5株植物內生真菌生物轉化檸檬烯和月桂烯產物分析
2023-05-12 10:59:28楊道茂王奇志裴真巧
華僑大學學報(自然科學版) 2023年3期
關鍵詞:植物
楊道茂, 王奇志, 裴真巧
(華僑大學 化工學院, 福建 廈門 361021)
萜類化合物的基本結構是由不同個數的異戊二烯首尾相連而成.根據異戊二烯規則,由2個異戊二烯構成的化合物稱為單萜.單萜類物質主要存在于揮發油中,廣泛應用于醫藥工業、香料工業、昆蟲信息素及昆蟲驅避劑等方面[1].某些單萜類化合物,如檸檬烯[2]、月桂烯、哌烯等,由于結構簡單、價格低廉,一直是理想的生物轉化底物模型,均可用細菌、真菌、酵母菌和植物細胞完成轉化[3].在已報道的微生物催化劑中,細菌占41%,真菌占33%[4].
在用于生物轉化的細菌中,有關假單胞屬細菌(Pseudomonassp.)的報道最多.該屬細菌能以檸檬烯[5-6]、酪醇[7-8]和月桂烯[9-10]為轉化底物.2013年,Molina等[11]總結了假單胞屬細菌生物轉化萜類物質的研究.真菌中,黑曲霉[12-13]、尖孢鐮刀菌[14-15]、指狀青霉菌[16-17]、解脂耶氏酵母[18-20]等微生物都有轉化單帖類化合物的相關報道.隨著生物轉化研究的深入,植物內生真菌在生物催化領域的作用開始受到重視.植物內生真菌因為需要與宿主植物一起共進化來適應環境的變化,從而顯示它們可能會產生豐富的酶以適應宿主[21].現有多種植物內生真菌應用于生物轉化,如海藻(Bostrychiaradicans)來源內生真菌Botryosphaeriasp.[22],植物內生真菌(Phomopsissp.,Glomerellacingulata,Diaporthephaseolorum,Aspergillusfumigatus)[23],火炬松來源植物內生真菌(Phomopsissp.)[24].因此,采用植物內生真菌生物轉化單萜類化合物理論上是可行的[25].
本文選取單環單萜中的(+)-檸檬烯和(-)-檸檬烯、無環單萜的月桂烯為轉化底物,以假臭草來源5株植物內生真菌為轉化菌株,采用氣相色譜-質譜聯用技術(GC-MS)分析轉化產物,以期為工業生物催化尋找合適的微生物菌種.
1 材料與方法
1.1 藥品與儀器
(+)-檸檬烯、(-)-檸檬烯、月桂烯,日本東京化成工業株式會社;無水乙醇、石油醚(60~90 ℃)、乙酸乙酯,國藥試劑有限公司;(1S,2S,4R)-(+)-二戊烯-1,2-二醇(檸檬烯-1,2-二醇,CAS號:38630-75-0),美國Sigma-Aldrich公司.以上試劑均為分析純.0.22 μm濾膜,天津市津騰實驗設備有限公司;GF254型薄層層析硅膠板(50 mm×200 mm),山東青島海洋化工廠分廠.
ZQZY-75CN型振蕩培養箱,上海支楚儀器有限公司;XH-C型旋渦混合器,江蘇省金壇市白塔新寶儀器廠;SW-CJ-1FD型潔凈工作臺,江蘇省蘇州安泰空氣技術有限公司;H1650型醫用離心機,湖南湘儀實驗室儀器開發有限公司;Angilent 8860 GC System-5977B型氣質聯用儀器(帶G4513A型自動進樣器),美國安捷倫科技有限公司;EYELA N-1100型旋轉蒸發儀,日本東京理化器械株式會社.
1.2 微生物與培養基
植物內生真菌(AlternariaNeesPS01,AlternariaNeesPS06,AlternariaNeesPS07,DiaportheNitschkePS10,AlternariaNeesPL14),分離自假臭草莖干部位,由王奇志博士饋贈.
發酵及轉化培養基:蛋白胨10 g·L-1,葡萄糖40 g·L-1,pH值為4.0~6.0.
1.3 檢測方法
薄層層析(TLC)方法:展開劑采用體積比V(石油醚)∶V(乙酸乙酯)=10∶1;顯色劑采用體積分數為1%的香草醛濃硫酸.
氣相色譜-質譜聯用檢測采用Agilent HP-5ms型色譜柱(30 m×250 μm×0.25 μm),載氣為氦氣,進樣量為1 μL.升溫程序:60 ℃保持1 min,20 ℃·min-1升溫到300 ℃,保持13 min.MS離子源溫度為230 ℃,MS四極桿溫度為150 ℃,質譜掃描范圍(m/z)為60~800.
2 實驗步驟
2.1 液體發酵法生物轉化單萜類化合物
實驗過程參考Rottava等[13]的做法并進行略微調整.菌種在馬鈴薯葡萄糖瓊脂培養基中活化6 d后,挑取一接種環菌絲接種于裝有100 mL培養基的250 mL錐形瓶中,在30 ℃,200 r·min-1下培養6 d;然后,在培養基中加入1 mL經0.22 μm濾膜過濾除菌的(+)-檸檬烯-乙醇溶液(檸檬烯與乙醇的體積比為1∶1)使檸檬烯的體積分數為0.5%,繼續培養4 d完成轉化過程;轉化結束,將發酵液過濾,取3 mL濾液到15 mm×150 mm玻璃試管中,用等體積乙酸乙酯萃取;接著,將有機相轉移到干燥玻璃試管中,并用無水硫酸鈉脫水;最后,取1 mL有機相進行TLC和GC-MS檢測.
(-)-檸檬烯和月桂烯的轉化及代謝產物處理操作與(+)-檸檬烯的處理步驟一致.
2.2 建立檸檬烯-1,2-二醇標準曲線
用色譜純乙酸乙酯溶解標準品配制質量濃度為8.6 g·L-1的母液,采用逐步稀釋法配制成不同質量濃度的樣品.采用GC-MS檢測方法建立檸檬烯-1,2-二醇的峰面積-質量濃度的標準曲線.
3 實驗結果與討論
3.1 有機相的TLC檢測
5株植物內生真菌分別轉化(+)-檸檬烯、(-)-檸檬烯和月桂烯,取其萃取相進行TLC檢測,結果如圖1所示.

(a) (+)-檸檬烯 (b) (-)-檸檬烯 (c) 月桂烯
由圖1可知:5株植物內生真菌對2種手性構型的檸檬烯底物均具有良好的轉化能力,且其產物比較單一,這有利于后期的產物分離,轉化結果比較理想;特殊的是,菌株PS07轉化(+)-檸檬烯時,與其他菌株相比,其產物多出了1個斑點,而菌株PS10轉化(-)-檸檬烯時,與其他菌株相比,其產物多出了2個淺藍色斑點;5株真菌對月桂烯均具有轉化能力,且轉化斑點的比移值(Rf)相同,推測轉化產物是相同的.
從菌株測序的結果可知,菌株PS01,PS06,PS07和PL14歸屬于AlternariaNees,菌株PS10歸屬于DiaportheNitschke.結果表明,同一種屬的真菌在轉化實驗時,其轉化產物一般是一致的,如菌株PL14,PS01,PS07,PS06在轉化檸檬烯、月桂烯的表現一樣,但菌株PS07在轉化(+)-檸檬烯時多出1個斑點.因此,各菌株在完成生物轉化實驗時,可以套用一般規律,但是不能忽視個別菌株的特殊性.
3.2 植物內生真菌對(+)-檸檬烯的轉化行為
5株植物內生真菌完成(+)-檸檬烯轉化后,其有機相的總離子色譜(TIC)圖,如圖2所示.圖2中:δ為絕對豐度;t為時間.

(a) 菌株PS01 (b) 菌株PS06 (c) 菌株PS07
由圖2可知:各峰信號主要為峰a(環氧檸檬烯)和峰b(檸檬烯-1,2-二醇),該結果與Bier等[24]報道的一致.從轉化機理上看,環氧檸檬烯是檸檬烯-1,2-二醇的前體,屬于已知的6條檸檬烯代謝路線之一[26],因此,該實驗結果是合理的.
3.3 植物內生真菌對(-)-檸檬烯的轉化行為
5株植物內生真菌轉化(-)-檸檬烯后,其有機相的TIC圖,如圖3所示.

(a) 菌株PS01 (b) 菌株PS06 (c) 菌株PS07
由圖3可知:5株植物內生真菌的主要產物均為檸檬烯-1,2-二醇,且底物(-)-檸檬烯的信號強度較高,只有菌株PS07的信號強度低,而圖2中底物(+)-檸檬烯的信號強度普遍都低,推測菌株可能更傾向于利用(+)-檸檬烯,原因在于該構型的底物為柑橘精油的主要成分.對比圖2和圖3發現,雖然底物手性不同,但是產物一樣,推測負責轉化的雙羥基加氧酶對底物無立體特異性要求.
3.4 檸檬烯-1,2-二醇標準曲線
建立檸檬烯-1,2-二醇標準曲線,如圖4所示.圖4中:S為峰面積;ρ為檸檬烯-1,2-二醇的質量濃度.由圖4得到標準曲線方程為Y=129 971.734 13X-15 963.437 46,R2=0.993.
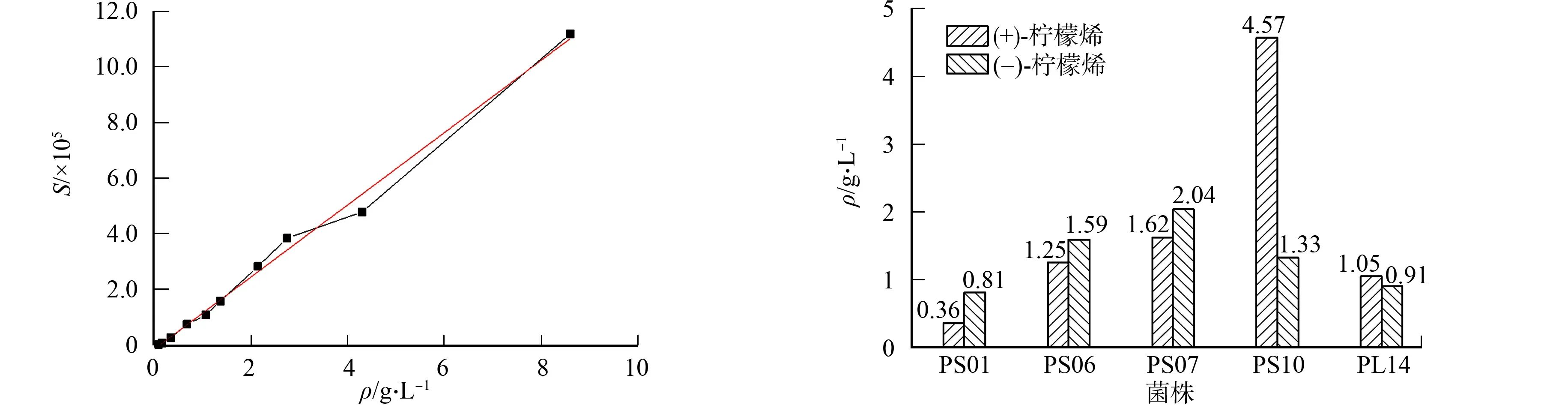
圖4 檸檬烯-1,2-二醇標準曲線 圖5 不同菌株生成檸檬烯-1,2-二醇的質量濃度
根據該標準曲線,結合圖2,3的檸檬烯-1,2-二醇峰的峰面積,計算出5株植物內生真菌轉化2種構型檸檬烯形成的檸檬烯-1,2-二醇的質量濃度,結果如圖5所示.
由圖5可知:5株植物內生真菌轉化2種手性檸檬烯生成檸檬烯-1,2-二醇的質量濃度差異較大.以(+)-檸檬烯為例,菌株PS10的產量最高,為4.57 g·L-1,是菌株PS01產量的12.7倍.如果底物為(-)-檸檬烯,則各菌株生成檸檬烯-1,2-二醇的質量濃度范圍為0.81~2.04 g·L-1.
3.5 植物內生真菌對月桂烯的轉化行為
考察5株植物內生真菌對月桂烯的轉化結果,其有機相的TIC圖,如圖6所示.

(a) 菌株PS01 (b) 菌株PS06 (c) 菌株PS07
由圖6可知:5株植物內生真菌轉化月桂烯后,與檸檬烯底物相比,其轉化產物更多樣;菌株PS01的主要產物為環氧羅勒烯,菌株PS06和PS07的主要產物為氧化芳樟醇,菌株PS10的產物為苯乙醇等化合物,菌株PL14則未檢索到相匹配的化合物.不同菌株轉化月桂烯后的產物復雜多樣,與Esmaeili等[9]的結果一致.Esmaeili等[9]采用銅綠假單胞菌轉化月桂烯,在反應1.5 d時的代謝產物為二氫芳樟醇和2,6-二甲基辛烷,反應3 d后的主要產物為2,6-二甲基辛烷和α-松油醇.由此可以看出,轉化月桂烯的產物多樣可能與轉化時間有關.
4 結論
實驗采用的5株植物內生真菌均具有生物轉化3種單萜化合物的能力.5株植物內生真菌轉化(+)-檸檬烯和(-)-檸檬烯的主要產物為檸檬烯-1,2-二醇,菌株PS10轉化(+)-檸檬烯生成檸檬烯-1,2-二醇的產量最高,為4.57 g·L-1.5株植物內生真菌轉化月桂烯的產物多樣,包含環氧羅勒烯、氧化芳樟醇等,未找到產物單一的轉化菌株.
檸檬烯-1,2-二醇具有抗癌活性[27-28],不僅能抑制CD4+和CD8+T淋巴細胞的促炎活性[29]并作為調味劑[14],還可以用來合成Prevezol C[30],是一種很重要的檸檬烯含氧衍生物.從價值上看,以Sigma-Aldrich的報價為例,底物(+)-檸檬烯的價格為1 584.8 元·L-1, 而產物檸檬烯-1,2-二醇的價格為530.09 元·g-1,轉化后的產物價值得到了極大的提升.檸檬烯-1,2-二醇理論上具有4種構型[31],用酶法轉化的產物具有單一手性的這一特性是化學合成方法無法具備的.因此,生物轉化檸檬烯合成具有更高附加值的衍生物的思路是可行且有意義的.
實驗結果表明:植物內生真菌是一種很有發展潛力的綠色生物催化劑,值得擴大底物篩選范圍并分離出更多的內生真菌用于生物轉化研究.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
