葡萄枝堆肥腐殖化的微生物養分限制機理探討
2023-05-15 06:27:14齊熙平黃懿梅賈鵬輝劉一帆謝健宇傅彬欣李建波
農業環境科學學報 2023年4期
齊熙平,黃懿梅,賈鵬輝,劉一帆,謝健宇,傅彬欣,李建波
(西北農林科技大學資源環境學院,農業農村部西北植物營養與農業環境重點實驗室,陜西 楊凌 712100)
我國的葡萄種植面積已達8.7 億hm2,每年冬季修剪下的枝條可達604 萬t[1],這些枝條含有豐富的生物質能源、常量元素和微量元素[2],其經過堆肥化處理可轉化為含腐殖質的有機肥料或土壤調理劑,用于提高土壤肥力[3]。但葡萄枝含有大量難以降解的木質素和纖維素,致使堆肥時間長、質量低[4]。高溫預處理可以破壞物料中木質纖維素的結構,增加堆肥中揮發性脂肪酸、氨基酸、多糖等營養物質的含量[5-6],促進放線菌富集以及醌類再生和縮合,增強胡敏酸聚合[7];菇渣具有相對較高的比表面積、發達的微孔結構,含有豐富的腐殖質前體物質和胞外酶,添加至堆肥中有利于有機物的降解和腐殖質的形成[8],Meng等[9]的研究表明,添加15%菇渣處理使污泥堆肥中胡敏酸含量增幅顯著高于未添加菇渣處理。
好氧堆肥實質上是一個由微生物主導的有機物分解并生成穩定腐殖物質的過程[10],微生物分泌的多種胞外酶可以分解復雜底物中最具限制性的養分,將高分子量有機化合物降解成可同化的分子從而促進系統中微生物養分循環和能量流動[11]。最常見的參與C、N、P循環的相關胞外酶包括β-1,4-葡萄糖苷酶(BG)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)、亮氨酸氨基多肽酶(LAP)和堿性磷酸酶(AP)[12]。胞外酶化學計量比,即與C、N、P 等營養物質獲取相關的胞外酶比率,可以反映微生物的營養需求和對有限資源的可用性[11]。Wang 等[12]發現添加沼渣和生物炭的稻草堆肥中參與C、N 和P 轉化的4 種胞外酶的活性都顯著提高,有機質分解加快,BG∶AP和(NAG+LAP)∶AP 比值相比對照顯著降低,微生物對P 的需求增加,受到P 的限制,微生物會分泌更多的堿性磷酸酶來維持內穩態。然而,從胞外酶活性及其化學計量變化的微生物養分限制的角度探討堆肥過程中影響腐植酸形成的機理研究還較少,葡萄枝中添加菇渣和使用高溫預處理之后的堆肥腐殖化效果及其微生物養分限制機理尚不清楚。
因此,本研究以葡萄冬剪枝條為試材,雞糞為調理劑,探討添加菇渣以及高溫預處理對堆肥過程中理化參數、腐植酸含量、胞外酶活性及其化學計量變化的影響,揭示堆肥過程中微生物胞外酶活性及其化學計量與腐植酸形成之間的關系,為豐富堆肥腐殖化機理和促進葡萄枝的高效肥料化利用提供理論依據和數據支撐。
1 材料與方法
1.1 堆肥材料及其基本性質
新鮮雞糞取自楊凌永順養雞場;葡萄枝和香菇菇渣分別來自西北農林科技大學葡萄酒學院葡萄教學標本圃和陜西省楊凌知君菌業科技公司,使用前將其粉碎至2~3 cm。物料基本理化性質見表1。

表1 堆肥原料的理化性質Table 1 Physicochemical properties of compost raw materials
1.2 堆肥方法及樣品采集
試驗共設3 個處理:對照組(CK),葡萄枝與新鮮雞糞按鮮質量比約1∶1 混合后添加適量水,調節堆肥初始含水率為50%~60%,C/N 為30∶1~35∶1,總質量18 kg;添加菇渣組(M),在CK 基礎上添加15%(濕質量)的菇渣;高溫預處理組(H),按照CK 的比例,將原料混合加熱至90 ℃且持續4 h 后,轉入堆肥反應器堆肥。3組物料混合均勻后置于3個60 L密閉強制通風好氧發酵裝置中堆肥45 d,裝置外使用保溫棉進行保溫以減少熱量損失。采用機械強制間歇曝氣,每1 h曝氣5 min,曝氣速率為73 L·min-1。堆制期間不補充水分。每日9:00 和17:00 用電子溫度計于堆體上、中、下部測定溫度(T),取其平均值作為當日的堆溫,同時測定環境溫度。根據溫度變化分別在第0、7、14、21、28、35、44 天取樣,取樣后手動翻堆,分別于堆肥物料上、中、下部采集樣品,每份樣品約200 g,樣品混勻之后分為兩份:一份于-20 ℃儲存用于測定含水率、pH、電導率(EC)、種子發芽指數、溶解性有機碳(DOC)、-N、-N 以及胞外酶活性;另一份風干后用球磨儀研磨,過1 mm 篩,用于測定總有機碳(TOC)、總氮(TN)、總磷(TP)、腐植酸碳及其組分。
1.3 測定指標及其方法
稱取新鮮樣品2 g,將樣品含水率折算后,按照干物質質量與液體體積比1∶10 加入相應體積的去離子水,180 r·min-1振蕩1 h,過濾得浸提液。使用配有pH Intelli CAL 電極和EC Intelli CAL 電極的HQ11d電位儀測定浸提液的pH 和EC。將10 顆蘿卜種子均勻散布在墊有濾紙的9 cm 培養皿中,吸取5 mL 浸提液浸潤,在25 ℃避光條件下培養72 h 后測定發芽率和根長,計算種子發芽指數,種子發芽指數=(試驗樣種子發芽數×試驗樣種子平均根長)/(對照樣種子發芽數×對照樣種子平均根長)×100%。
含水率采用烘干法測定;TN 采用凱氏定氮法測定;TP采用釩鉬黃比色法測定。DOC用TOC 儀(日本島津TOC-L)測定。樣品經KCl(2 mol·L-1)浸提稀釋后采用流動分析儀測定-N 與-N[13]。有機質采用馬弗爐灼燒減重法測定(550 ℃,6 h),測定值除以系數1.724 得TOC 含量[14]。腐植酸碳、胡敏酸碳(HA-C)及富里酸碳(FA-C)采用科諾諾娃法測定[15]。腐殖化率=腐植酸碳含量/總有機碳含量×100;腐殖化指數=胡敏酸碳含量/總有機碳含量×100。
胞外酶活性使用多功能酶標儀(瑞士Tecan Infinite M200 PRO 型)通過96 微孔板熒光法測定[16]。測定指標包括1種參與C 循環的酶:β-1,4-葡萄糖苷酶(BG),通過4-MUB-β-D-葡萄糖苷的釋放量來表示;2種參與N循環的酶:β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和亮氨酸氨基多肽酶(LAP),分別通過4-MUB-N-乙酰基-β-D-葡萄糖苷和L-亮氨酸-7-氨基-4-甲基香豆素鹽酸鹽的釋放量來表示;1種參與P循環的酶:堿性磷酸酶(AP),通過4-MUB-磷酸的釋放量來表示。
1.4 數據處理與分析
試驗數據采用Excel 2019 進行統計與分析;采用SPSS 22.0中的單因素方差分析(One-way ANOVA)進行組間的顯著性檢驗;采用OriginPro 2021 制圖。冗余分析采用Canoco 5 完成;采用Amos 24.0 構建結構方程模型,并使用Powerpoint 2019繪制轉化關系圖。
2 結果與分析
2.1 堆肥溫度、含水率、EC、pH、種子發芽指數的變化
堆肥過程中高溫期維持時間長有助于殺滅寄生蟲和病原體[17],提高堆肥效率。如圖1 所示,CK、M、H分別于第3、8、4天達到高溫期(>50 ℃),且高溫期分別保持了19、15、19 d,符合《糞便無害化衛生要求》(GB 7959—2012)。添加菇渣推遲了堆體進入高溫期的時間,其原因可能是菇渣中存留的菌絲以及某些致病菌或污染物影響了堆體土著微生物的生長[18]。

圖1 堆肥過程中溫度的變化Figure 1 Changes of temperature during composting
含水率變化如圖2a 所示,H 的含水率降低,略低于其他兩組。各組進入高溫期后由于有機物降解產水,含水率稍有提升,隨后逐漸下降。如圖2b 所示,第7~14 天,M 和H 的EC 值顯著高于CK(P<0.05),可能是相比CK,M 和H 中有機物分解產生的、揮發性脂肪酸-N和-N較多所致。14 d之后,3組的EC 值逐漸升高,可能是由于堆體水分降低引發了鹽離子的富集濃縮[19]。堆肥結束時,CK、M、H 的EC值分別為3.46、3.12、3.42 mS·cm-1,差異不顯著。如圖2c所示,堆肥過程中3組pH呈現先升后降的趨勢,這主要是由于微生物在分解有機氮過程中產生了堿性氨,14 d之后,-N 以氣體的形式揮發導致pH 下降,堆肥結束時,CK、M、H 的pH 分別為8.53、8.45、8.59,差異不顯著。CK、M、H 的種子發芽指數分別為103.15%、100.02%和91.00%(圖2d),差異不顯著,均達到70%以上,符合《有機肥料》(NY/T 525—2021)的要求。
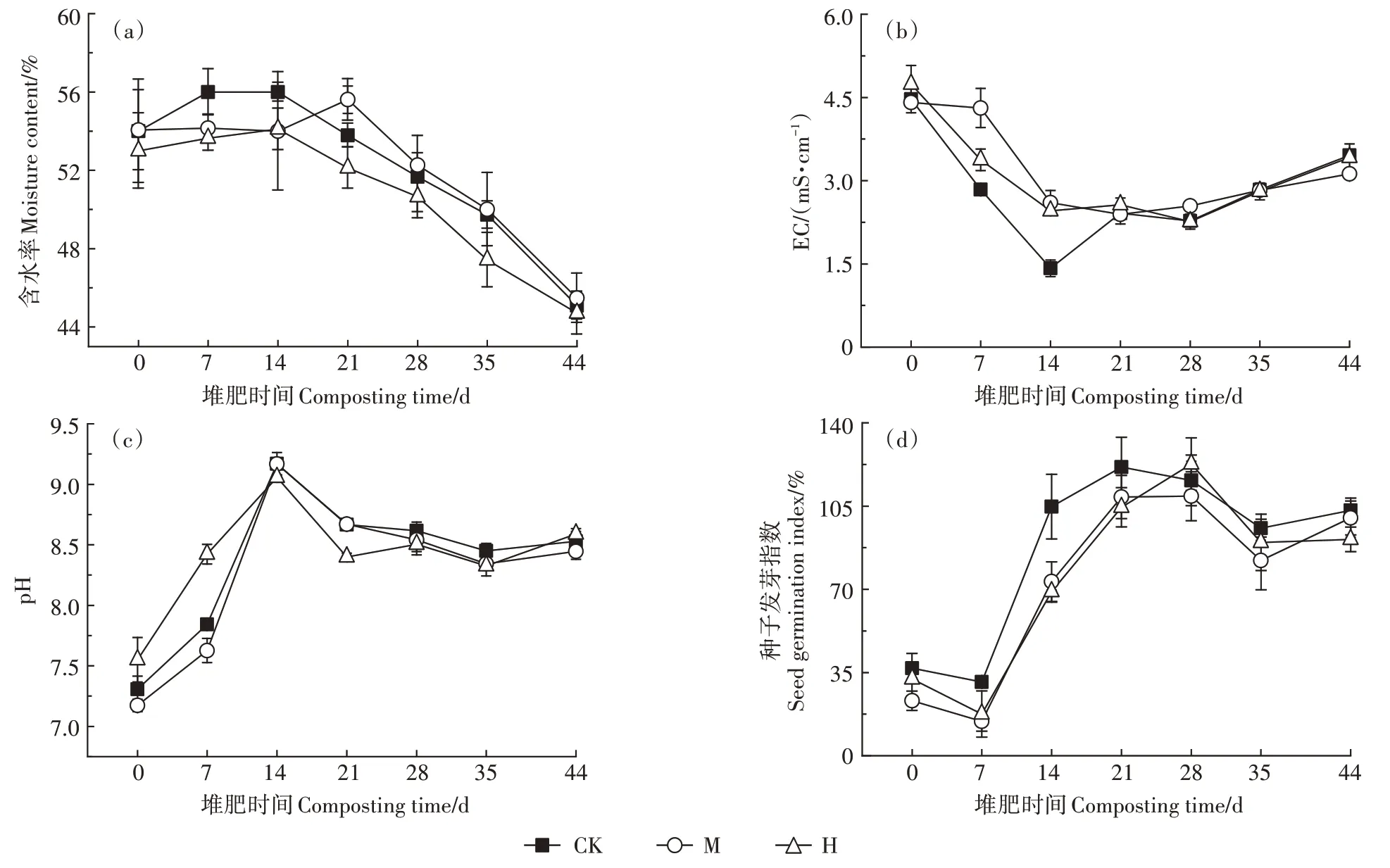
圖2 堆肥過程中含水率、pH、EC、種子發芽指數的變化Figure 2 Changes of moisture content,pH,EC and seed germination index during composting
2.2 堆肥中腐植酸碳、HA-C、FA-C含量的變化
如圖3a所示,第0天,H 的腐植酸碳含量較CK 高7.99%。第0~14 天,有機物降解劇烈,各組腐植酸碳含量迅速下降,之后逐漸上升。第14天和第28天,M和H 的腐植酸碳含量顯著高于CK(P<0.05)。最終CK、M、H 的腐植酸碳含量分別為93.03、100.81、100.90 g·kg-1,M 和H 顯著高于CK(P<0.05)。如圖3b所示,第14 天,M 和H 的HA-C 含量開始同時高于CK,直至堆肥結束。總體來說,M 和H 在第14~44 天分別較CK 提高了7.64%~37.62%和9.26%~36.96%的HA-C 含量。其中M 和H 在第21~28 天的HA-C 含量較CK 增加了7.64%~37.62%和9.47%~36.96%,增速較快,這反映了M 和H 在降溫階段促進了HA-C 的形成。最終CK、M、H的HA-C含量分別為42.71、51.70、49.29 g·kg-1,M 和H 顯著高于CK(P<0.05)。FA-C 的變化趨勢與腐植酸碳的相似,至堆肥結束,CK、M、H的FA-C 含量分別為50.32、49.11、51.61 g·kg-1,差異不顯著,與初期相比分別降低了35.77%、34.81%和40.74%。當腐殖化率≥7.0、腐殖化指數≥3.5 時堆體達到腐熟[17]。堆肥過程中腐殖化指標變化情況如表2所示,最終CK、M、H的腐殖化率和腐殖化指數分別為21.87、23.91、23.96 和10.04、12.26、11.71,均達到腐熟。相比CK,H 的腐殖化率提升了9.56%,M 的腐殖化指數提高了22.11%。

表2 堆肥過程中腐殖化指標變化情況Table 2 Changes of humification indicators during composting
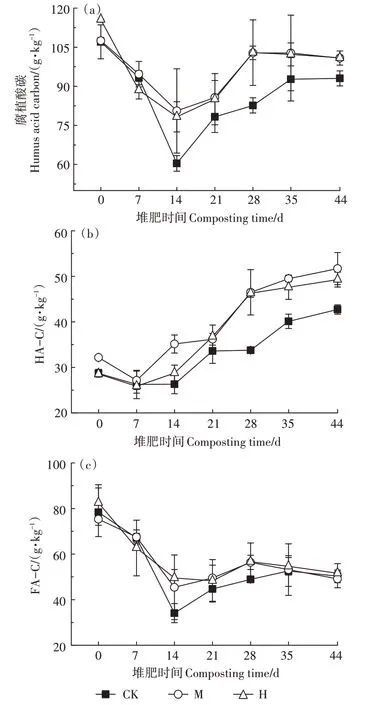
圖3 堆肥過程中腐植酸碳及其組分的變化Figure 3 Changes of humus acid carbon and its components during composting
2.3 堆肥中養分及化學計量的變化
如圖4a 至圖4c所示,3組的TOC 含量呈下降趨勢,TN 和TP含量呈上升趨勢。堆肥第14~21天,H 的TOC 含量下降速率最快,為4.35%,而CK 和M 只有1.39%和1.14%。堆肥結束時,CK、M、H 的TOC 含量分別較初始下降了8.06%、10.00%、9.19%,M 和H 的TOC降解率相比CK分別提高了1.94個和1.13個百分點。CK、M、H 初始TN 含量分別為15.31、13.50、13.21 g·kg-1,M 和H 顯著低于CK(P<0.05),7 d后,M 和H 的TN 含量開始高于CK。堆肥結束時,CK、M、H 的TN含量較初始分別提升了66.44%、107.28%、116.85%,M 和H 的TN 提升率比CK 分別高出40.84 個和50.41個百分點,TN 含量較CK 分別提高了9.81% 和12.44%。CK、M、H 的TP 含量較初始分別提升了54.09%、95.47%、125.49%,M 和H 的提升速率明顯高于CK(P<0.05),分別高出41.38 個和70.40 個百分點,最終TP含量較CK分別提高了9.84%和9.04%。
DOC 是堆肥中微生物合成自身生命體最直接的碳源,因此容易發生微生物降解[3]。各組DOC 含量在0~14 d 內快速下降(圖4d),其原因是微生物對易利用碳的快速分解利用,第14~28 天,M 和H 的DOC 含量顯著高于CK(P<0.05),可能與M 和H 中難降解有機物被降解為小分子可溶性有機物有關。堆肥結束時,各組DOC 穩定在11 g·kg-1左右,差異不顯著。由于有機氮的降解,各組-N 在第7 天稍有提升,之后隨著堆肥的進行逐漸降低,最終穩定在0.15 g·kg-1左右(圖4e),且各組間差異不顯著。各組的-N含量呈降低趨勢(圖4f),主要是堆肥中反硝化作用使-N 轉化為N2O 揮發所致,35 d 之后硝化作用增強,M 和H 的-N 含量上升,且顯著高于CK(P<0.05)。

圖4 堆肥過程中TOC、TN、TP、DOC、-N、-N的變化Figure 4 Changes of TOC,TN,TP,DOC,-N,-N during composting
如圖5a 所示,第0 天,CK、M、H 的C∶N 分別為30.22、34.71、35.10,M 和H 顯著高于CK(P<0.05),原因是菇渣本身C∶N 較高以及高溫預處理期間會使一部分氮素以氣體形式損失。隨著堆肥進行,有機碳的降解速率高于有機氮的降解速率,C∶N逐漸降低。第14 天,CK、M、H 的C∶N 分別是29.33、27.63、25.25,H下降最快,較第0天下降了28.06%。堆肥結束時各組C∶N 穩定在15 左右,M 和H 顯著低于CK(P<0.05)。如圖5b 所示,第0 天,M 和H 的C∶P 顯著高于CK(P<0.05),第21~35 天,M 的C∶P 高于其他兩組。最終3組的C∶P 穩定在50 左右,且差異不顯著。如圖5c 所示,3 組的N∶P 比值從堆肥開始到結束變化較小,都在3左右,M和H的N∶P在整個堆肥期間都高于CK。
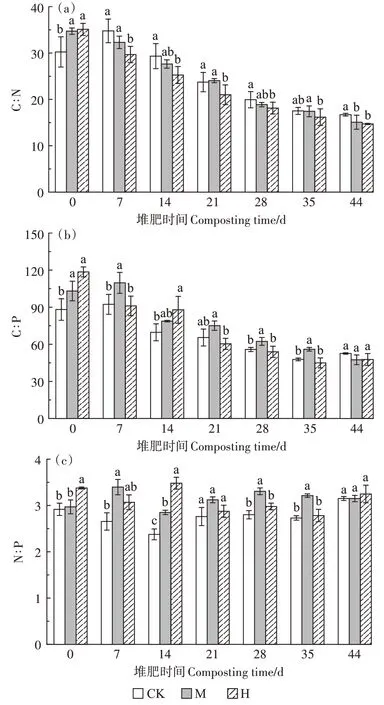
圖5 堆肥過程中化學計量的變化Figure 5 Changes of stoichiometry during composting
2.4 堆肥中胞外酶活性及其化學計量的變化
BG 可將寡糖水解成單糖,為微生物提供可利用的底物和能源[20];NAG可以催化細胞壁主要成分幾丁質和肽聚糖的水解[21],主要用于獲取微生物容易利用的N[22]。LAP 能切割蛋白質中的肽鍵,在蛋白水解中起重要作用[12]。AP 是堆肥過程中參與P 循環的主要磷酸酶,可將有機磷水解成可被植物吸收和代謝的不同形式的磷酸鹽[23]。
從圖6可以看出,第0~7天,高溫預處理導致H的各種酶活性較CK 顯著降低(P<0.05),而添加菇渣可顯著提升各種酶的活性(P<0.05)。第14 天,CK 和M中的AP 活性受高溫影響較前期略微下降,而H 中的不降反升,并從此時到堆肥結束H 的AP 活性一直高于CK。第14天直至堆肥結束,H的BG活性顯著高于其他兩組(P<0.05),且在高溫期時也有較大的活性(1 540.15 nmol·h-1·g-1),比CK 和M 分別高8.97%~13.07% 和9.82%~44.87%。第14天溫度達到56~58℃,各組的NAG活性急劇下降,而LAP活性同時升高,CK、M、H的LAP活性較第0天分別提升了176.33%、128.48%、368.76%,M 和H 的LAP 活性顯著高于CK(P<0.05)。第21 天直至堆肥結束,3 組的NAG活性都較高,除第28天外,其他時間下H的NAG活性顯著高于CK(P<0.05)。第28~44 天,M 的LAP活性高于另外兩組。
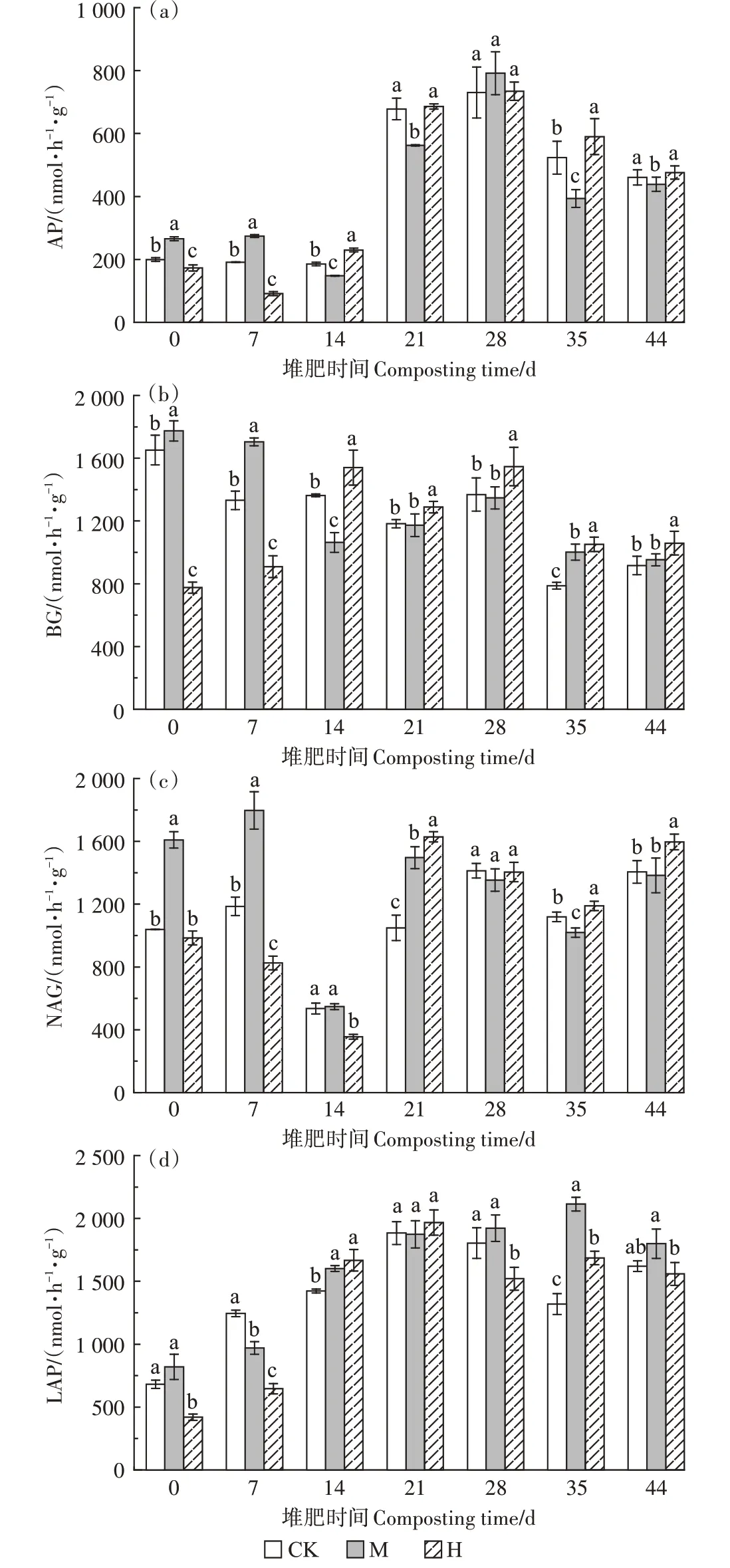
圖6 堆肥過程中酶活性的變化Figure 6 Changes of enzyme activity during composting
如圖7所示,堆肥第0天,CK、M、H的BG∶(NAG+LAP)比率分別為1.00、0.74、0.55,BG∶AP 比率分別為8.63、6.83、4.40,(NAG+LAP)∶AP比率分別為8.69、9.15、7.96,相比CK,M 和H中的微生物受到N和P 養分的限制。第14 天,M 的BG∶(NAG+LAP)顯著低于CK,(NAG+LAP)∶AP顯著高于CK,與CK相比M中的微生物受到N的限制;H的BG∶AP 和(NAG+LAP)∶AP 顯著低于CK,與CK 相比H 中的微生物受到P 的限制。從第21 天開始到堆肥結束,3 組的BG∶(NAG+LAP)、BG∶AP、(NAG+LAP)∶AP 比值變化無較大起伏,第44 天,CK、M、H 的酶促C∶N∶P活性比為1.00∶3.22∶0.45、1.00∶3.22∶0.47、1.00∶2.78∶0.40,各組間無顯著差異。
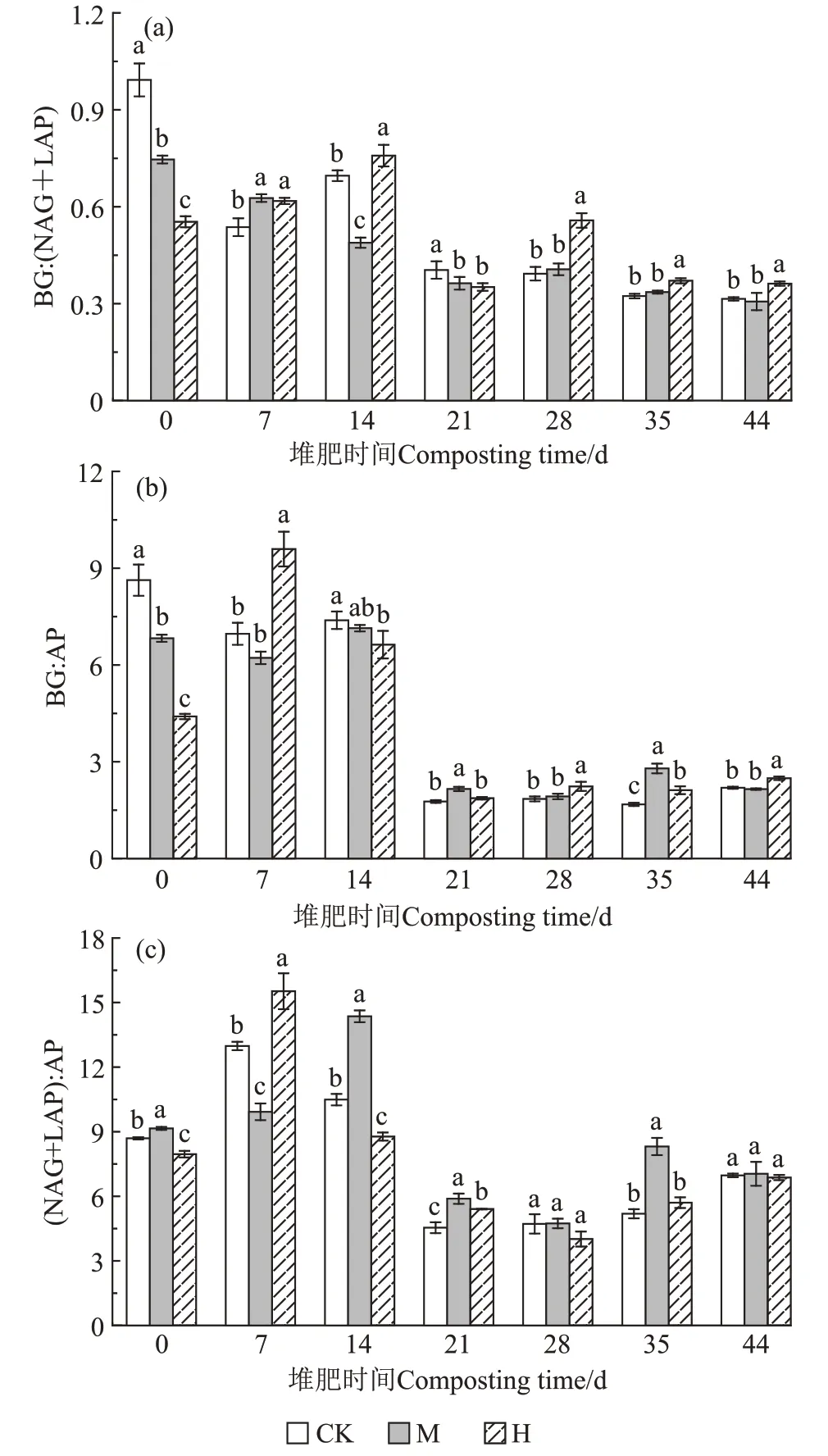
圖7 酶化學計量在不同時期的變化Figure 7 Enzyme stoichiometry in different periods
2.5 腐植酸組分與理化性質、胞外酶活性之間的關系
堆肥理化性質、化學計量、胞外酶活性(藍色箭頭)和腐植酸組分(紅色箭頭)的冗余分析(RDA)結果如圖8所示:HA-C與AP、NAG、LAP呈正相關,與TN、TP、N∶P、pH呈正相關,與FA-C、BG、DOC、TOC、C∶N、C∶P、T、EC、-N、-N呈負相關。

圖8 堆肥過程中基本理化性質與腐植酸碳組分的冗余分析Figure 8 Redundancy analysis of basic physicochemical properties and carbon components of humic acid in composting process
根據RDA 相關性分析結果構建的結構方程模型(SEMs)如圖9 所示,可以看出3 組化學計量影響相關胞外酶活性進而影響HA-C 的形成,3 個模型的擬合指數已經滿足完美擬合指標的標準(X2/df<2,P>0.05,GFI>0.950,CFI>0.900 以及RMSEA<0.05)[24]。3 組影響HA-C 形成的關鍵胞外酶不同:在CK 中,C∶N 調節BG 和AP的活性,從而影響HA-C 的生成,C∶N、BG 和AP 都與HA-C 的生成呈負相關;在M 中,隨著堆肥的進行,C∶N 逐漸降低,LAP 活性增強,促進HA-C 的生成;同理,在H 中,隨C∶N 的降低,AP 活性增強,進而促進了HA-C的生成。

圖9 化學計量和胞外酶對HA-C形成的影響Figure 9 Effects of stoichiometry and extracellular enzymes on the formation of HA-C
3 討論
3.1 添加菇渣和高溫預處理對葡萄枝堆肥腐殖化的影響
本研究3 個處理的堆肥在堆制45 d 后都達到了無害化衛生標準要求,且添加菇渣和高溫預處理促進了葡萄枝堆肥胡敏酸的生成。在堆肥過程中,糖類和氨基酸等腐殖質前體可被微生物降解和轉化成腐植酸,當可利用的有機物不足時,不穩定的腐植酸會被微生物降解,以提供微生物所需的能量[25]。本研究堆肥化的第7~14 天,溫度較高,CK 的腐植酸碳含量陡降,下降速率為35.26%,而M和H只下降了14.89%和11.57%,且腐植酸中較穩定的組分胡敏酸開始升高,上升速率較CK 快,說明在M 和H 中一些有機碳更多地是轉化為胡敏酸而不是被降解,從而提高了堆肥的腐植酸含量。高溫預處理可促進堆體中多糖與類蛋白反應產生類腐殖質物質,羧基、羥基和芳核等腐殖質物質也會增多[26],因此,高溫預處理初始和最終的腐植酸含量都較高。菇渣為芳構化和氧化反應提供了豐富的底物,底物中含有大量的纖維結構成分,而且菇渣本身含有較多的腐植酸[27],因此添加菇渣的堆肥中最終HA-C 含量最多。堆肥化第7 天,M 和H 的TN 就已經高于CK,說明添加菇渣和高溫預處理可以減少堆肥中的N 損失,加速TN 的濃縮效應。菇渣中的易降解有機物較多,其快速分解導致較多短鏈脂肪酸累積[28],從而可中和一部分NH+4-N,減少N 素損失,這與Meng 等[9]在污泥堆肥中添加菇渣的研究結果一致。高溫預處理會抑制反硝化細菌生長以及產生有機酸降低物料pH 而起到保N 作用[29]。研究表明,部分腐植酸的形成是通過含N 和含P 化合物與其他腐殖質前體的縮合實現的[30-31]。本試驗中,TN、TP 與HA-C 的含量都是隨著堆肥的進行逐漸增加,三者之間存在正相關關系(圖8),說明一些有機物分解產生的N、P離子以與其他前體螯合的形式被回收,并存在于HA-C 中。TN、TP 和HA-C 的含量同時有效提升,與Jia 等[25]在菇渣堆肥中添加園林廢棄物的研究結果一致。這也是添加菇渣和高溫預處理可促進胡敏酸生成的原因之一。
3.2 添加菇渣和高溫預處理對胞外酶活性的影響
胞外酶是微生物釋放的可以降解復雜大分子的蛋白質,參與養分循環并具有重要作用,其活性強弱可以表示養分轉化能力的大小[32]。添加菇渣在堆肥初期可顯著提升4 種胞外酶的活性。香菇菇渣富含降解纖維素及木質素的真菌[4],可增加堆肥中有機物的含量,為真菌、細菌等微生物的生長提供了更多更易利用的碳源和氮源,從而使堆肥初期酶活性較高。M 的NAG 酶活性在堆肥初始達到峰值,可能是由于菇渣中富含真菌菌絲,微生物殘余積累的肽聚糖豐富,死亡的真菌菌絲體通過微生物演替而分解[33]。由于90 ℃的高溫預處理,H 的AP、BG、LAP 活性在堆肥初期顯著低于其他兩組(P<0.05),可能是因為超高溫堆肥衍生出的芳烴比例較高,且酚基碳的強度更高[34],而酚類物質的濃度增加會導致和養分循環相關的微生物活性的降低[12]。有機物的分解速率可以用胞外酶活力來表征[32],各組高溫時期只有BG 和LAP的活性沒有因為高溫而明顯下降,可能由于此時微生物大量繁殖,纖維素、蛋白質等碳源、氮源快速分解,微生物的同化吸收更需要這兩種酶參與,H 的4 種胞外酶活性在高溫期和腐熟期都顯著高于CK(P<0.05),表明高溫預處理有助于大分子有機物的降解。各組4 種酶的活性都是在第28 天開始下降并逐漸穩定,原因是在升溫和高溫階段,易代謝底物較多,微生物大量繁殖,大分子有機物迅速分解;而在后期,大部分容易利用的有機物已被降解完全,堆體穩定,微生物活性減弱,酶活性降低。此外,酶和腐殖質之間形成的復合物也破壞了酶與底物的相互作用,導致在堆肥后期胞外酶活性降低[12]。
3.3 堆肥養分和胞外酶活性的化學計量與HA-C 生成的關系
化學計量能夠有效說明養分限制類型,是反映C、N、P 循環以及養分平衡的重要參數[35]。堆肥C∶N是體現堆肥有機物分解速率的指標,比值越高表明有機物的礦化分解速率越慢[16]。本研究中,從第7 天開始到堆肥結束,H 的C∶N 比其他兩組都低,尤其在高溫期迅速下降,下降速率相比CK 高247.41%,也是在同一時期,H 的HA-C 含量較CK 顯著上升,表明高溫預處理有利于有機物降解礦化和胡敏酸的快速形成。C∶P 也能在一定程度上反映有機質的礦化速率,通常C∶P 越低,越利于微生物對有機物的分解,同時提高堆肥全P 的含量[36]。與C∶N 相同,H 的C∶P 下降速率快,也顯示出高溫預處理有利于有機物礦化。第14天之前,M 和H 的C∶P 高于CK,第14 天之后,H 的C∶P開始低于CK,第44天,M和H的C∶P都低于CK。整個堆肥期間,M 和H 的N∶P 一直高于CK,M 在第7 天到第14 天的C∶P 顯著高于其他兩組(P<0.05),結合相應時期M 的BG∶AP 較小以及N∶P 較大,說明M 組微生物受環境影響需要分泌更多的AP來滿足生長需求,在此期間M 中的微生物受到P養分的限制。研究證明,微生物會最大限度地優化自己的儲備資源,易利用的底物是提升微生物生物量和分泌胞外酶的基礎[37]。堆肥過程中,當可用的能量和營養物質變得有限時,微生物就會釋放該元素的獲取酶,以應對養分供應的變化,從而改變生態酶的化學計量特征[37]。SEMs 顯示(圖9),在整個過程中,各組的HA-C 含量受不同胞外酶的影響:CK 的HA-C 含量與BG 和AP都呈顯著負相關(P<0.01)。當BG 酶分泌較多時,纖維素和木質素逐漸被分解,而它們的類衍生物是構成腐植酸的核心骨架和主要成分[17],因此不利于胡敏酸聚合。M 的HA-C 含量與LAP 呈顯著正相關(P<0.05),在H 中HA-C 含量和AP 呈顯著正相關(P<0.01)。即在M 和H 中產生的微生物N 和P 限制會提高蛋白質、有機磷降解酶的活性,導致N 和P 礦化增加,以應對N、P 限制,礦化后的物質有利于合成胡敏酸:如NH+4-N 可通過非生物途徑作為前體直接參與胡敏酸的形成[38];P 和腐植酸、胡敏酸之間可以螯合,螯合產物還能促進P 的溶解度和生物利用率[31]。從圖7 可以看出,BG∶(NAG+LAP)、BG∶AP、(NAG+LAP)∶AP的值都是在第21天之前變化較大,與CK相比此時M 和H 中的微生物受到的養分限制較大,第21 天之后比值減小并穩定,也是因為前期有機質降解,組分變化快,難以降解的大分子有機物需要微生物分泌相應的胞外酶來降解。
4 結論
(1)葡萄枝雞糞好氧堆肥高溫期(>50 ℃)持續10 d 以上,種子發芽指數均達到70%以上,滿足堆肥無害化和腐熟標準。添加菇渣(M)和高溫預處理(H)改變了堆肥基質的特性,加速了有機質降解,提升了養分含量,促進了腐殖化。堆肥第44 天,M 和H 的TN提升率相比CK 提高了40.84 個和50.41 個百分點,TP含量提升率相比CK 提高了41.38 個和71.40 個百分點、TOC 含量的降解率分別比CK 提高了1.94 個和1.13個百分點;相比CK,H 的腐殖化率提升了9.56%,M的腐殖化指數提升了22.11%。
(2)添加菇渣和高溫預處理影響了參與C、N、P循環功能的酶的活性和化學計量,顯著提升了4 種胞外酶的活性:M 組在第0~7 天胞外酶活性較高,H 組在第14~44 天胞外酶活性較高,以應對N 和P 養分的限制,降解更多的大分子有機物,提升N 和P 的含量,有利于胡敏酸的生成,進而提升了胡敏酸的含量。
