油茶根際土壤真菌群落組成及多樣性分析
2023-05-17 06:47:24羅鑫吳躍開張念念許杰楊再華
中國農業科技導報 2023年2期
羅鑫, 吳躍開, 張念念, 許杰, 楊再華
(貴州省林業科學研究院,貴陽 550005)
根際土壤微生物(rhizosphere soil microorganism)種類繁多,是根際微生態系統(rhizosphere microecosystem)的重要組成部分[1]。其中,根際土壤真菌(rhizosphere soil fungi)對植物攝取營養物質、促進生長發育、提高宿主植物抗病性以及維持根際微生態系統平衡等起重要調節作用[2-3]。在根際微生態系統中,土壤理化性質的差異會影響根際土壤真菌群落的組成和數量,且在不同植物種類下兩者關系復雜多樣。吳佳偉等[4]研究表明灌木林、草坡、濕地3種植被的根際土壤真菌多樣性與土壤全氮、全鉀和速效鉀含量呈顯著相關。魏世清等[5]研究指出油茶林根際土壤真菌數量與速效鉀含量呈顯著正相關,與有機質、有效磷、速效氮呈極顯著正相關。可見,植物根際土壤真菌多樣性及數量與土壤環境因子存在一定的相關性,但其真菌群落組成與土壤環境因子的關系尚不明確。
油 茶(Camellia oleiferaAbel.)為 山 茶 科(Theaceae)山茶屬(Camellia)的常綠小喬木,是我國南方特有的純天然高級優質油料。油茶中提取的茶籽油具有很高的營養價值,在工業、旅游文化、生態和食品等方面具有不可估量的經濟價值[6]。油茶在我國已有2 300年的栽植歷史,其中,貴州以得天獨厚的山區環境成為我國主要的油茶產出省份,油茶種植面積達17.56×104hm2,在天柱、玉屏、黎平、望謨和冊亨等多地區擁有代表性的油茶產業基地[7]。目前,貴州油茶種植已具備一定規模,但近年來多處油茶林頻發茶餅病、炭疽病、軟腐病等病害,致使油茶品質和產量下降,一定程度上影響了貴州油茶產業。因此,急需通過對油茶林生長環境進行探究,明確土壤微生物對油茶生長和病害的影響。
目前,油茶的相關研究主要集中在植物病原菌的鑒定和活性物質的提取,有關根際土壤微生物多樣性的研究雖已有少許[8-10],但其與土壤環境因子的相關性研究卻鮮見報道。因此,本研究通過高通量測序,對貴州不同地區油茶根際土壤真菌的群落組成及多樣性進行分析,并進一步探究油茶根際土壤真菌組成與土壤環境因子的關系,以期為開發有益的油茶根際微生物資源、調控土壤環境因子預防油茶病害提供科學依據。
1 材料與方法
1.1 油茶根際土壤的采集
于2020 年9 月4—10 日,分別在貴州省貴陽(Guiyang,GY)、天柱(Tianzhu,TZ)、玉屏(Yuping,YP)、望謨(Wangmo,WM)、威寧(Weining,WN)、黎平(Liping,LP)、冊亨(Ceheng,CH)7個地區采集油茶根際土壤樣品,采樣地信息詳見表1。每個采樣點隨機選擇4 棵長勢相近且健康的6 年生油茶樹,每棵樹之間距離大于20 m。去除表面枯葉及雜質,沿油茶樹主根垂直地面向下挖取至油茶根系,再以主根為中心向外挖取至35~40 cm 細根發達的根系,在東南西北四周均收集地面以下10—20 cm 深度的植物細根土壤共500 g,裝入無菌塑封袋中充分混勻,迅速置于冰盒中保存并帶回實驗室。將采集到的土壤樣品分成2份,其中1份置于-80 ℃超低溫冰箱中保存,用于提取土壤真菌DNA;另1 份土壤置于常溫下陰干后,一部分過0.25 mm孔徑篩用于測定土壤pH及有機質、全氮、全磷和全鉀含量,另一部分過2 mm孔徑篩的土壤用于測定堿解氮、有效磷、速效鉀含量。

表1 油茶根際土壤采樣地點及信息Table 1 Sampling sites and information of C. oleifera rhizosphere soil
1.2 土壤理化性質測定
根據鮑士旦[11]的方法測定土壤理化性質。土壤pH 采用點位測定法;有機質(organic matter,OM)含量采用油浴加熱重鉻酸鉀氧化法;全氮(total nitrogen,TN)含量采用堿解擴散法+半微量開氏法;全磷(total phosphorus,TP)含量采用消煮液+鉬銻抗比色法;全鉀(total potassium,TK)含量采用消煮液+ 火焰光度法;堿解氮(alkali hydrolyzed nitrogen,AN)含量采用堿解擴散法+半微量開氏法;有效磷(available phosphorus,AP)含量采用碳酸氫鈉浸提+鉬銻抗比色法;速效鉀(available potassium,AK)含量采用乙酸銨浸提+火焰光度法。每個樣品重復4次。
1.3 土壤真菌群落組成及多樣性分析
1.3.1 DNA 提取與測序 根據CTAB 法[12]對土壤樣品進行總DNA 的提取。利用引物ITS1F(5'-CTTGGTCATTAGAGAGAGTAA-3')和ITS2R(5'-GCTGCGTTCTTCTCTCGATGGC-3')進行ITS 序列擴增。PCR(ABI GeneAmp?9700 型)擴增采用TaKaRarTaqDNA 聚合酶和20 μL 反應體系:10×Buffer 4 μL,dNTPs(2.5 mmol·L-1)2 μL,正向引物ITS1F(5 μmol·L-1)0.8 μL,反 向 引 物ITS2R(5 μmol·L-1)0.8 μL,rTaq聚 合 酶0.2 μL,BSA 0.2 μL,模板DNA 10 ng,隨后添加ddH2O至20 μL。PCR 程序:95 ℃ 180 s;98 ℃ 30 s,55 ℃ 30 s,72 ℃45 s,35個循環;72 ℃ 10 min,4 ℃保存。PCR 產物經上海美吉生物醫藥科技有限公司Illumina Hiseq 平臺進行高通量測序,而后于I-Sanger 生物信息分析云平臺(http://www.i-sanger.com/)進行數據處理和根際土壤真菌多樣性分析。
1.3.2 序列處理與組成分析 利用FLASH 將所有原始序列數據依次進行過濾、拼接、剔除嵌合體后獲得有效序列。通過平臺QIIME(http://qiime.org/scripts/assign_taxonomy.html)將識 別 閾 值在97% 以上的序列聚類為操作分類單元(operational taxonomic units,OTU)[13]。從每個OTU中選擇最豐富的序列作為該OTU 的代表性序列,計算出每個OTU 在各個樣品中的相對豐度(relative abundance),并在Unite(http://unite.ut.ee/index.php)真菌數據庫進行物種注釋,獲得每個OTU 在各級(界、門、綱、目、科、屬、種)分類水平信息[14]。所有樣本的數據按最小序列數41 857抽平后,再進行后續多樣性分析。采用單因素方差分析(one-way,ANOVA)比較不同地區間豐度分布存在差異的物種,多重比較采用Tukeykramer 檢驗差異性顯著水平。真菌組成以及物種差異均使用R 3.5.0 軟件進行繪制。
1.3.3 多樣性分析 Alpha 多樣性分析采用Mothur(http://www.mothur.org/wiki/Schloss SOP#Alpha diversity)指數分析程序包計算各樣本的Shannon指數、Simpson指數、ACE指數和Chao 1指數,并用t檢驗進行差異顯著性檢驗[15]。Beta多樣性分析通過非加權組平均法(unweighted pair-group method with arithmetic,UPGMA)聚類樣品OTU,基于unweighted unifrac距離算法下對樣品在門水平上進行聚類,并獲得層級聚類樹。通過非度量多維尺度(nonmetric multi-dimensional scale,NMDS)分析檢驗真菌在屬水平群落組成的差異,同時利用相似性分析(analysis of similarities,ANOSIM)檢驗不同地區間樣本差異是否顯著大于同一地區內樣本差異[16]。
1.4 數據分析
采用SPSS 20.0 軟件進行數據的統計分析。利用R 軟件的冗余分析(redundancy analysis,RDA)研究油茶根際土壤真菌相對豐度前10 的類群、土壤環境因子以及土壤樣本三者間的關系,并利用envfit 函數檢驗土壤環境因子間關系的顯著性;同時將根際土壤真菌群落豐度與土壤環境因子進行相關性分析。
2 結果與分析
2.1 油茶根際土壤的理化性質
7 個地區油茶根際土壤的理化性質如表2 所示,各地區土壤均呈酸性,pH 4.35~4.79。依據全國第二次土壤普查養分分級標準[17],威寧地區土壤有機質、全氮、全磷含量均達到Ⅰ級,為極豐富水平;黎平地區土壤中有機質、全氮、全磷和堿解氮含量達到Ⅳ或Ⅴ級,呈適宜至缺乏之間水平。土壤中全鉀含量最高是玉屏地區,處于Ⅲ級最適水平;其次為冊亨和望謨地區。有效磷和速效鉀含量最高的分別是貴陽和天柱地區,但貴陽地區土壤的有效磷含量僅為Ⅳ級(適宜水平),而天柱地區速效鉀含量則處于Ⅱ級(豐富水平)。由此表明,7 個地區間油茶根際土壤具有不同的理化性質特征。
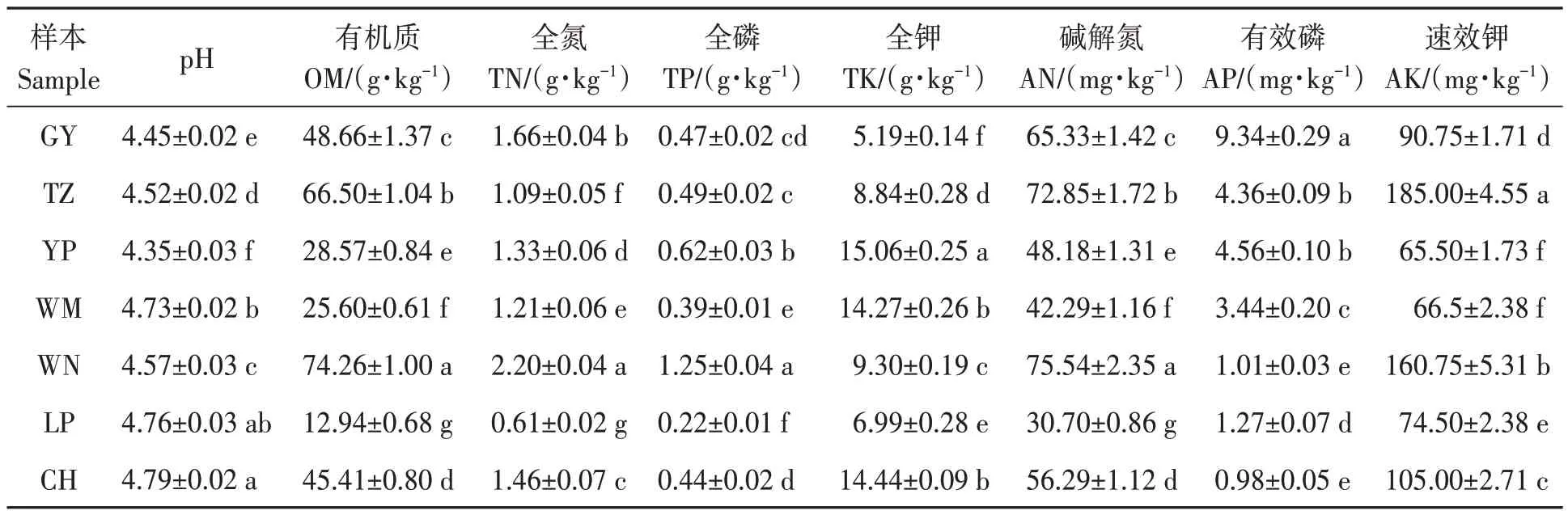
表2 不同地區油茶根際土壤的理化性質Table 2 Physicochemical properties of C. oleifera rhizosphere soil from different regions
2.2 測序結果與Alpha多樣性分析
經Illumina Miseq 高通量測序后,7 個地區的土樣中共獲得1 959 963 條有效ITS 序列(表3)。基于97%相似度水平下,共聚成了634 個OTUs。覆蓋度達99.90%~99.97%,表明測序結果基本覆蓋了樣本中絕大多數真菌的生物學信息,能夠準確反映油茶根際土壤真菌群落的結構組成及多樣性。對于多樣性指數(Shannon 指數和Simpson 指數),7 個地區的多樣性從大到小依次為望謨>冊亨>威寧>天柱>黎平>玉屏>貴陽;對于豐富度指數(ACE 指數和Chao 1 指數),從大到小依次為冊亨>天柱>望謨>貴陽>威寧>黎平>玉屏。由此表明,7 個地區的油茶根際土壤真菌群落多樣性和豐富度存在差異。
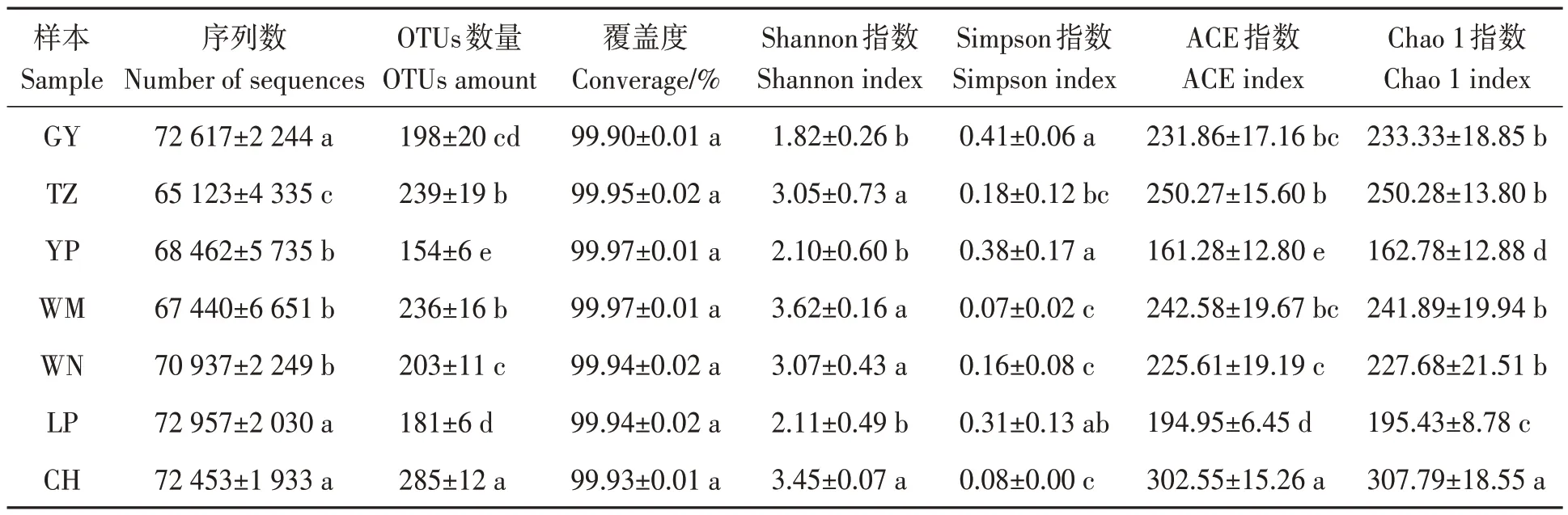
表3 油茶根際土壤真菌ITS測序結果及Alpha多樣性Table 3 ITS sequencing results and alpha diversity of rhizosphere soil fungi of C. oleifera
2.3 土壤真菌的Beta多樣性分析
使用韋恩圖分析7 個地區間共有和特有的OTUs,結果(圖1)表明,7 個地區共有的OTUs 為42 個,為各地區特有的OTUs 數量從多到少依次為望謨>冊亨>威寧>天柱>貴陽>玉屏>黎平,各地區特有的OTU 數量分別占其總OTU 數量的56.70%、41.67%、40.85%、31.15%、27.59%、12.05%、10.64%。基于門水平的UPGMA 聚類樹顯示(圖2),天柱與玉屏地區的根際土壤真菌群落組成較相似;威寧、貴陽和望謨3個地區的根際土壤真菌群落結構較相似;而黎平地區與其他地區的土壤真菌組成差異較大。
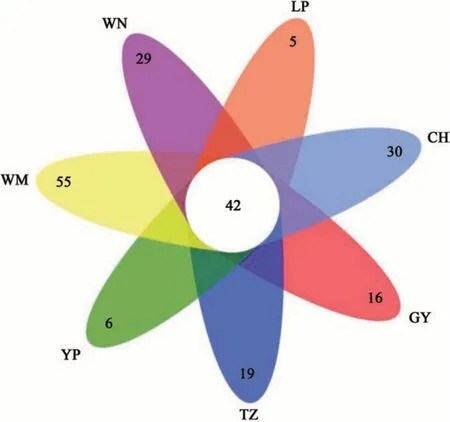
圖1 OTUs數量韋恩圖Fig. 1 Venn profile of OTU number
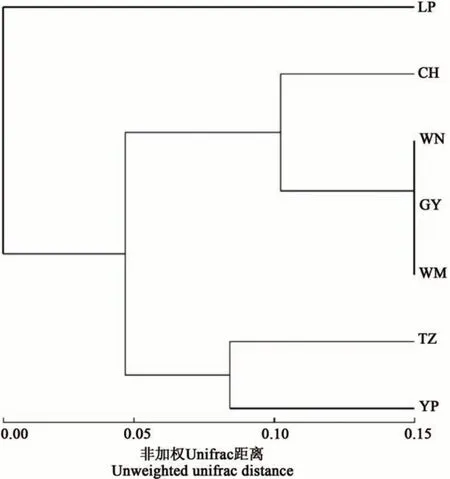
圖2 油茶根際土壤真菌在門水平上的UPGMA聚類樹Fig. 2 UPGMA cluster tree of fungi in the rhizosphere soil of C. oleifera at phylum level
基于unweighted unifrac 距離在屬水平上的NMDS 分析(圖3)表明,7 個地區的油茶根際土壤真菌群落組成存在顯著差異(P=0.001),即不同地區的土壤樣品具有分組意義。且ANOSIM(圖4)表明,組間的距離顯著大于各地區的土壤樣本。綜上所述,各地區間油茶根際土壤真菌群落組成差異顯著。
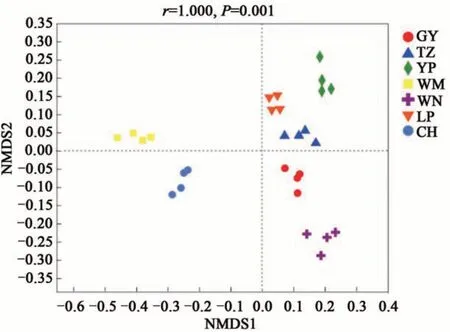
圖3 基于非加權unifrac距離算法下油茶根際土壤真菌在屬水平上的NMDS分析Fig. 3 NMDS analysis of rhizosphere soil fungi form C.oleifera based on unweighted unifrac algorithm
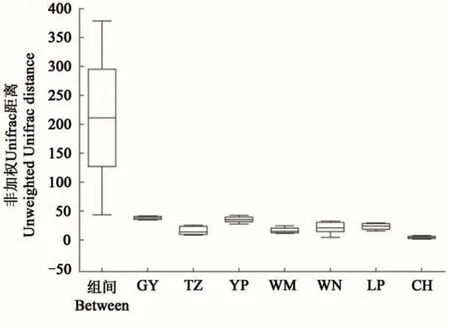
圖4 ANOSIM分析距離盒狀圖Fig. 4 Anosim analysis distance box diagram
2.4 土壤真菌的群落組成及物種分布差異分析
7 個地區的油茶根際土壤真菌測序所獲得的634個OTUs隸屬于9門、32綱、73目、141科、213屬。在門水平上,包含子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)、被 孢 霉 門(Mortierellomycota)、Rozellomycota、球囊菌門(Glomeromycota)、毛霉門(Mucoromycota) 、壺 菌 門(Chytridiomycota) 、Zoopagomycota 和Calcarisporiellomycota(圖5)。玉屏地區根際土壤真菌優勢門為擔子菌門(61.71%);而其余6個地區根際土壤真菌的優勢菌門均為子囊菌門(54.00%~83.62%)。不同地區在門水平上的組成和相對豐度存在差異,其中,子囊菌門在貴陽地區油茶根際土壤中的相對豐度較高,為83.62%;擔子菌門在玉屏地區油茶根際土壤中的相對豐度較高,為61.71%;被孢霉門在天柱地區油茶根際土壤中的相對豐度較高,為24.02%;Rozellomycota在黎平地區油茶根際土壤中的相對豐度較高,為7.67%。
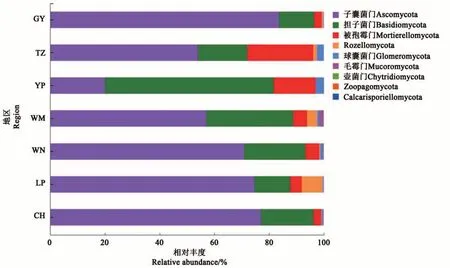
圖5 油茶根際土壤真菌在門水平上的組成及相對豐度Fig. 5 Composition and relative abundance of fungi in rhizosphere soil of C. oleifera at phylum level
在屬水平上,除未能分類到屬的真菌類群外,7 個地區均檢測到沙蜥屬(Saitozyma)、Archaeorhizomyces、被 孢 霉 屬(Moretierella)、Clavaria、青 霉 屬(Penicillium)、木 霉 屬(Trichoderma)、籃狀菌屬(Talaromyces)、瓶霉屬(Cladophialophora)、枝孢菌屬(Cladosporium)和Geminidasidium(圖6)。同時,不同地區間油茶根際土壤真菌的組成和相對豐度也存在差異,貴陽、天柱、玉屏和望謨地區根際土壤真菌的優勢菌屬為沙蜥屬,其相對豐度依次為11.18%、14.75%、59.11%和17.61%;而威寧地區的優勢菌屬為Archaeorhizomyces,相對豐度為38.13%;冊亨地區的優勢菌屬為Trichoglossum,相對豐度為10.26%。此外,Trichoglossum和Cuphophuyllus是冊亨地區油茶根際土壤中的特有菌屬,相對豐度分別為10.26%和8.49%;Ilyonectria為望謨地區的特有菌屬,相對豐度為2.65%;Sebacina和Tomentella為威寧地區的特有菌屬,相對豐度分別 為 1.85% 和 1.60%;Coniosporium和Metschnikowia為天柱地區的特有菌屬,相對豐度分別為1.03%和1.00%;而貴陽和黎平地區未發現特有菌屬。

圖6 油茶根際土壤真菌在屬水平上的組成及相對豐度Fig. 6 Composition and relative abundance of fungi in rhizosphere soil of C. oleifera at genus level
分析在各地區樣本間存在顯著分布差異的真菌菌屬,結果(圖7)表明,豐度排前10的菌屬分別為沙蜥屬、Archaeorhizomyces、被孢霉屬、青霉屬、Clavaria、木霉屬、籃狀菌屬、瓶霉屬、Oidiodendron和Ilyonectria。其中,沙蜥屬在玉屏地區根際土壤中的相對豐度顯著高于其他地區;Archaeorhizomyces、瓶霉屬和Oidiodendron在威寧地區土壤中的相對豐度顯著高于其他地區;被孢霉屬、木霉屬和籃狀菌屬在天柱地區根際土壤中的相對豐度顯著高于其他地區,而青霉屬、Clavaria和Ilyonectria在望謨地區土壤中的相對豐度顯著高于其他地區。
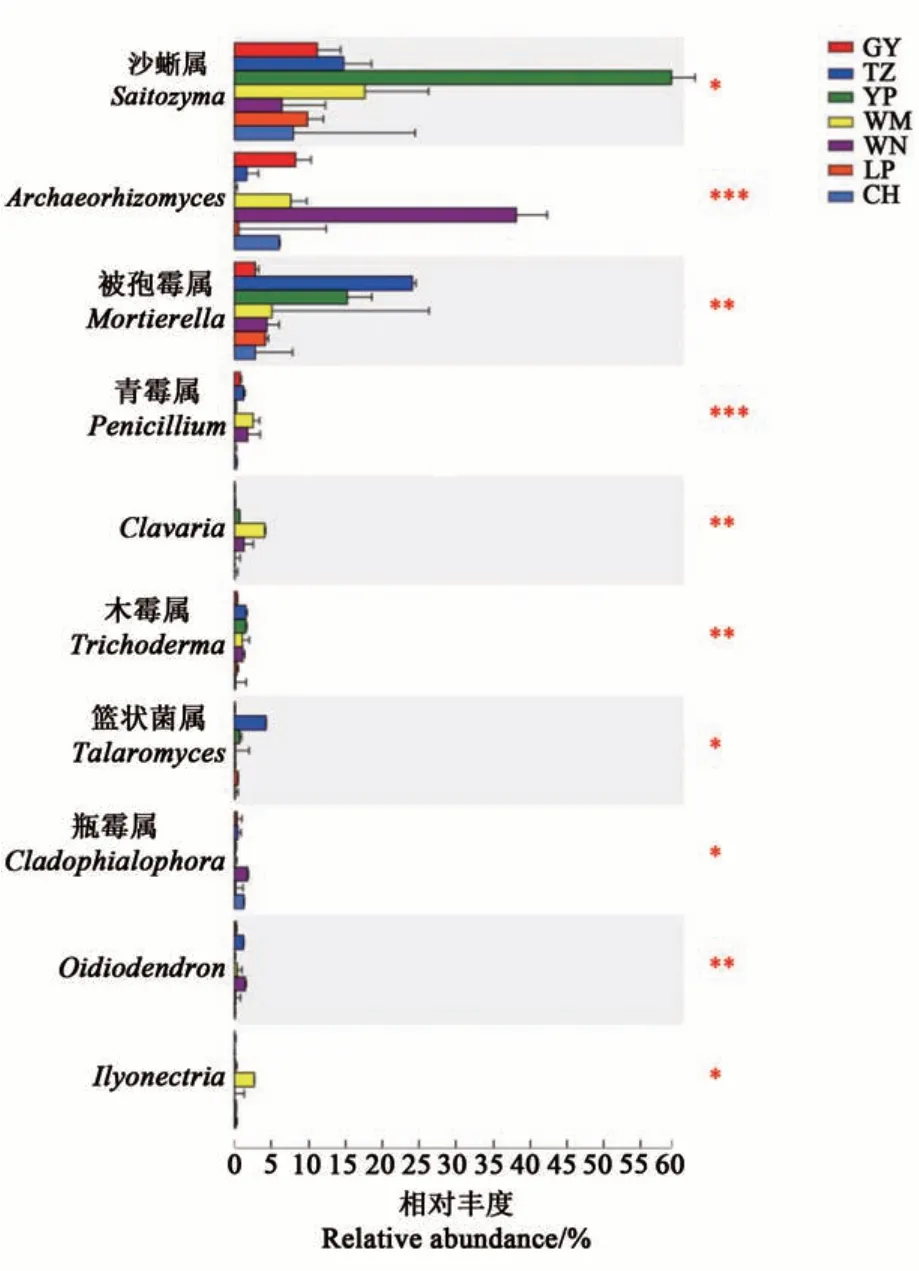
圖7 油茶根際土壤真菌的物種差異分析Fig. 7 Differential species analysis of fungi in the rhizosphere soil of C. oleifera
2.5 土壤真菌與土壤環境因子的相關性分析
對油茶根際土壤樣品中相對豐度排前10 位的真菌菌屬與土壤環境因子進行RDA 分析,結果(圖8)顯示,第1、第2軸的解釋率分別為44.08%和22.21%,累計總解釋率達66.29%,表明該結果較好地反映了真菌群落與土壤環境因子之間的相互關系。土壤環境因子對油茶根際土壤真菌群落的影響從大到小依次為:全鉀>有效磷>速效鉀>有機質>全磷>堿解氮>全氮>pH。Cuphophyllus、Trichoglossum、Archaeorhizomyces等菌屬均與土壤全磷、全氮、堿解氮、有機質、速效鉀含量及土壤pH 呈正相關,與土壤有效磷含量呈負相關;而被孢霉屬和沙蜥屬與土壤有效磷含量呈正相關,與土壤全磷、全氮、堿解氮、有機質、速效鉀含量及土壤pH 呈負相關。沙蜥屬、被孢霉屬、Cuphophyllus和Trichoglossum與土壤全鉀含量呈正相關;而Archaeorhizomyces與土壤全鉀含量相關不顯著。經顯著性檢驗(表4)表明,土壤全鉀、有效磷、速效鉀和有機質含量對真菌群落結構影響顯著。由此表明,土壤全鉀、有效磷、速效鉀和有機質含量是影響油茶根際土壤真菌群落組成的主要因子。
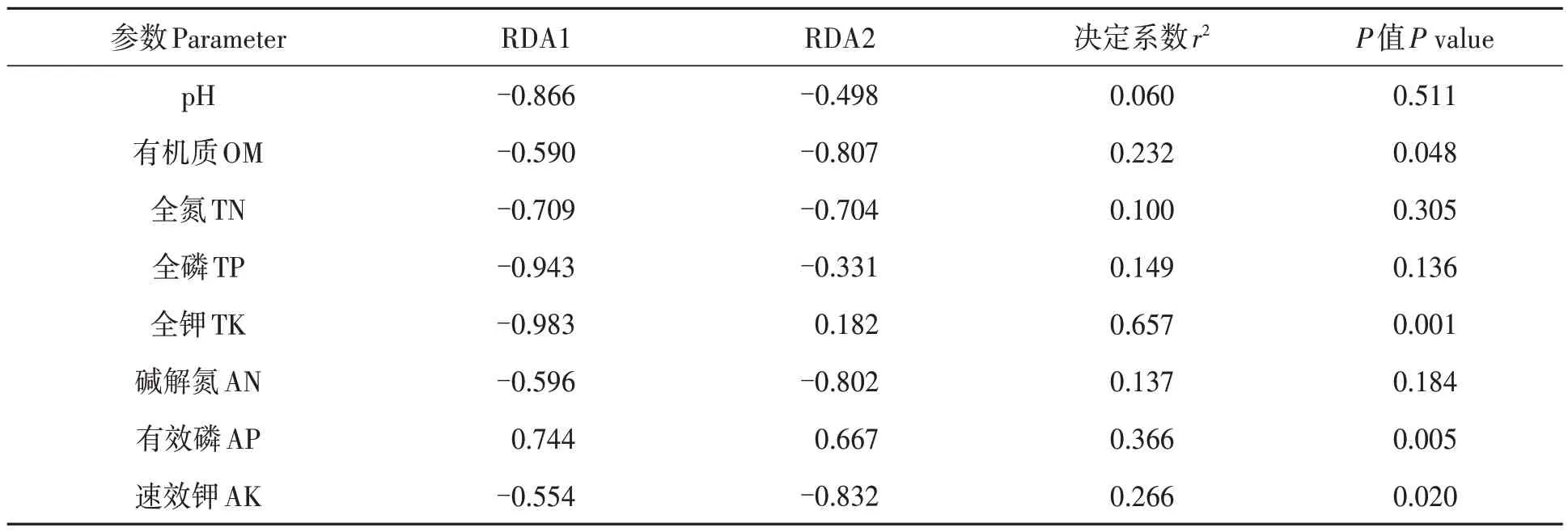
表4 前10優勢真菌類群與土壤環境因子envfit函數檢驗Table 4 Envfit permutation function test of top ten dominant fungal groups and soil environmental factors
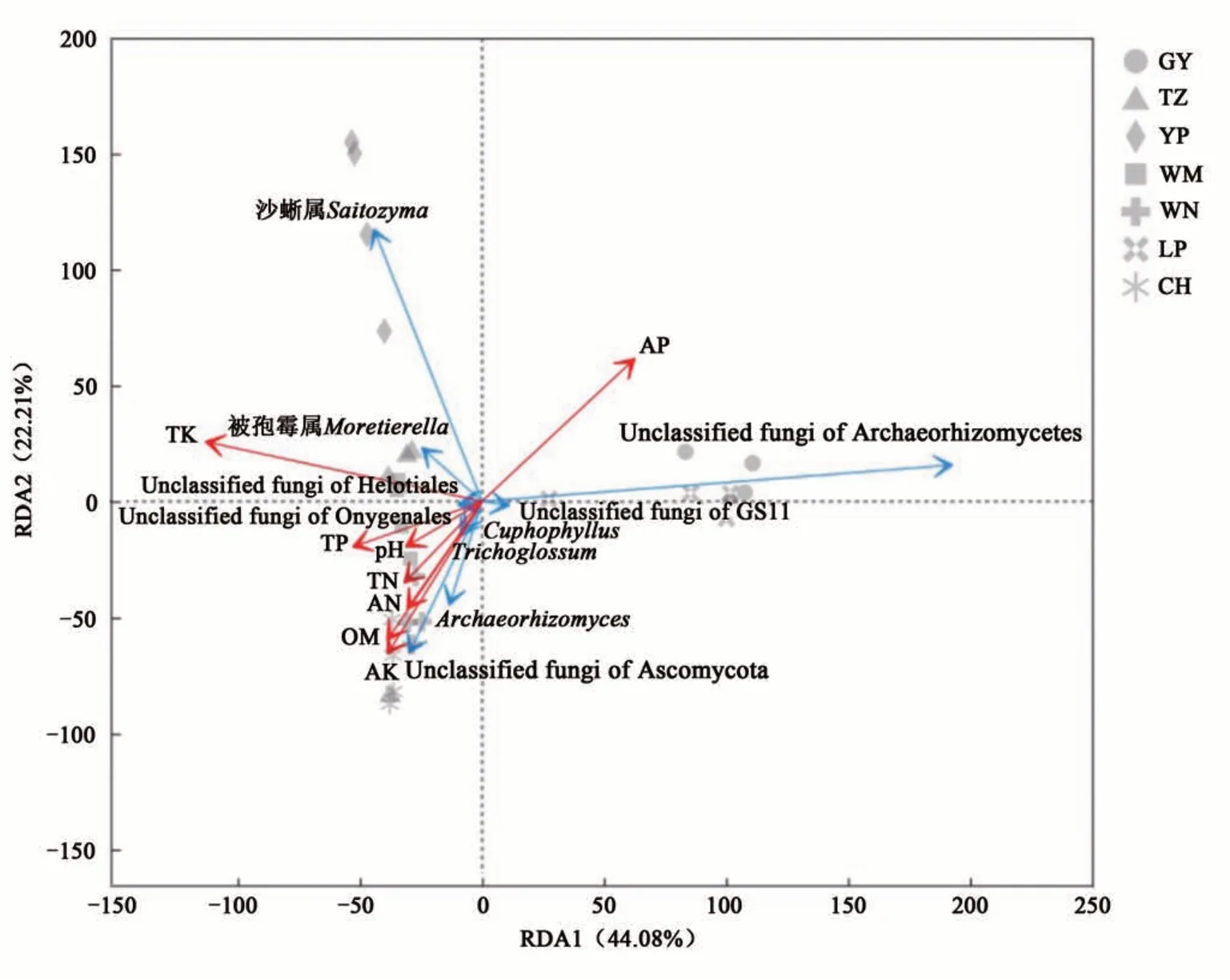
圖8 前10優勢真菌類群與土壤環境因子在屬水平上的RDA分析Fig. 8 RDA analysis of top ten dominant fungal groupsand soil environmental factors in C. oleifera rhizosphere soil at genus level
Spearman 相關分析(圖9)顯示,Ilyonectria與土壤pH呈極顯著正相關,與土壤有機質、全磷和堿解氮含量呈極顯著負相關;Archaeorhizomyces與土壤全氮含量呈極顯著正相關;籃狀菌屬和假黑孢霉(Coniosporium)與土壤全氮含量呈極顯著負相關;鐮刀菌屬(Fusarium)與土壤pH 呈顯著正相關,與有機質、全氮、全磷和堿解氮含量呈顯著負相關;沙蜥屬與土壤有效磷含量呈顯著正相關;Cuphophyllus和Cladophialophora與土壤有效磷含量呈顯著負相關;Cladophialophora和Oidiodendron與土壤速效鉀含量呈極顯著正相關;沙蜥屬、Ilyonectria和與土壤速效鉀含量呈顯著負相關。
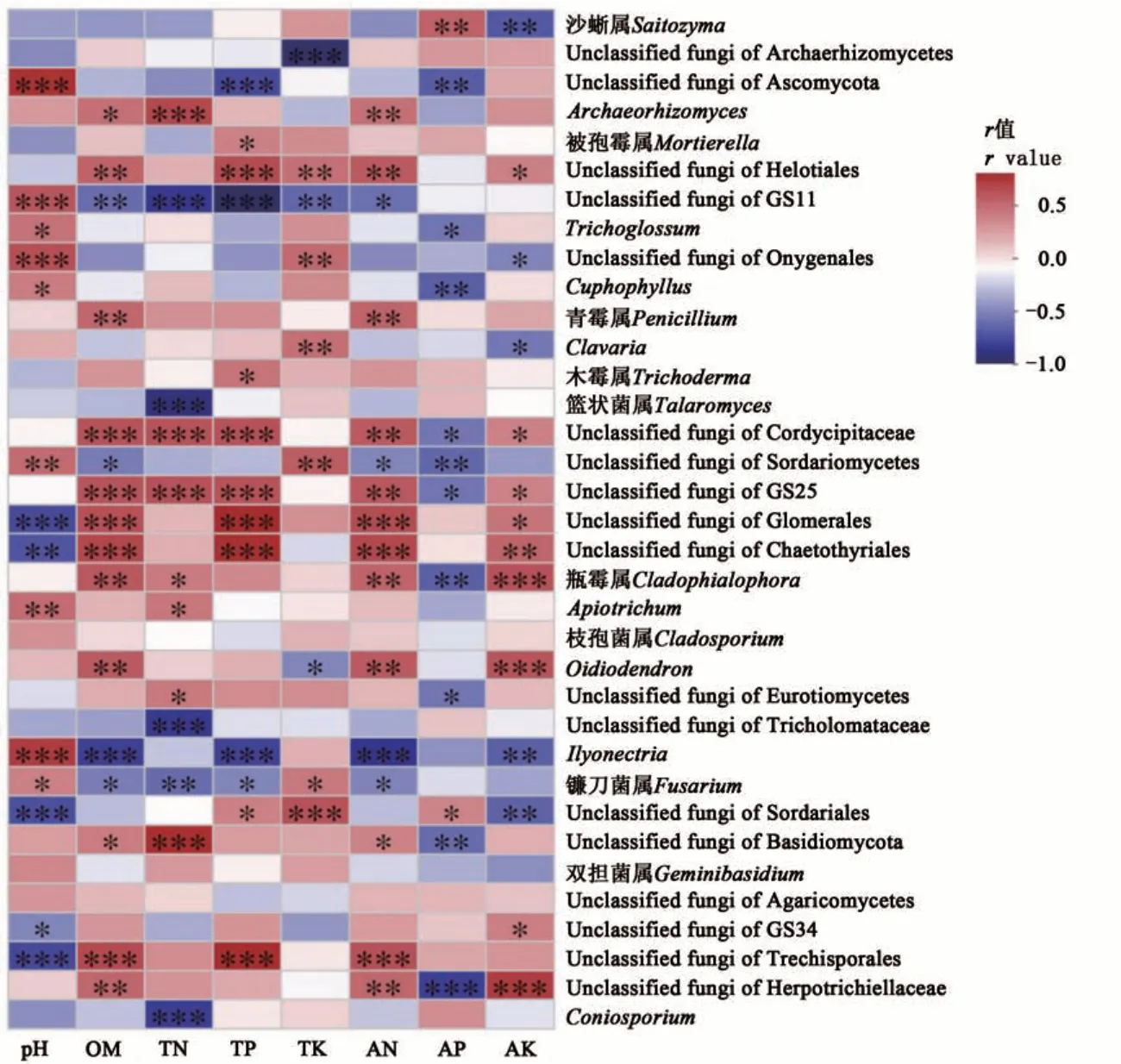
圖9 屬水平物種豐度與土壤環境因子的Spearman相關性分析Fig. 9 Spearman correlation analysis of species abundance and soil environmental factors at genus level
3 討論
油茶喜好有機質和養分豐富的酸性(pH 5.0~6.5 )土壤,但也具有一定的耐干旱瘠薄特性[18]。本研究中,威寧、天柱、貴陽和冊亨4 個地區油茶土壤的有機質含量達Ⅰ級;玉屏和望謨地區油茶土壤有機質含量達Ⅲ級;黎平地區則處于Ⅳ級。總體上看,各個地區土壤的有機質含量豐富,且呈酸性,這與油茶的生長習性相吻合。另外,7 個地區的油茶均栽植于丘陵或者斜坡地帶,其中,天柱和冊亨兩地的健康油茶長勢優越,這2個地區分別位于貴州的東南和西南,與馬帥兵等[19]提出的貴州油茶的最適宜栽培區主要分布在東部和南部地區相一致。7 個地區土壤的堿解氮含量均處于Ⅳ級和Ⅴ級(適宜但偏缺乏水平),有效磷含量多處于Ⅴ級至Ⅵ級之間(缺乏水平),速效鉀含量均處于Ⅱ級到Ⅳ級之間(豐富水平),表明油茶根際土壤呈現少氮、低磷、富鉀的狀態。而7個地區間土壤的全氮、全磷和全鉀含量存在顯著差異,這可能與不同地區的立地土壤條件不同有關。
根際土壤真菌具有降解有機質、釋放營養和微量元素及調控土壤養分的作用,在促進植物生長過程中扮演著重要角色[20-21]。本研究中7 個地區油茶根際土壤真菌均具有豐富的多樣性,但真菌多樣性與群落組成具有差異。研究表明,根際土壤真菌的組成及多樣性受植物種類和土壤環境等多種因素影響[22-23]。本研究表明,子囊菌門和擔子菌門在油茶根際土壤中的相對豐度較高,可能是由于子囊菌門可分解土壤中腐爛的動植物遺骸,釋放有機物供植物吸收利用;而擔子菌門可與植物共生形成菌根,有利于植物生長發育[24]。在屬水平組成上,沙蜥屬、Archaeorhizomyces、被孢霉屬和Clavaria等普遍存在于7 個地區的油茶根際土壤中,與傅海平等[25]的研究結果有部分相似,而與其他物種植物差異較大[26-27]。在不同地區間表現出顯著差異的菌屬中,木霉屬具有降解木質素的功能,而青霉屬可產生有機酸代謝產物溶解土壤中的無機磷,從而調節土壤肥力,間接影響植物生長[28]。由此表明不同地區油茶根際土壤理化性質的差異一定程度上可能受到功能性土壤真菌分布差異的影響。另外,被孢霉屬可與根際微生物互作,在植物代謝、繁殖和抗逆性中發揮重要作用,是影響油茶生長和品質的重要真菌[29]。可見,油茶根際土壤真菌的分布差異可能是土壤環境與微生物間互作的結果[30]。然而,土壤環境與土壤中微生物的互作對影響根際土壤微生物的組成分布是否具有協同效應還需要進一步研究。
土壤是根際土壤真菌賴以生存的環境,其環境因子與根際土壤真菌有著密切聯系。土壤環境因子制約著根際土壤真菌群落的種類和數量,而環境因子的變化又受到根際土壤真菌種類和數量的影響[31-32]。本研究顯示,油茶根際土壤的全鉀、有效磷、速效鉀和有機質含量是影響根際真菌群落組成的主要因子,說明這些土壤因子對于7 個地區土壤真菌的群落組成和豐度起著重要作用。Spearman 相關性分析表明,Archaeorhizomyces、青霉屬與土壤堿解氮含量呈顯著正相關;沙蜥屬、被孢霉屬與有效磷含量呈正相關;Cladophialophora、Oidiodendron與速效鉀含量呈極顯著正相關。由此表明,可通過施用相關的微生物氮肥、磷肥、鉀肥提高土壤養分,促進油茶生長[33]。在本研究中,沙蜥屬和被孢霉屬是油茶根際土壤中豐度較高的優勢菌屬,但各地區土壤中有效磷含量并不高,推測可能是由于在植物根系吸收的同時部分土壤微生物在增殖和定植于植物根系過程中也吸收了土壤中大量的磷元素[34]。研究表明,Ilyonectria可引發多種植物患病,鐮刀菌屬也是土壤中常見的致病菌[35-36]。對于缺乏有機質以及氮磷鉀的土壤,植物由于養分獲取不足,抗性下降,為土壤中病原菌的侵害提供了機會[37]。本研究發現Ilyonectria與土壤pH 呈極顯著正相關,與土壤有機質、全磷和堿解氮含量呈極顯著負相關;鐮刀菌屬與土壤pH 呈顯著正相關,與有機質、全氮、全磷和堿解氮含量呈顯著負相關。這表明Ilyonectria和鐮刀菌屬在油茶根際土壤真菌群落中的豐度變化具有潛在的致病風險。因此,及時關注土壤pH 變化、調控土壤中有機質及氮磷鉀含量可成為預防和監控油茶病害的有效途徑。本研究結果為油茶根際土壤真菌與土壤環境因子相關性研究提供了新的結論,并為開發微生物菌肥資源和調控土壤環境因子預防植物病害奠定了科學研究基礎。
