi)部結(jié)構(gòu)兩相界面上發(fā)生了全反射現(xiàn)象。全反射現(xiàn)象的發(fā)生條件之一為光由光密介質(zhì)進入光疏介質(zhì),故含膠細胞內(nèi)部結(jié)構(gòu)的折射率小于含膠細胞細胞質(zhì)的折射率。綜上所述,該內(nèi)部結(jié)構(gòu)在光源照射的情況下顯得十分明亮,是因為其折射率小于二甲苯透明后的含膠細胞細胞質(zhì)的折射率,故該內(nèi)部結(jié)構(gòu)與二甲苯透明的細胞質(zhì)折射率更低且存在明顯的相界面。
當年生杜仲莖的整體染色冷凍切片結(jié)果見圖5,含膠細胞(染色后呈紅色部分)主要分布在皮層韌皮部,有的含膠細胞單獨生長,有的含膠細胞聚集生長。單獨生長的含膠細胞胞體垂直于軸方向生長時其胞體截面近似圓形,故含膠細胞胞體可近似為圓柱狀處理(圖5白色箭頭處)。
2.2莖皮層中含膠細胞的體積計算
莖皮層中3個單獨生長含膠細胞圖像見圖1A、B、C所示,對其進行形態(tài)學數(shù)據(jù)測量,參考圖1D并按照式(1)進行體積估算,同時計算膨大端體積占含膠細胞體積比例w,結(jié)果見表1。
經(jīng)體積估算得到單個含膠細胞體積范圍為1×10-4~3×10-4 mm3。含膠細胞膨大端長軸遠小于整個含膠細胞的長度,短軸長卻為胞體平均寬度的2倍左右。計算結(jié)果顯示,膨大端在含膠細胞總體積中占據(jù)相對較大的比例(4%~11%)。
2.3杜仲種子萌芽期子葉和幼苗期子葉含膠細胞的形態(tài)結(jié)果分析
種子萌芽期子葉局部顯微圖像見圖6。在種子萌芽期子葉靠近胚一端處(圖6B紅框位置)觀察到子葉中原位生成的含膠細胞束向杜仲胚方向延伸的一連串細胞結(jié)構(gòu)(圖6A),該結(jié)構(gòu)遠離含膠細胞集束,在子葉中孤立存在。由于含膠細胞原位生成并以侵人性生長方式協(xié)調(diào)地生長,同時隨著植物的生長而伸長,故在種子萌芽期子葉中,原位發(fā)生的含膠細胞束中的含膠細胞結(jié)構(gòu)位置越遠離集束、在子葉中存在越孤立、結(jié)構(gòu)伸長的越短,則其發(fā)育時期越早。因此,一連串含膠細胞結(jié)構(gòu)可以確定為含膠細胞的早期結(jié)構(gòu),是含膠細胞的最初狀態(tài)。含膠細胞的早期結(jié)構(gòu)可以分為兩個部分:圖6A結(jié)構(gòu)右側(cè)存在三個中間彎曲細長、兩端膨大的啞鈴狀結(jié)構(gòu),結(jié)構(gòu)間緊鄰但不連續(xù),細胞質(zhì)較周邊組織有明顯的折射率差異。圖6A中從右至左的3個啞鈴狀結(jié)構(gòu)兩端最遠距離分別為63.5、38.2、48.9 um;3個結(jié)構(gòu)內(nèi)部細長處寬度在5 um左右,具有較大長寬比,基本符合田蘭馨文獻中原始細胞敘述。其中,有側(cè)第一個啞鈴狀結(jié)構(gòu)顯著不對稱,其靠近胚的一端寬度達到16.9 um,尺寸接近于含膠細胞膨大端短軸長,而另一端寬度僅為8.5 um。圖6A結(jié)構(gòu)左側(cè)存在一個連續(xù)結(jié)構(gòu),右端微有膨大但與啞鈴狀原始細胞間兩端距離緊密卻存在界限;胞身含有2個膨大節(jié),寬度分別為12.9um和10.9 um;兩個膨大節(jié)間胞體長度為40.9 um。
杜仲幼苗期子葉含膠細胞圖顯示:杜仲子葉中的成熟含膠細胞胞體細長,具有不規(guī)則形態(tài)的膨大端,細胞整體均一、透明、染色均勻(圖7A)。有在螺紋導(dǎo)管存在的葉脈區(qū)域,發(fā)現(xiàn)眾多具有與成熟含膠細胞相同染色的斷續(xù)結(jié)構(gòu),其近似構(gòu)成了一條曲線(圖78紅色框線處)。相比種子萌芽期子葉,幼苗期子葉中含膠細胞密度更低,可以清晰地看到單獨生長的含膠細胞。
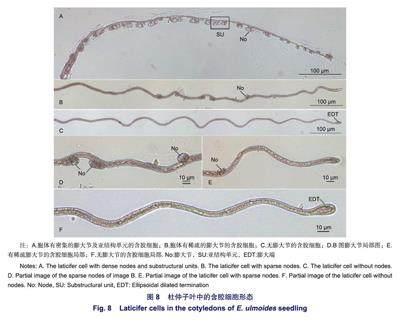
以圖6A結(jié)構(gòu)作為含膠細胞發(fā)育過程形態(tài)研究起點,圖7A含膠細胞作為發(fā)育過程形態(tài)研究終點,分析解離的染色后含膠細胞束中的含膠細胞成熟過程的中間形態(tài)。含膠細胞特殊形態(tài)如圖8所示:圖8A、B、C分別是3種有代表性的成熟中含膠細胞結(jié)構(gòu)。圖8A得到的結(jié)構(gòu)類似于圖6A,整體結(jié)構(gòu)連續(xù),其中,右側(cè)第1個啞鈴狀結(jié)構(gòu)顯著不對稱,右端寬度為11.9 um,左端寬度為5.9 um,兩端最遠距離為45.2 um;通過對比圖8A結(jié)構(gòu)的自身差異發(fā)現(xiàn),距結(jié)構(gòu)右側(cè)最遠端距離86.7 um至273.3 um區(qū)域內(nèi)的膨大節(jié)間胞體較其他位置胞體有明顯的伸長跡象。圖8B中結(jié)構(gòu)僅有5個膨大節(jié),膨大節(jié)分布稀疏,相鄰2個膨大節(jié)之間的胞體長度存在顯著差異;胞體一端未見明顯膨大端。圖8C是一種連續(xù)的但不含膨大節(jié)的結(jié)構(gòu),除存在內(nèi)部結(jié)構(gòu)外,整體形態(tài)與成熟的含膠細胞相同,橢圓形的膨大端出現(xiàn)。圖8A、B、C含膠細胞結(jié)構(gòu)均具有相連的類似含膠細胞胞體的連續(xù)結(jié)構(gòu),內(nèi)部存在一定的內(nèi)部結(jié)構(gòu),區(qū)別在于膨大節(jié)有無、相鄰膨大節(jié)之間胞體長度差異和膨大端的有無。由此可以認為,圖8A、B、C分別顯示的是3個處于不同發(fā)育時期的含膠細胞,圖8A和圖8B的差異可能是由于隨著含膠細胞逐漸成熟,不對稱的啞鈴狀原始細胞結(jié)構(gòu)消失,膨大節(jié)之間胞體伸長及連接相鄰亞結(jié)構(gòu)單元的胞體伸長造成的。由以上結(jié)果對其發(fā)育時序排序為:圖8A-圖8B-圖8C。故以發(fā)育時期先后順序?qū)z細胞形態(tài)圖像排序為:圖6A-圖8A-圖8B-圖8c-圖7A。
圖8D為圖8B胞體膨大節(jié)局部放大圖。圖8E為膨大節(jié)分布稀疏且無膨大端的含膠細胞局部圖像,含膠細胞與圖8B含膠細胞處于同一發(fā)育時期,其細胞內(nèi)部存在不連續(xù)的橢球狀結(jié)構(gòu)。圖8F為無膨大節(jié)、有膨大端的含膠細胞的局部圖像,含膠細胞外形已非常接近成熟的含膠細胞,但內(nèi)部具有一些平行結(jié)構(gòu),該結(jié)構(gòu)于含膠細胞末端膨大區(qū)匯合。平行的內(nèi)部結(jié)構(gòu)形成,可能代表著含膠細胞進入了新的發(fā)育時期。
2.4透射電子顯微鏡照片結(jié)果分析
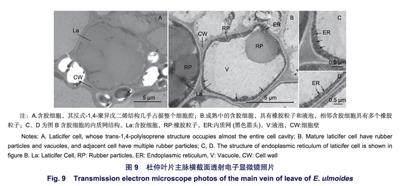
圖9為杜仲葉片主脈橫截面中取得含膠細胞的透射電子顯微照片。反式-1,4-聚異戊二烯結(jié)構(gòu)存在于含膠細胞細胞壁包裹的細胞腔之中。圖9A含膠細胞直徑11 um,反式-1,4-聚異戊二烯結(jié)構(gòu)占據(jù)整個細胞腔截面積的94.2%。圖9B中含膠細胞長9.1 um,寬6.9 um;含膠細胞腔截面中,液泡占據(jù)大部分面積,細胞壁內(nèi)側(cè)的細胞質(zhì)基質(zhì)將液泡環(huán)繞。細胞質(zhì)基質(zhì)中存在直徑分別為3.3 um和0.5 um的2個明顯的橡膠粒子,占細胞腔截面積的34.1%。在細胞質(zhì)基質(zhì)中多處存在平行于細胞壁和液泡膜的內(nèi)質(zhì)網(wǎng)(圖9C、D),同時在細胞質(zhì)基質(zhì)中分布著許多電子致密物質(zhì)。左側(cè)緊鄰的含膠細胞具有多個橡膠粒子,橡膠粒子周圍存在著許多深灰色點狀的電子致密物質(zhì)。
3討論
3.1杜仲莖皮層含膠細胞內(nèi)部結(jié)構(gòu)
由2.1節(jié)結(jié)果可知,杜仲莖中含膠細胞是一種兩端為橢球狀膨大端、中部為細長的圓柱狀胞體的無分支·非鉸接乳管細胞,胞體呈近似直線狀或折線狀,細胞整體基本與杜仲莖的軸方向平行,長度約在1~3 mm范圍,與田蘭馨等對當年生杜仲嫩枝中成熟含膠細胞形態(tài)的敘述一致。研究中發(fā)現(xiàn),大部分經(jīng)二甲苯透明后的含膠細胞染色均一且透明,其內(nèi)部無低折光率的內(nèi)部結(jié)構(gòu),也無其他明顯的細胞器結(jié)構(gòu)(圖4A、B),符合田蘭馨等以薄切片法研究得到“含膠細胞成熟時,細胞器解體”的結(jié)論,也符合Nakazawa等通過掃描電子顯微鏡發(fā)現(xiàn)皮層附近韌皮部的反式-1,4-聚異戊二烯呈巨大的圓柱形橫截面,認為反式-1,4-聚異戊二烯結(jié)構(gòu)最終會占據(jù)細胞壁包圍的腔室的結(jié)論,故經(jīng)二甲苯透明后無內(nèi)部結(jié)構(gòu)的含膠細胞為成熟的含膠細胞。研究中發(fā)現(xiàn),少部分經(jīng)二甲苯透明后的含膠細胞膨大端中心存在折射率低于透明后含膠細胞細胞質(zhì)的球狀結(jié)構(gòu)(圖4C),另有少部分經(jīng)二甲苯透明后的含膠細胞胞體內(nèi)部的中線處存在折射率低于透明后含膠細胞細胞質(zhì)的線狀連續(xù)結(jié)構(gòu)(圖4D),而兩類含膠細胞大部分胞質(zhì)著色與成熟含膠細胞無明顯差異。以往報道結(jié)果認為,成熟含膠細胞的細胞質(zhì)中充斥著橡膠粒子,不含細胞器。本研究2.1節(jié)中,含膠細胞經(jīng)二甲苯透明后,二甲苯滲透后與細胞質(zhì)共同組成一個有機相,該相與周圍組織折射率近似;而該有機相包裹著一個折射率更低的結(jié)構(gòu)相。該低折射率特殊結(jié)構(gòu)相的存在,說明莖皮層內(nèi)部具有成熟的外部形態(tài)的含膠細胞內(nèi)部仍具有明顯的特殊細胞器結(jié)構(gòu)。低折射率特殊結(jié)構(gòu)存在于膨大端和胞體的核心部位,故其可能與含膠細胞的細胞形態(tài)發(fā)育調(diào)控有著密切的關(guān)聯(lián)。
3.2杜仲含膠細胞生成過程的細胞形態(tài)變化
由2.3節(jié)結(jié)果顯示,杜仲種子萌發(fā)后,含膠細胞在子葉中產(chǎn)生;成熟的含膠細胞為胞體細長,兩端略有形態(tài)結(jié)構(gòu)略有不規(guī)則膨大端,結(jié)構(gòu)連續(xù)且透明的特殊細胞。該結(jié)果與趙喜源描述杜仲葉片含膠細胞“多數(shù)含膠細胞的膨大端呈倒立水滴狀,部分呈元寶狀和近似圓球狀”一致。
根據(jù)2.3節(jié)得到的含膠細胞形態(tài)圖像的發(fā)育時序?qū)Χ胖俸z細胞生成過程的細胞形態(tài)變化進行分析。圖6A所示杜仲幼苗期子葉內(nèi)一連串的含膠細胞早期結(jié)構(gòu),左側(cè)為連續(xù)結(jié)構(gòu),右側(cè)為3個互不連續(xù)的啞鈴狀原始細胞。圖6A右側(cè)第1個啞鈴狀原始細胞顯著不對稱,可能與原始細胞的分裂形成有關(guān)。圖6A結(jié)構(gòu)內(nèi)部存在的這種形態(tài)差異,代表著組織結(jié)構(gòu)可能是由不連續(xù)原始細胞融合后變?yōu)檫B續(xù)的含膠細胞。圖3連貫的結(jié)構(gòu)中存在10個明顯的膨大節(jié),膨大節(jié)分布密集,節(jié)間胞體長度短,與圖6A和圖8A結(jié)構(gòu)相似,同處于含膠細胞發(fā)育早期。圖3所示莖中含膠細胞早期結(jié)構(gòu)中,胞體內(nèi)標注的啞鈴狀結(jié)構(gòu)右端膨大節(jié)與另一個啞鈴狀結(jié)構(gòu)左端膨大節(jié)緊密相連,左端膨大節(jié)與另一個啞鈴狀結(jié)構(gòu)右端膨大節(jié)同樣緊密相連且在2個膨大節(jié)之間出現(xiàn)了較短的胞體結(jié)構(gòu),證明了啞鈴狀的原始細胞產(chǎn)生后即與相鄰的原始細胞融合連接,進而在連接處產(chǎn)生胞體。胞體產(chǎn)生后不斷伸長,故啞鈴狀結(jié)構(gòu)成為細長的含膠細胞的一部分,最終無法區(qū)分。基于啞鈴狀原始細胞的融合現(xiàn)象,本研究將融合后的原始細胞啞鈴狀結(jié)構(gòu)稱之為含膠細胞的亞結(jié)構(gòu)單元。啞鈴狀原始細胞的膨大端經(jīng)過融合降級為亞結(jié)構(gòu)單元的膨大節(jié),由亞結(jié)構(gòu)單元內(nèi)部的胞體和亞結(jié)構(gòu)單元之間的胞體相連接。圖8A中杜仲幼苗期子葉含膠細胞中各亞結(jié)構(gòu)單元的2個相鄰膨大節(jié)的胞體長度較小,連接亞結(jié)構(gòu)單元之間的胞體長度也較小。圖8B中杜仲幼苗期子葉含膠細胞中僅有5個膨大節(jié),膨大節(jié)分布稀疏,相鄰2個膨大節(jié)之間的胞體長度存在顯著差異;胞體一端未見明顯膨大端。圖8C杜仲幼苗期子葉含膠細胞除存在內(nèi)部結(jié)構(gòu)外,整體形態(tài)與成熟的含膠細胞相同,這時膨大節(jié)消失,膨大端形成,成熟的含膠細胞外形形成。故子葉中含膠細胞膨大端產(chǎn)生與成熟后期有關(guān),這一結(jié)果與田蘭馨等認為莖中含膠細胞膨大端在插人生長逐漸緩慢時形成的敘述相符。基于啞鈴狀的原始細胞相互融合成為連續(xù)的結(jié)構(gòu)后逐步發(fā)育為含膠細胞的事實,本研究認為,杜仲子葉中的含膠細胞起源于一連串啞鈴狀的原始細胞。
2.4節(jié)中結(jié)果顯示了葉片主脈中含膠細胞的兩種形態(tài)。Nakazawa等研究發(fā)現(xiàn),反式-1,4-聚異戊二烯最初以棒狀粒子形式在含膠細胞中合成,并隨著細胞成熟而轉(zhuǎn)變?yōu)槔w維狀結(jié)構(gòu),最終充滿細胞腔。基于此結(jié)果,通過對比分析含膠細胞腔截面反式-1,4-聚異戊二烯結(jié)構(gòu)的面積百分比,確定細胞所處反式-1,4-聚異戊二烯積累時期并判斷細胞發(fā)育的相對時序。圖9A中含膠細胞腔截面的反式-1,4-聚異戊二烯結(jié)構(gòu)面積為94.2%,幾乎充滿著截面;圖9B中含膠細胞腔截面反式-1,4-聚異戊二烯結(jié)構(gòu)面積為34.1%,故圖9A中含膠細胞較圖9B中含膠細胞成熟度更高。圖9B中含膠細胞細胞質(zhì)分布于細胞腔內(nèi)側(cè)并環(huán)繞著液泡;內(nèi)質(zhì)網(wǎng)在細胞質(zhì)基質(zhì)中平行于液泡和細胞壁分布;直徑為0.5 um的橡膠粒子被細胞質(zhì)基質(zhì)包裹,故橡膠粒子在細胞質(zhì)基質(zhì)中生成。液泡-細胞質(zhì)基質(zhì)-內(nèi)質(zhì)網(wǎng)-細胞質(zhì)基質(zhì)的細胞結(jié)構(gòu)分布,可能與含膠細胞積累反式-1,4-聚異戊二烯的功能有關(guān)。
基于以上分析并結(jié)合以往文獻報道,總結(jié)杜仲含膠細胞生成過程的細胞形態(tài)變化:首先在剛剛萌發(fā)的杜仲幼苗子葉中,韌皮部分生組織通過分裂方式形成一連串啞鈴狀的原始細胞。一連串細胞中延伸最遠的啞鈴狀原始細胞呈非對稱形態(tài)(圖6A);相鄰的啞鈴狀的原始細胞融合,融合后啞鈴結(jié)構(gòu)作為含膠細胞的亞結(jié)構(gòu)單元存在,形成一個連續(xù)的由多個亞結(jié)構(gòu)單元組成的含膠細胞早期形態(tài),在相鄰啞鈴狀亞結(jié)構(gòu)單元融合處形成新的胞體結(jié)構(gòu)(圖8A、圖3);在含膠細胞早期形態(tài)中,膨大節(jié)之間胞體不斷不均勻徑向延長,細胞開始插入生長,同時膨大節(jié)逐漸融人胞體(圖8B、D、E、圖2);當胞體徑向延長完成,含膠細胞中的膨大節(jié)消失,膨大端出現(xiàn),成熟含膠細胞的外觀形成,其內(nèi)部結(jié)構(gòu)逐漸規(guī)則(圖8C、F),橡膠粒子在細胞質(zhì)基質(zhì)中逐漸合成(圖9B)。當含膠細胞成熟,細胞內(nèi)橡膠粒子合成和積累完畢并相互融合,反式-1,4-聚異戊二烯充滿整個細胞腔(圖9A),細胞器解體,含膠細胞內(nèi)部變得均一且透明(圖7A)。田蘭馨等認為,原始細胞不經(jīng)融合就開始插入生長,進行胞體伸長;而本研究結(jié)果顯示啞鈴狀的原始細胞首先相互融合,成為細長胞體的亞結(jié)構(gòu)單元,再進行胞體伸長,最后膨大節(jié)消失,含膠細胞外形成熟。
在幼苗期子葉中發(fā)現(xiàn)了斷續(xù)的含膠結(jié)構(gòu)(圖7B),同時該時期子葉中含膠細胞密度較幼苗期子葉更低。幼苗期子葉中斷續(xù)的含膠結(jié)構(gòu)可能是由于子葉組織內(nèi)部的含膠細胞在某些酶及協(xié)同因子作用下形態(tài)發(fā)生轉(zhuǎn)化,一些結(jié)構(gòu)產(chǎn)生較大的變化或消失,最終導(dǎo)致含膠細胞數(shù)量下降,這種形態(tài)變化可能反映了含膠細胞的凋亡機制,與子葉作為植物幼苗發(fā)育過程能量供應(yīng)來源的功能相關(guān)。
3.3杜仲莖與子葉中含膠細胞形態(tài)對比
對杜仲莖和子葉中成熟的含膠細胞形態(tài)進行對比發(fā)現(xiàn),二者均為由細長的圓柱狀胞體和2個膨大端構(gòu)成的無分支-非鉸接乳管細胞,基本結(jié)構(gòu)相同。二者存在成熟狀態(tài)的細胞膨大端存在形態(tài)差異。杜仲莖中含膠細胞膨大端形態(tài)更加規(guī)則,接近橢球體;子葉含膠細胞形態(tài)更加不規(guī)則。
3.4實驗設(shè)計與創(chuàng)新點
杜仲含膠細胞結(jié)構(gòu)長達毫米級別且以插入方式生長,細長的細胞往往在組織空間中呈折線狀分布。田蘭馨等采用組織切片技術(shù)對含膠細胞發(fā)生和發(fā)育進行了研究,其應(yīng)用組織切片技術(shù)雖然能夠很好地確定細胞結(jié)構(gòu)在切片平面中空間位置,進而進行分析,但本研究發(fā)現(xiàn),莖中杜仲含膠細胞呈折線狀分布的特性,在采用切片研究含膠細胞外部形態(tài)時,很大程度上存在無法得到完整的含膠細胞外部形態(tài)的可能。本研究以整體觀察法研究易環(huán)剝、厚度適宜杜仲2年生莖皮層中的杜仲含膠細胞,取得了杜仲含膠細胞的完整外部形態(tài)圖像,為后續(xù)研究提供了數(shù)據(jù)支撐。以往的含膠細胞形態(tài)研究中,在含膠組織的變性過程中均采用溴-碘-冰醋酸變性劑。溴-碘-冰醋酸變性劑利用溴對橡膠雙鍵結(jié)構(gòu)進行加成反應(yīng)變性橡膠,利用溴的氧化性使組織漂白和并增加組織通透性,但該法使用的溴單質(zhì)存在劇毒、易揮發(fā)和難于稱量的缺點。本研究創(chuàng)新以次氯酸鈉溶液-鹽酸變性試劑,配置簡單,效果良好,安全性更佳。創(chuàng)新建立以次氯酸鈉溶液-鹽酸作為變性試劑、以蘇丹III的乙醇溶液作為染色液的變性整體染色法,實現(xiàn)了含膠細胞的特異性染色。本研究采用二甲苯作為透明劑對莖皮樣品進行整體透明,取得了莖皮層含膠細胞內(nèi)部結(jié)構(gòu)的整體圖像。本研究創(chuàng)新聯(lián)用變性整體染色法與冷凍切片技術(shù),以柔軟的杜仲當年生嫩莖作為材料進行切片研究,減小了制樣過程影響,染色效果更好,結(jié)果圖像的細節(jié)更豐富。
研究含膠細胞生成過程的細胞形態(tài)變化,需要確定含膠細胞處于何種發(fā)育時期,進而確定結(jié)構(gòu)變化發(fā)生的相對時序。在成熟的杜仲組織中,由于含膠細胞發(fā)生的時刻和位置隨機,故不能機械地按照含膠細胞所處組織空間位置來確定其發(fā)育時間的先后。要準確地確定某種植物幼苗中乳管或含膠細胞的起源方式,通常需要對種子和幼苗進行觀察。本研究選擇杜仲的萌芽期種子子葉作為研究含膠細胞最初起源和發(fā)育過程細胞形態(tài)變化的材料,以其中含膠細胞最初形態(tài)作為起點;選擇幼苗期子葉作為研究成熟含膠細胞形態(tài)的材料,以其中成熟的含膠細胞作為終點。進而在杜仲萌芽期子葉中尋找含膠細胞發(fā)育過程各時期的中間形態(tài),從而形成完整的含膠細胞起源、發(fā)育和成熟機制的細胞形態(tài)學證據(jù)鏈。本研究針對種子萌芽期子葉中的脂類物質(zhì),采用次氯酸鈉溶液作為漂白變性試劑并利用其堿性去除了脂類物質(zhì),有效地透明了種子萌芽期子葉,取得了清晰的含膠細胞早期結(jié)構(gòu)原位觀察結(jié)果。針對萌芽期子葉存在許多集束生長、纏繞度高且不利于整體觀察的含膠細胞束的問題,以堿性離析法將子葉破碎,游離含膠細胞,尋找到含膠細胞發(fā)育過程各時期的中間形態(tài)。
4結(jié)論
杜仲莖皮層中成熟的含膠細胞是一種由細長的圓柱狀胞體和近似為橢球體的2個膨大端構(gòu)成的無分支一非鉸接乳管細胞;透明后的含膠細胞大部分無內(nèi)部結(jié)構(gòu),小部分存在低折光率的內(nèi)部結(jié)構(gòu)。杜仲子葉中含膠細胞起源于一連串啞鈴狀的原始細胞。杜仲子葉中含膠細胞生成過程的細胞形態(tài)變化過程包括:啞鈴狀原始細胞相互融形成含膠細胞早期形態(tài);啞鈴狀亞結(jié)構(gòu)單元間形成胞體,隨著胞體徑向延長,細胞插入生長,膨大節(jié)逐漸減少;當含膠細胞胞體徑向延長完成,胞體膨大節(jié)消失,膨大端出現(xiàn),成熟的含膠細胞外觀形成,細胞內(nèi)部結(jié)構(gòu)逐漸規(guī)則;當含膠細胞成熟,細胞內(nèi)橡膠粒子合成和積累完畢并相互融合,細胞器解體,反式-1,4-聚異戊二烯充滿整個細胞腔,含膠細胞變得均一且透明。幼苗期子葉中斷續(xù)的含膠結(jié)構(gòu)可能反映了含膠細胞的凋亡機制。
