茄子果形性狀遺傳研究
2023-05-17 02:44:28房桂萍成玉富
蔬菜 2023年5期
關鍵詞:模型
房桂萍,成玉富,徐 強
(揚州大學園藝與植物保護學院,江蘇 揚州 225000)
茄子(Solanum melongenaL.)的品種類型豐富多樣,這些品種資源為茄子新品種選育與雜種優勢的利用提供了良好的基礎。我國的茄子種植和消費的區域性較強,且消費者對茄子外觀品質的要求也存在區域性,選育符合區域消費者喜好的商品果形是茄子育種的目標之一。同時,茄子果形也是評價外觀品質的重要指標。
植物遺傳改良研究過程中形成2種主要方法:一類是世代平均法,由Mather于1949年提出;另一類是混合模型,由Stewart和Elston在1973年首次提出,為“一個主基因和多基因”的遺傳模型,在此基礎上Maclean和Morton在1974年進一步提出混合模型,同時,也解決了模型應用中的參數估計問題。隨后,在植物數量性狀分析模型中,Elkind等[1]提出“主基因+多基因”的遺傳分析模型,該模型不僅可以估算出單基因的遺傳效應值與多基因的遺傳效應值,而且可以估算出兩者間的互作效應。這些模型雖然應用了主基因與多基因的混合遺傳模型,但主要著眼于對主基因的分析,對多基因的遺傳效應還不夠重視,遺傳分析時可以選擇的遺傳模型不多,遺傳分析群體僅包括F2等比較單一的遺傳群體,試驗精確度不足。
綜合前人的研究基礎,蓋鈞鎰等[2]、王建康等[3]將數量遺傳學與混合分布理論相結合,分析過程中將整個群體作為分析對象,建立了一套系統的植物數量性狀遺傳分析模型,即主基因+多基因遺傳分析模型。該模型的建立與發展更好地實現了將統計學與遺傳學的統一[4-6]。本試驗采用主基因+多基因遺傳分析模型對茄子果形性狀進行遺傳分析,以期為茄子育種提供指導。
1 材料和方法
1.1 試驗材料
試驗所用3個親本材料均為揚州大學園藝與植物保護學院蔬菜遺傳育種實驗室提供的高代純合自交系茄19-37、茄19-38和茄19-39,茄19-37為白果、白肉、長筒果;茄19-38為綠果、綠肉、高圓果;茄19-39為白果、白肉、短筒果。
1.2 試驗方法
2020年將3個親本材料配制成2個雜交組合(組合Ⅰ茄19-37×茄19-38,組合Ⅱ茄19-38×茄19-39),2021年進行回交、自交得到2個組合的6個世代群體:P1、P2、F1、F2、B1、B2,2022年3月將2個組合的6個世代群體定植到大田中,其中親本P1和P2各種植30株,雜交一代F1各種植40株,F2(F1自交得到)、B1(F1×P1得到)和B2(F1×P2得到)各種植120株,共種植920株,雙行定植,行距60 cm,株距45 cm,田間常規管理。
1.3 果形類型描述
每株采收發育正常的商品成熟果,直尺測量果實縱徑,從果柄基部到果頂為縱徑,游標卡尺測量果實最大橫切為橫徑,計算果形指數,果形指數=果實縱徑/果實橫徑。根據茄子種質資源描述規范對試驗中涉及的果實類型進行描述(表1)[7]。
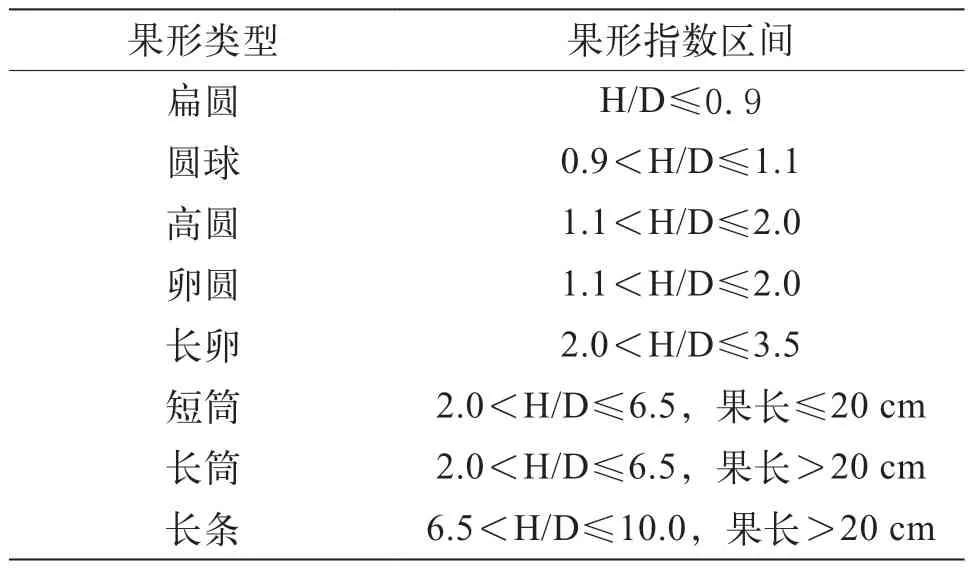
表1 茄子果形分類標準
1.4 統計分析方法
根據蓋鈞鎰等[2]對主基因-多基因遺傳體系分離分析方法的介紹,主要步驟如下:(1)分離世代的理論分布,在二倍體核遺傳、不存在母體效應、主基因和多基因間不存在互作和連鎖(也可能存在連鎖)、配子和合子均無選擇的假定下,每一主基因由于多基因和環境的修飾呈正態分布,整個分離世代的表現分布為多個主基因型正態分布的混合分布。不分離世代P1、P2、F1等的表現為其基因型由于環境修飾而呈正態分布;(2)建立5類遺傳模型,每類中按在基因加性效應基礎上有無顯性效應分類:正向顯性、負向顯性、完全顯性或者部分顯性,2對主基因時還要考慮有無上位性效應、有無連鎖等各種遺傳模型,各類模型見表2;(3)按照采用的試驗群體類型及世代建立各類別遺傳模型的混合分布函數,并由之建立各模型的對數極大似然函數;(4)由各試驗群體及世代的觀察次數分布通過IECM(interated EMC)算法從各模型的對數極大似然函數中估計出相應的各世代、各成分分布的參數(包括成分分布的平均數、方差及該成分分布所占的比例等)以及相應的似然數值;(5)通過各模型的似然函數值計算出AIC(Akaike's information criterion)值,AIC值最小的模型為相對最佳模型,有可能存在多個相對最佳模型。同時進行一組樣本分布與模型所代表的理論分布間的合適性測驗,包括均勻性檢驗(U12、U22、U32)、Smirnov檢驗(nW2)和Kolmogorov檢驗(Dn),綜合各檢驗結果,結合各群體的實際分布情況,選擇相對最佳的遺傳模型;(6)由最佳遺傳模型的各成分分布參數估計出主基因和多基因的有關遺傳參數,從而估計各種遺傳方差。

表2 遺傳模型代號和含義
利用主基因+多基因多個世代聯合分析的方法[8],采用曹錫文等[9]的植物數量性狀分離分析Windows軟件包SEA在R語言環境下對茄子6個世代的果形指數進行數據分析。
2 結果與分析
2.1 親本及雜交后代的果形分布
2個組合的親本及雜交后代群體的果形指數統計如表3,組合Ⅰ中,長筒果形母本37與高圓果形父本38雜交,F1果形為短筒形,果形指數均值為2.26,介于2個親本數值之間,接近于低值親本;F2分離世代的果形指數為1.11~7.35,除了長筒形與高圓形2個親本類型外,還出現了卵圓、長卵、短筒和長筒等果形類型,并出現了超親類型;B1分離世代的果形指數為1.38~9.38,也出現了超親類型;B2分離世代的果形指數為0.81~2.96,沒有出現超親類型;F2、B1、B2這3個分離群體的變異系數均超過了20%,說明變異幅度較大。組合Ⅱ中,高圓果形母本38與短筒果形父本39雜交,F1果形為短筒形,果形指數均值為3.11,介于2個親本數值之間,接近于低值親本;F2分離世代的果形指數為2.26~4.99,且出現超親類型;B1分離世代的果形指數為2.23~4.52,B2分離世代的果形指數介于1.86~4.97,均出現了超親類型;F2、B1、B2這3個分離群體變異系數為15%~20%,變異幅度較大。
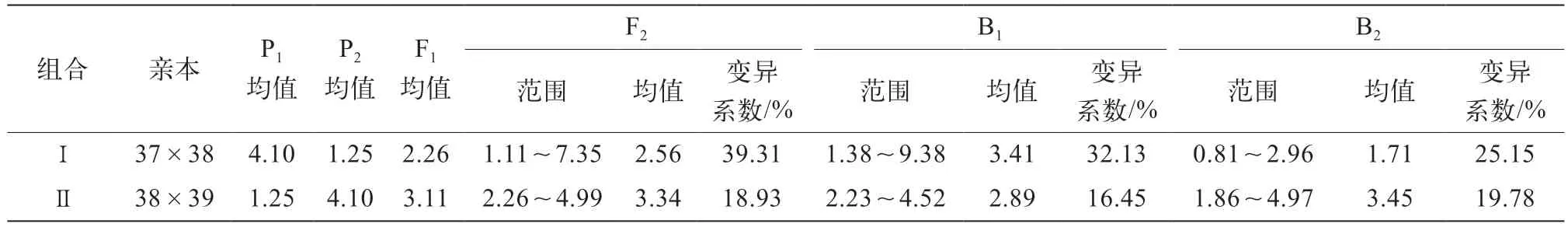
表3 2個組合茄子親本及F1、F2、B1和B2代果形指數均值統計
2.2 最優模型的確定
利用極大對數似然法計算得到2個組合6個世代果形指數的24種遺傳模型的極大對數似然值和AIC值(表4)。根據AIC值最小原則,組合Ⅰ中E-0模型AIC值最小,為515.092 8,可能為最佳模型,其次為E-3模型(AIC值為518.890 8),與之相近,挑選為候選模型。同樣,組合Ⅱ中E-3模型AIC值最小,為777.965 1,可能為最佳模型;B-1模型的AIC值為778.237 8,與E-3相近,選為候選模型。
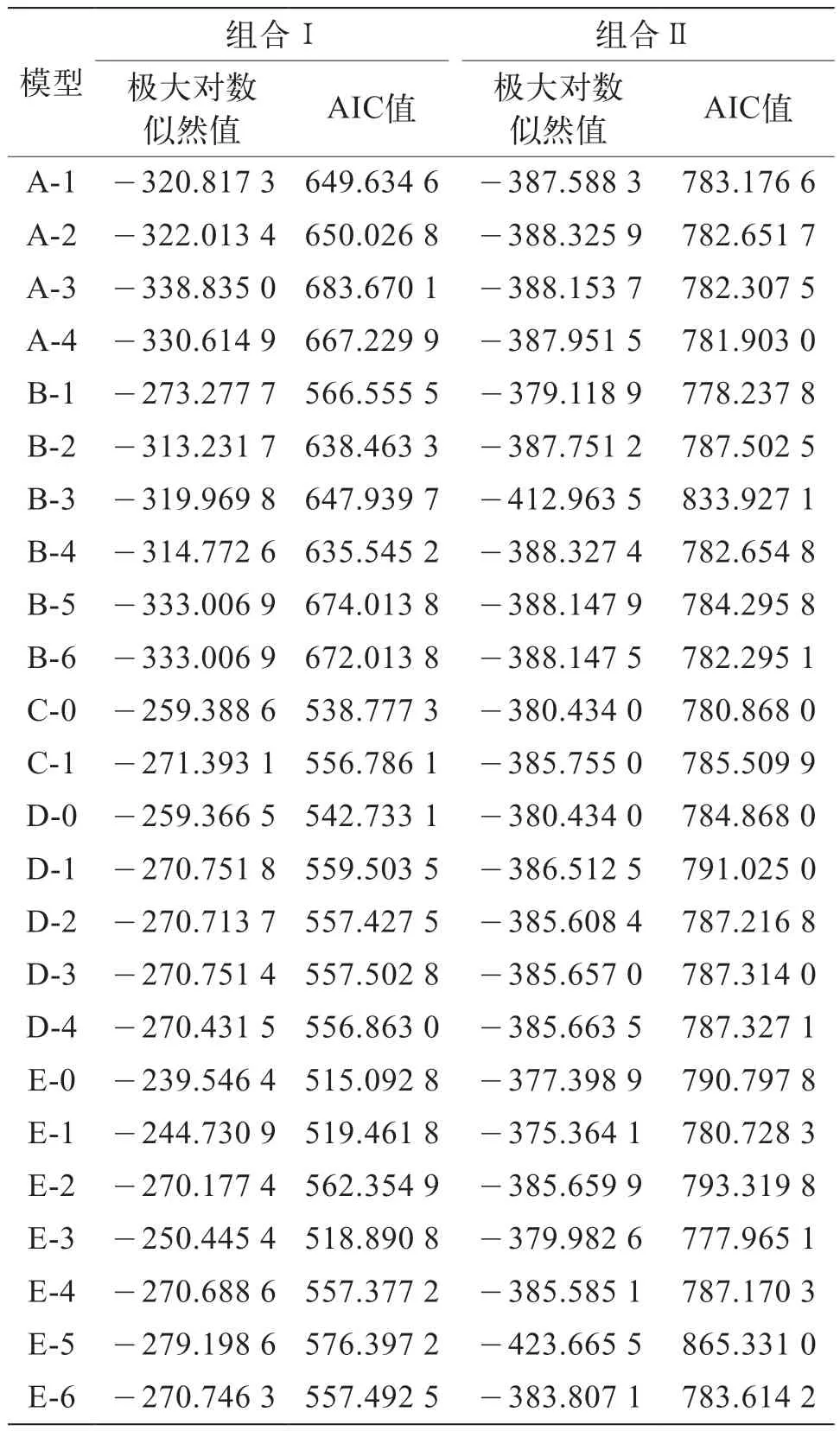
表4 2個茄子組合果形指數遺傳模型的極大對數似然值和AIC值
對2組模型進行適合性檢驗(表5、表6),通過均勻性檢驗(U12、U22、U32)、Smirnov檢驗(nW2)及Kolmogorov(Dn)檢驗,選擇最優遺傳模型發現,組合Ⅰ中E-0模型有2個參數出現顯著水平,E-3模型有1個參數出現顯著性差異,根據備選模型適合性檢驗中選擇統計量達到顯著水平個數最少這一原則,選擇E-3模型為最適模型。組合Ⅱ中B-1和E-3模型均有1個參數出現顯著性差異,則選擇2個備選模型中AIC值最小(最小值為777.965 1)的模型為最適模型,即選擇E-3模型為組合Ⅱ的最佳模型。
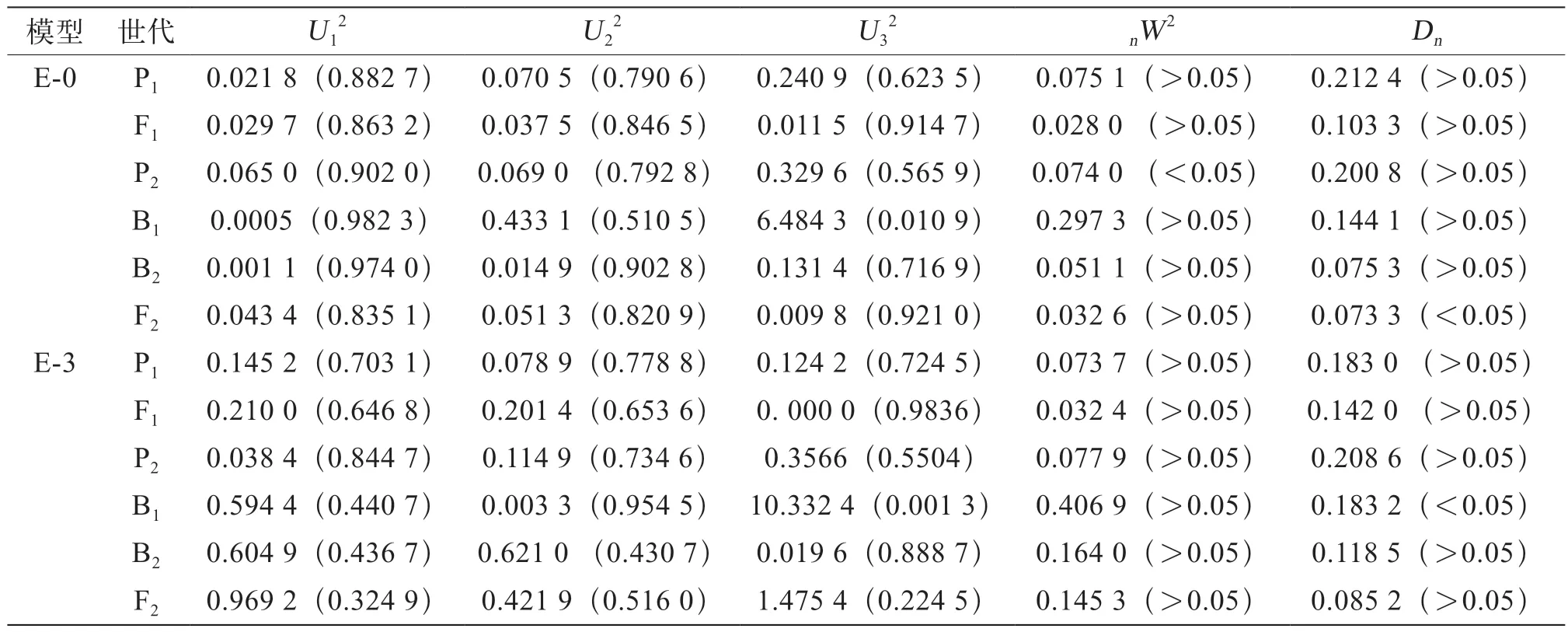
表5 茄子組合Ⅰ遺傳模型的適合性檢驗結果
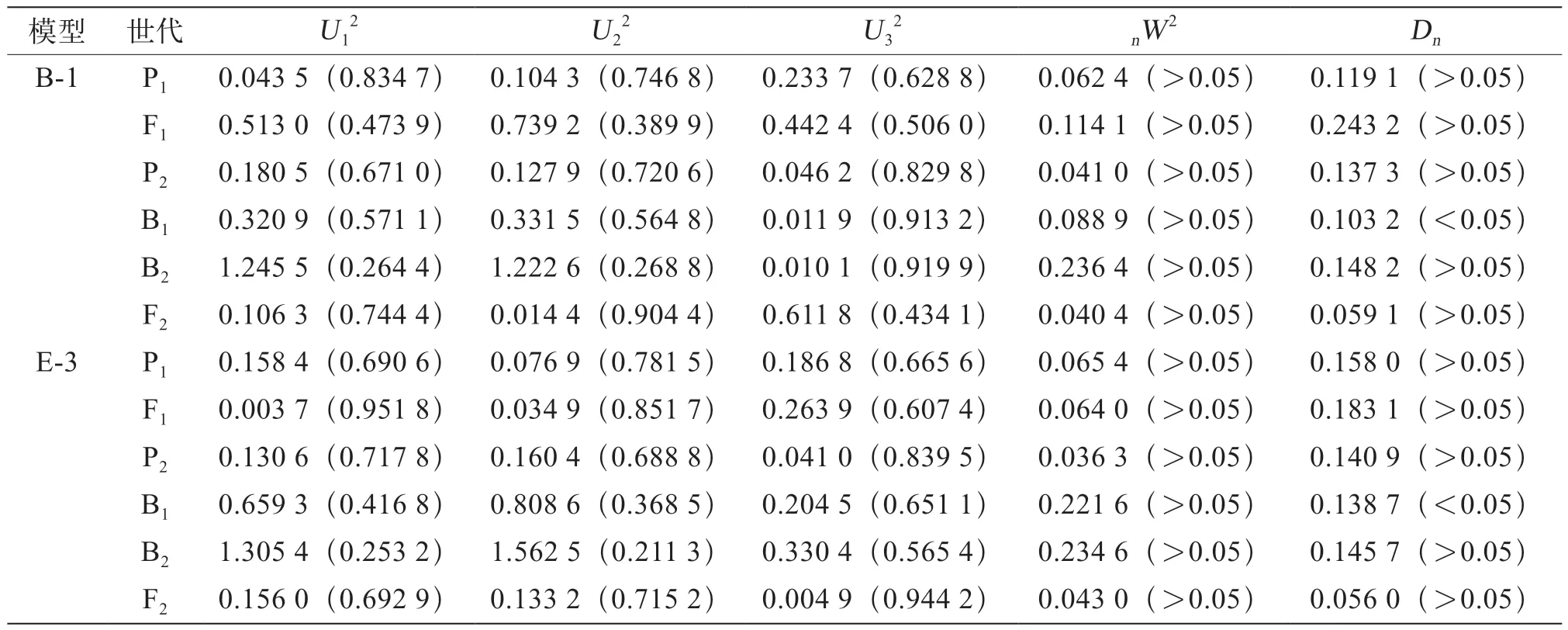
表6 茄子組合Ⅱ遺傳模型的適合性檢驗結果
2.3 最優模型的遺傳參數估計
利用最小二乘法對組合Ⅰ的E-3模型和組合Ⅱ的E-3模型的成分分布參數進行估算,得到了2個模型的一階和二階遺傳參數(表7、表8)。通過對最優遺傳模型的一階和二階遺傳參數分析可知(表7),組合Ⅰ的第一主基因加性效應為負向的-0.041 3,第二主基因加性效應為0.302 4,多基因的加性效應為1.164 4,多基因的顯性效應為0.219 0,多基因的加性效應大于顯性效應;組合Ⅰ的B1、B2、F2分離世代的主基因遺傳率(hmg2)分別為51.010 4%,40.153 5%、38.436 1%,多基因遺傳率(hpg2)分別為39.980 1%、31.356 5%、51.762 8%,環境效率(1-hmg+pg2)分別為9.009 5%、28.490 0%、9.801 1%,B1、B2世代主基因遺傳率大于多基因遺傳率,以主基因遺傳為主,F2世代的多基因遺傳率大于主基因遺傳率,且F2世代的多基因遺傳率最高,B2世代遺傳受環境影響較大。
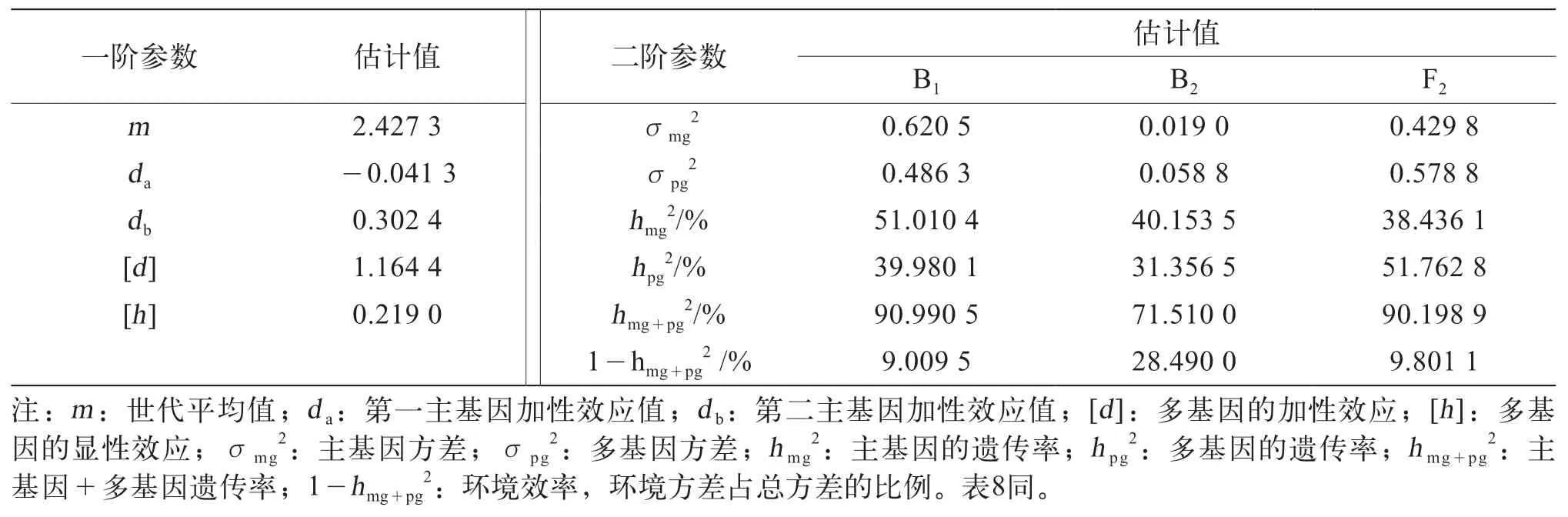
表7 茄子組合Ⅰ果形性狀最優模型遺傳參數估計
由組合Ⅱ的一階和二階遺傳參數可知(表8),第一主基因和第二主基因加性效應分別為負向的-0.021 9和-0.845 6,多基因的加性效應為0.864 1,多基因的顯性效應為0.333 6,多基因的加性效應大于顯性效應;組合Ⅱ的B1、B2、F2分離世代的主基因遺傳率(hmg2)分別為33.470 2%、30.789 1%、33.935 0%,主基因遺傳率F2最高,其次為B1;多基因遺傳率(hpg2)分別為22.138 0%,27.583 9%、41.169 7%,F2世代的多基因遺傳率最高;環境效率(1-hmg+pg2)分別為44.391 8%、41.627 0%、24.895 3%,B1和B2世代遺傳受環境影響較大。
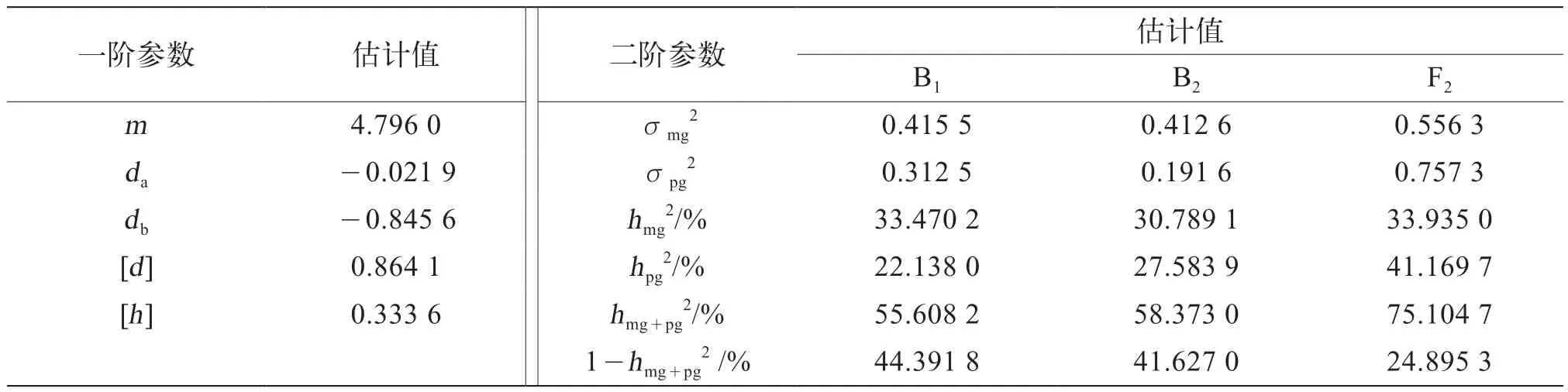
表8 茄子組合Ⅱ果形性狀最優模型遺傳參數估計
3 結論與討論
本研究結果表明,茄子果形指數遺傳屬于數量性狀,2個組合果形指數遺傳模型均適于E-3模型,即2對加性主基因+加性-顯性多基因模型,組合Ⅰ表現出一負一正的主基因加性效應,組合Ⅱ表現出2個負向的主基因加性效應,2個組合多基因的加性效應均大于顯性效應,說明以加性效應遺傳為主;組合Ⅰ中B1、B2世代主基因遺傳率均大于多基因遺傳率,以主基因遺傳為主;F2世代多基因遺傳率大于主基因遺傳率,以多基因遺傳為主;B2世代環境效率較高,為28.490 0%。組合Ⅱ中B1、B2分離世代的主基因遺傳率均大于多基因遺傳率,以主基因遺傳為主;F2的多基因遺傳率大于主基因遺傳率,以多基因遺傳為主;B1和B2世代遺傳受環境因素影響較大,環境效率高于40%。
潘玉朋等[10]認為采用主基因與多基因聯合分析方法對果形遺傳的研究,適用于瓜果類園藝作物果形數量遺傳性狀的遺傳研究。成穎等[11]采用圓形櫻桃番茄自交系與梨形自交系構建6個世代群體,分析其遺傳規律發現,果形遺傳模型符合E-5模型(2對完全顯性主基因+加性-顯性多基因模型),與本試驗遺傳模型略有差異。陳婕英[12]以近圓形冬瓜自交系與長圓柱形冬瓜自交系構建4個世代群體,分別分析了果實橫徑、果實縱徑和果形指數3個性狀的遺傳規律,其中果形指數遺傳模型符合E-0模型,即2對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型,和本研究模型稍有不同。喬軍等[13]選用扁圓、長筒和短筒3個果形差異明顯的茄子高代自交系構建2個雜交組合6個世代群體,以果形指數和三角果形指數分析茄子果形遺傳規律,發現2個組合均適用于D-2模型,即1對加性主基因+加性-顯性多基因模型,與本研究的模型不同,但2個組合遺傳都以加性遺傳為主,分析可能是由于材料的不同和環境的差異導致遺傳模型發生變化。
多個研究表明,茄子果形遺傳受加性效應的影響,在育種工作中,需注意對親本果實形狀的選擇,同時環境因素也會在遺傳過程對果形基因的表現有累積作用,在選種時應注意。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
