海南東寨港兩種紅樹植物旱季和雨季的光合生理特征比較
2023-05-22 22:25:47鄭熊葉文偉趙從舉梅敏華
廣西植物 2023年4期
鄭熊 葉文偉 趙從舉 梅敏華

摘 要:植物光合生理生態特性是退化植物群落恢復、重建植物種選擇的重要依據。為研究不同紅樹植物光合生理生態特性,該研究于2021年旱季的4—5月、雨季的7—9月利用LI-6400光合儀,測定紅樹植物秋茄和海蓮的光合生理參數和主要生態因子,并采用通徑分析方法分析主要生態因子對凈光合速率的影響。結果表明:(1)秋茄旱季凈光合速率日均值(8.43 μmol-2·s-1)略低于雨季(8.67 μmol-2·s-1),差異不顯著;海蓮旱季凈光合速率日均值(7.03 μmol-2·s-1)顯著低于雨季(9.41 μmol-2·s-1);旱季秋茄凈光合速率日均值顯著高于海蓮,而雨季秋茄凈光合速率日均值顯著低于海蓮。(2)旱季、雨季秋茄蒸騰速率、氣孔導度、胞間CO2濃度等光合生理因子日均值變化幅度小于海蓮,水分利用效率也低于海蓮。(3)旱季、雨季兩種紅樹植物均存在“光合午休”現象。旱季,秋茄屬于非氣孔限制,而海蓮屬于氣孔限制;雨季,秋茄和海蓮均屬于氣孔限制。(4)旱季影響秋茄凈光合速率的主要決策因子是光合有效輻射,主要限制因子是相對濕度,而生態因子對海蓮凈光合速率均起限制作用,其中飽和水汽壓差是主要限制因子;雨季秋茄和海蓮的主要決策因子均是光合有效輻射,主要限制因子均是飽和水汽壓差。綜上研究表明,秋茄對于生境變化有著更強的適應性,海蓮不僅具有較好耐旱性能,而且更適合高溫多雨生境,該研究結果為退化紅樹林的恢復重建植物種選擇提供了科學依據。
關鍵詞: 凈光合速率, 季節變化, 紅樹植物, 通徑分析, 海南東寨港
中圖分類號:Q945
文獻標識碼:A
文章編號:1000-3142(2023)04-0649-09
Abstract:The photosynthetic physiological and ecological characteristics of plants are an important basis for the selection of plant species for restoration and reconstruction of degraded ecosystems. In order to study the photosynthetic physiological and ecological characteristics of different mangrove plants, from April to May in dry season and from July to September inrainy season of 2021, the LI-6400 photosynthetic measurement system was used to monitor the photosynthetic physiological parameters and main ecological factors of Kandelia obovata and Bruguiera sexangular in Dongzhaigang National Nature Reserve. The effects of major ecological factors on net photosynthetic rate were analyzed by using path analysis. The results were as follows: (1) The daily average net photosynthetic rate of Kandelia obovata in the dry season (8.43 μmol-2·s-1) was slightly lower than that in the rainy season (8.67 μmol-2·s-1), and the difference was not significant. In contrast, the rate of Bruguiera sexangular in the dry season (7.03 μmol-2·s-1) was significantly lower than that in the rainy season (9.41 μmol-2·s-1). The rate of Kandelia obovata was significantly higher than that of Bruguiera sexangular in dry season, while the rate of Kandelia obovata was significantly lower than that of Bruguiera sexangular in rainy season. (2) The daily average variations of transpiration rate, stomatal conductance and intercellular CO2 concentration of Kandelia obovata were smaller than those of Bruguiera sexangular and the water use efficiency was also lower than that of B. sexangular in dry and rainy seasons. (3) There was a phenomenon of “midday depression of photosynthesis” in both dry and rainy seasons of? Kandelia obovata and Bruguiera sexangular. In dry season, the main reason for the phenomenon of Kandelia obovata might be due to non-stomatal limitation, while Bruguiera sexangular due to stomatal limitation. Inrainy season, the phenomena of two mangrove plants were both mainly caused by stomatal limitation. (4) In the dry season, the main decision factor affecting the net photosynthetic rate of Kandelia obovata was photosynthetically active radiation, and the main limiting factor was relative humidity, while ecological factors played a limiting role on the net photosynthetic rate of Bruguiera sexangular of which saturated water vapor pressure difference was the main limiting factor. In the rainy season, the main decision factor was photosynthetically active radiation, and the main limiting factor was saturated water vapor pressure difference for both Kandelia obovata and Bruguiera sexangular. In summary, Kandelia obovata is more adaptable to habitat changes, and Bruguiera sexangular not only has better drought tolerance performance, but also more suitable for hot and rainy habitats, which will be the scientific reference for the selection of plant for restoration and reconstruction of degraded mangroves.
Key words: net photosynthetic rate, seasonal change, mangrove plants, path analysis,Dongzhaigang of Hainan
紅樹林是生長在熱帶和亞熱帶海岸潮間帶內以紅樹植物為主的灌木或喬木組成的木本植物群落(羅忠奎等,2007),在防風消浪、促淤造陸、固岸護堤和維護生物多樣性等方面發揮重要作用,是人類重要的生態安全屏障(王友紹等,2021)。由于紅樹林位于海陸過渡帶,生態環境脆弱,加上沿海地帶日益密集的開發活動的影響,我國紅樹林遭受嚴重破壞(但新球等,2016)。海南東寨港國家級自然保護區雖然有全國成片面積最大、種類最全的紅樹林,但與20世紀60年代相比,這里的紅樹林濕地面積減少近50%,紅樹林濕地生態系統面臨嚴重威脅(孫艷偉等,2015;楊玉楠等,2020)。2012年以來,地方政府開展系列強有力的環境綜合整治工作,禁止濕地圍墾,嚴控海水養殖和污水排放;同時,開展紅樹林濕地恢復重建的研究(徐蒂等,2014)。
適生物種的選擇是受損紅樹林濕地恢復重建的關鍵(Kamali & Hashim, 2011)。近年來,東寨港自然保護區開展了紅樹林種苗基地建設和栽培撫育研究,栽培了秋茄(Kandelia obovata)、海蓮(Bruguiera sexangula)、紅海欖(Rhizophora stylosa)、桐花樹(Aegiceras corniculatum)、欖李(Lumnitzera racemosa)、海桑(Sonneratia caseolaris)、木欖(Bruguiera gymnorrhiza)等紅樹植物,修復受損紅樹林生態系統(呂佳和李俊清,2008;張穎等,2021)。但是,紅樹林恢復重建研究還比較滯后(吳瑞等,2015;Zhang et al., 2021),紅樹林立地條件研究較少(鐘才榮等,2018;吳庭天等,2020),已修復紅樹林濕地植物群落健康狀況評價缺失(王麗榮等,2011)。
光合生理參數既是植物生長發育的基礎,也是評價植物生長健康狀況的重要指標,還是植物對環境的適應能力和受脅迫程度的反映(黃麗等,2012;秦文華等,2022)。東寨港自然保護區地處熱帶季風氣候區,光照強,氣溫高,旱季降水稀少,雨季降水豐沛,紅樹植物適應這樣氣候的光合生理機制尚不清楚(Feng et al., 2022)。目前,對于紅樹植物光合生理生態特性研究主要集中于紅樹林光合固碳(魏龍等,2020)、溫度(Akaji et al., 2019;鄭春芳等,2020)、鹽度(Lopes et al., 2019)、光強(Feng et al., 2022)、重金屬(Ma & Yang, 2022)等因子脅迫下植物光合作用的變化特征,并且聚焦于幼苗的室內控制實驗,而對自然狀態下不同紅樹種類的光合特性差異和不同季節植物光合特性的變化的綜合分析還相對較少。鑒于此,本文選取海南東寨港國家級自然保護區紅樹林生態修復重要樹種秋茄和海蓮為研究對象,通過對旱季、雨季光合生理參數與生態因子監測,擬解決以下問題:(1)秋茄和海蓮光合生理參數時間變化;(2)秋茄和海蓮凈光合速率及其與生態因子關系;(3)秋茄、海蓮立地及生境適宜性。本研究可為退化紅樹林生態系統的恢復、重建、保育提供科學依據,為國家“南紅北柳”生態修復工程提供數據積累。
1 材料與方法
1.1 試驗區概況
試驗地點位于海南省海口市東北部海南省東寨港國家級自然保護區境內(110°32′—110°37′ E、19°57′—20°01′ N),保護區地處熱帶北緣,屬熱帶季風氣候,年均溫23.8 °C,7月和1月平均氣溫分別是27.9 °C和17.9 °C;全年平均日照時長2 200 h;年平均降雨量1 676.4 mm,分配極不均勻,全年分為旱、雨兩季,雨季大約于5月底或6月初開始,10月底或11月初結束,降水量占全年的70%~90%;太陽年輻射總量約462 kJ·cm-2。該地最高潮水位2.61 m,最低潮水位0.48 m,平均潮差1.0 m。土壤類型為紅樹林沼澤化鹽土,pH值為7.0~7.5,海水鹽度為25‰~28‰(趙鵬,2010)。保護區有紅樹植物29種,半紅樹植物12種,是海蓮、秋茄、紅海欖等紅樹植物重要的植物種源基地(王佳燕,2007),建有種苗基地,用于受損紅樹林濕地恢復與重建。
1.2 試驗材料和方法
試驗樣地(110°35′ E、19°57′ N)位于東寨港自然保護區內的恢復試驗區,地處高潮線附近的紅樹林群落內緣,觀測期間會受到周期性潮汐的浸淹,按照均勻、隨機和代表性原則在樣地內各選取3株長勢良好、無病蟲害的秋茄和海蓮作為監測植株(表1)。在監測植株冠層中上部,各選取3片生長完好、充分展開、光照充足、朝向一致的成熟葉片(梢端完全展開葉第3~5葉位葉片),每片葉重復3次測量,取平均值進行分析。若需要更換測量葉片,則更換年齡、長勢相同枝條上的同位葉片。
利用LI-6400光合儀,于2021年旱季后期4月23—24日、5月12日和雨季7月3日、8月20日、9月30日,選取晴朗無云的時段,從6:00到18:00測定紅樹植物秋茄和海蓮的凈光合速率(net photosynthetic rate,Pn,μmol·m-2·s-1)、蒸騰速率(transpiration rate,Tr,mmol·m-2·s-1)、氣孔導度(stomatal conductance,Gs,mol·m-2·s-1)、胞間CO2濃度(intercellular CO2 concentration,Ci,μmol·mol-1)等生理指標,監測時間間隔1 h。同步記錄光合有效輻射(photosynthetically active radiation,PAR,μmol·m-2·s-1)、大氣溫度(air temperature,Ta,℃)、大氣相對濕度(relative humidity,RH,%)、飽和水汽壓差(vapor pressure deficit,VPD,kPa)、大氣CO2濃度(atmospheric CO2 concentration,Ca,μmol·mol-1)等微氣象環境參數。
1.3 數據處理和分析
旱季、雨季參數為代表性時段測量均值,氣孔限制值(stomatal limitation value,Ls,%)計算公式為Ls=1-Ci/Ca×100%;水分利用效率(water use efficiency,WUE,mmol·mol-1)計算公式為WUE=Pn/Tr。采用Excel對測定數據進行處理,運用SPSS 22.0分析生態因子對Pn的影響,并運用Origin 8.0繪圖。
2 結果與分析
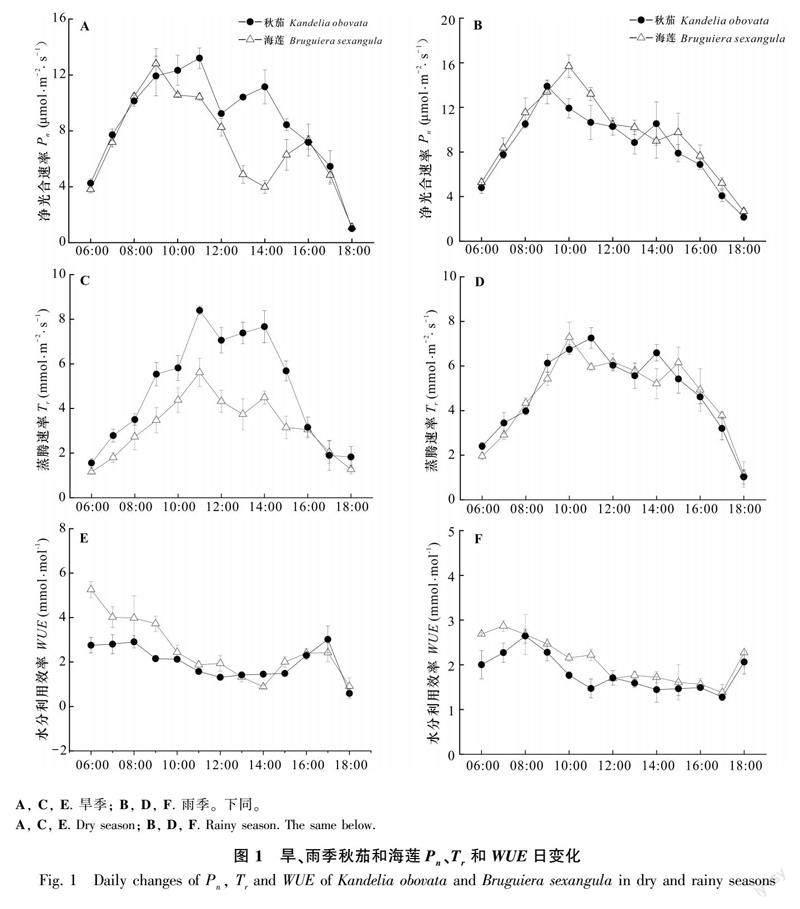
2.1 秋茄、海蓮的光合生理因子日變化
2.1.1 Pn、Tr、WUE的日變化 旱季、雨季秋茄和海蓮的Pn日變化均為“雙峰型”曲線且第一峰值大于第二峰值,“光合午休”現象明顯(圖1:a, b)。旱季,秋茄Pn兩峰值之間時間間隔較海蓮短,秋茄兩峰值較海蓮分別高4.8%和44.7%;雨季,秋茄Pn兩峰值出現時間較海蓮早,第一峰值秋茄較海蓮低12.8%,而第二峰值秋茄較海蓮高2.3%。秋茄雨季Pn第一峰值和第二峰值較旱季分別高12.0%和5.8%,海蓮分別高22.5%和33.5%。旱季秋茄、海蓮的日均Pn分別為8.43、7.03 μmol·m-2·s-1,雨季分別為8.67、9.41 μmol·m-2·s-1,秋茄旱季日均Pn略低于雨季,差異不顯著(P=0.401),而海蓮旱季日均Pn顯著低于雨季(P<0.05);旱季秋茄日均Pn顯著高于海蓮,而雨季秋茄日均Pn顯著低于海蓮(P<0.05);與秋茄相比,海蓮旱季與雨季日均Pn變化幅度較大。
旱季、雨季秋茄和海蓮的Tr日變化呈“雙峰”型曲線(圖1:c, d)。旱季,秋茄和海蓮Tr兩峰值出現時間相同,秋茄兩峰值較海蓮分別高49.6%和88.1%;雨季,秋茄Tr兩峰值之間時間間隔較海蓮要短,秋茄與海蓮的兩峰值比較接近。秋茄旱季Tr的第一峰值和第二峰值分別較雨季高13.9%和9.4%,而海蓮旱季第一峰值和第二峰值較雨季分別低25.6%和37.6%。旱季秋茄、海蓮日均Tr分別為4.79、3.15 mmol·m-2·s-1,雨季分別為4.77、4.69 mmol·m-2·s-1;秋茄旱季日均Tr略高于雨季,差異不顯著(P=0.897),海蓮旱季顯著低于雨季(P<0.05);旱季秋茄日均Tr顯著高于海蓮(P<0.05),而雨季秋茄日均Tr略高于海蓮,差異不顯著(P=0.163);與秋茄相比,海蓮旱季與雨季日均Tr變化幅度較大。
旱季和雨季的秋茄和海蓮WUE峰值均出現在7:00—8:00,之后持續降低(圖1:e,f)。旱季秋茄、海蓮日均WUE分別為1.99、2.50 mmol·mol-1,雨季分別為1.81、2.11 mmol·mol-1;秋茄和海蓮旱季的日均WUE均顯著高于雨季(P<0.05),旱季和雨季秋茄的日均WUE顯著低于海蓮(P<0.05);與秋茄相比,海蓮旱季和雨季的日均WUE變化幅度較大。
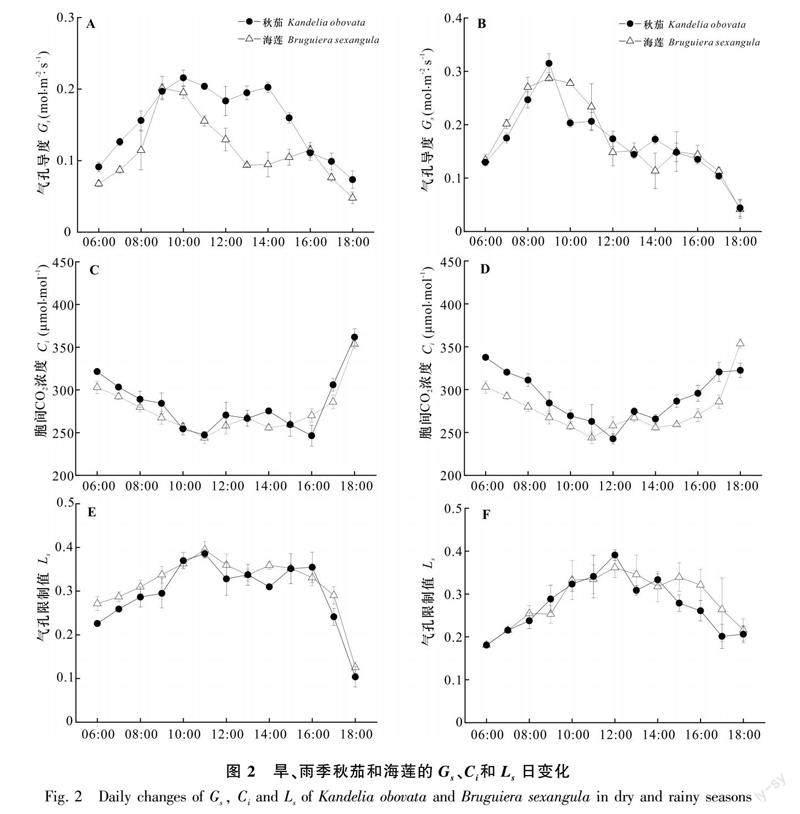
2.1.2 氣孔導度Gs、胞間CO2濃度Ci、氣孔限制值Ls日變化 旱季、雨季秋茄和海蓮的Gs日變化均呈“雙峰型”曲線,且變化幅度較大(圖2:a, b)。旱季,秋茄Gs兩峰值之間時間間隔較海蓮要短,秋茄兩峰值較海蓮分別高12.6%和74.5%;雨季,秋茄Gs兩峰值之間時間間隔較海蓮要短,秋茄兩峰值較海蓮高4.8%和15.2%。秋茄旱季Gs第一峰值較雨季低28.2%,第二峰值較雨季高17.2%,海蓮分別低33.2%和17.4%。旱季秋茄、海蓮日均Gs分別為0.15、0.11 mol·m-2·s-1,雨季分別為0.16、0.17 mol·m-2·s-1。秋茄和海蓮旱季的日均Gs均顯著低于雨季(P<0.05);旱季秋茄日均Gs顯著高于海蓮,而雨季秋茄顯著低于海蓮(P<0.05)。與秋茄相比,海蓮雨季和旱季日均Gs變幅較大。
旱季、雨季秋茄和海蓮的Ci日變化均呈“W”型變化曲線,即峰值出現在6:00和18:00,午間較低,略有波動(圖2:c,d)。旱季秋茄、海蓮日均Ci分別為283.55、275.75 μmol·mol-1,雨季分別為291.85、286.03 μmol·mol-1。秋茄和海蓮旱季日均Ci均顯著低于雨季,旱季和雨季秋茄日均Ci均顯著高于海蓮(P<0.05)。與秋茄相比,海蓮雨季和旱季的日均Ci變幅較大。
旱季、雨季秋茄和海蓮的Ls日變化均呈“雙峰型”曲線(圖2:e, f)。旱季秋茄兩峰值之間時間間隔較海蓮要長,而雨季秋茄兩峰值之間時間間隔較海蓮要短;旱季秋茄Ls日均值較海蓮低6.4%,雨季秋茄較海蓮高3.4%,旱季秋茄和海蓮的兩峰值時間間隔均要比雨季長一些。
2.2 秋茄、海蓮Pn與生態因子的相關性分析
相關分析表明,旱季秋茄Pn與生態因子PAR、Ta和VPD呈極顯著正相關,與RH呈極顯著負相關;旱季海蓮Pn與生態因子PAR呈極顯著正相關,與其他因子相關性不顯著(表2)。雨季秋茄Pn與生態因子PAR呈極顯著正相關,與VPD呈顯著相關,與Ta和RH相關性不顯著;雨季海蓮Pn與生態因子PAR呈極顯著正相關,與其他因子相關性不顯著。
通徑分析結果表明,旱季和雨季,生態因子對秋茄Pn的直接通徑系數排序均為PAR>RH>Ta>VPD,而對海蓮Pn的直接通徑系數排序均為PAR>Ta>RH>VPD(表2)。旱季、雨季PAR對秋茄和海蓮Pn的直接作用大于其他因子的直接作用,同時也大于通過其他因子的間接作用,表明PAR對秋茄和海蓮Pn具有直接決定影響。盡管旱季和雨季RH對秋茄Pn的直接通徑系數較大,起到促進作用,但其他因子的間接通徑系數之和為負,且數值較大,抵消了正向直接作用,導致RH與Pn呈負相關。旱季和雨季VPD對秋茄和海蓮Pn的直接通徑系數為負,起到抑制作用,但其他因子的間接通徑系數之和為正,且數值較大,抵消了負效應,導致VPD與Pn呈正相關。旱季和雨季,生態因子Ta對秋茄Pn的直接通徑系數均小于海蓮,表明與秋茄相比,海蓮Pn對溫度變化更敏感。
決策系數R2表明,旱季,生態因子對秋茄和海蓮Pn的決策系數排序分別為PAR>Ta>VPD>RH、Ta>RH>PAR>VPD。雨季,生態因子對秋茄和海蓮Pn的決策系數排序均為PAR>Ta>RH>VPD(表2)。由此可見,旱季影響秋茄Pn的主要決策因子為PAR,主要限制因子為RH,而生態因子對海蓮Pn均起限制作用,其中VPD為主要限制因子;雨季影響秋茄和海蓮Pn的主要決策因子均為PAR,主要限制因子均為VPD。
3 討論與結論
3.1 秋茄和海蓮的Pn季節變化
不同樹種以及同一樹種的Pn因光照、溫度、土壤等環境因子影響存在較大差異(Pandi et al., 2018;Lele et al., 2021)。研究表明,紅樹植物Pn、Gs以及Tr隨光照、溫度的降低而降低(刁俊明和陳桂珠,2008;Feng et al., 2022),而紅樹植物的凈光合速率、氣孔導度因水土環境中鹽分的增加而降低(Lopes et al., 2019;Ravi et al., 2022)。本研究發現,雨季秋茄和海蓮的Pn和Gs高于旱季,原因可能在于熱帶地區雨季較高的光合有效輻射、比旱季更適宜的溫度以及較低的海水鹽度使得紅樹植物葉片Gs增大,內外氣體交換增加,Pn升高。黃麗(2013)也證實夏季秋茄Pn高于春季。本研究還發現旱季秋茄Pn和Gs高于海蓮,而雨季則是海蓮高于秋茄,這主要與秋茄、海蓮耐鹽性和耐光抑制的差異有關(廖寶文等,2010;Xing et al., 2019);海蓮耐鹽性較秋茄弱,雨季降水豐富,海水鹽度降低,有利于耐鹽性相對較弱的海蓮生長,而旱季較高的海水鹽度抑制海蓮生長;加上海蓮耐光抑制能力強于秋茄,是海蓮在海水鹽度較低以及光照強度更高的雨季光合作用較秋茄強的重要原因。
3.2 秋茄、海蓮光合午休的生理機制
凈光合速率下降可歸因為氣孔限制和非氣孔限制(Farquhar & Sharkey, 1982)。如果Pn下降,Ci也下降且Ls增加,則Pn降低的主要原因是由氣孔因素引起,若Ci上升而Ls下降,則Pn降低則由非氣孔因素引起。本研究發現,雖然旱、雨季秋茄和海蓮Pn均存在“光合午休”現象,但其機制并不相同。旱季秋茄在Pn下降時,Ci上升而Ls下降,表明旱季秋茄Pn下降屬于非氣孔限制。張小燕等(2021a)研究發現,高溫能大幅降低秋茄種群葉綠素含量和PS Ⅱ和PS Ⅰ效率;研究區旱季后期的高溫可能導致秋茄葉片的葉綠體蛋白質結構改變,葉片葉綠素含量降低,PS Ⅱ和PS Ⅰ活性下降,從而引起Pn下降。旱季海蓮Pn下降的時段內,Ci下降且Ls上升,表明旱季海蓮Pn下降屬于氣孔限制。旱季后期因溫度較高、蒸發強,降水較少,海水鹽度增加,而鹽度的升高會使得耐鹽性相對較弱的海蓮吸水困難,疊加上葉片蒸騰失水,導致氣孔關閉,葉片凈光合速率下降(廖寶文,2010;Reef et al., 2015)。雨季,兩種紅樹植物在“光合午休”期間,Pn下降的時段內,Ls上升達到最大且Ci處于最低值,均表現為氣孔限制。可能在于雨季午間光合有效輻射和溫度高,而相對濕度較低,植物為減少蒸騰而關閉氣孔,增加了CO2向葉內擴散阻力,導致CO2供應不足,從而使Pn下降;李林鋒等(2015)研究發現紅樹植物木欖7月份的“光合午休”以及黃麗等(2012)發現秋茄8月份午間Pn下降均由氣孔限制原因引起。
3.3 秋茄、海蓮生境適宜性
與海蓮相比,秋茄旱季、雨季的Pn、Tr、Gs、Ci等光合生理日均值變化幅度均較小,表明秋茄具有較強的生理調控能力,對生境的變化有較強的適應性。秋茄的根系可以通過合成大量滲透物質來抵抗鹽脅迫,秋茄適宜生長的鹽度范圍比海蓮要大(廖寶文,2010;邢建宏等,2017);秋茄葉片柵欄組織、海綿組織異常發達,單寧含量多,具有較強耐輻射能力和保持水分能力(韋江玲和王增軍,2019)。與雨季相比,秋茄、海蓮旱季的WUE較高,表明秋茄、海蓮為適應熱帶季風氣候區旱季高溫少雨、高鹽的環境脅迫,形成較高的水分利用效率適應機制。Clough和Sim(1984)研究發現,紅樹植物的水分利用效率會隨著高溫、干旱、高鹽等外界脅迫環境壓力的增大而增加。本研究發現,旱季、雨季海蓮的WUE均高于秋茄,表明海蓮具有更好的耐旱性能。海蓮木質部具有寬窄兩種類型導管的生態解剖學特征以及葉片氣孔密度與葉脈密度小而密的特征增強其抵御干旱脅迫的能力(蔣夢瑩,2008;張小燕等,2021b),是其耐旱性能較好的重要原因。
秋茄和海蓮Pn與生態因子間的關系分析結果顯示,旱季和雨季,生態因子Ta對秋茄Pn的直接通徑系數均小于海蓮,表明海蓮Pn對溫度變化較秋茄更加敏感;旱季生態因子Ta對秋茄Pn的直接通徑系數和決策系數均大于雨季,海蓮的直接通徑系數和決策系數則均為雨季大于旱季,表明海蓮較秋茄更喜歡高溫多雨的生境。
綜上所述,雨季秋茄和海蓮的“光合午休”均屬于氣孔限制;旱季海蓮的“光合午休”屬于氣孔限制,而秋茄屬于非氣孔限制。雨季秋茄和海蓮Pn日均值均高于旱季;旱季、雨季秋茄Pn、Tr等光合生理季節變幅均低于海蓮,其對于生境變化適應性更強,具有更廣的生境范圍;海蓮旱季、雨季水分利用效率均高于秋茄,其對干旱逆境條件有著更強的適應性。因此,在濱海濕地修復工程中,依據紅樹植物生理特性以及當地環境特征,適地適樹適法開展退化紅樹林生態系統的恢復和重建工作,對國家的“南紅北柳”生態工程建設與可持續發展具有重要意義。
參考文獻:
AKAJI Y, INOUE T, TOMIMATSU H, et al., 2019. Photosynthesis, respiration, and growth patterns of Rhizophora stylosa seedlings in relation to growth temperature[J]. Trees, 33(4): 1041-1049.
CLOUGH BF, SIM RG, 1989. Changes in gas exchange characteristics and water use efficiency of mangroves in response to salinity and vapour pressure deficit[J]. Oecologia, 79(1): 38-44.
DAN XQ, LIAO BW, WU ZB, et al., 2016. Resources, conservation status and main threats of mangrove wetlands in China[J]. Ecol Environ Sci, 25(7): 1237-1243.[但新球, 廖寶文, 吳照柏, 等, 2016. 中國紅樹林濕地資源、保護現狀和主要威脅[J]. 生態環境學報, 25(7): 1237-1243.]
DIAO JM, CHEN GZ, 2008. Effect of light indensity on growth and photosynthetic properties of Sonnerratia apetala seeding[J]. For Res, 21(4): 486-492.[刁俊明, 陳桂珠, 2008. 光強對無瓣海桑幼苗的生長和光合特性的影響[J]. 林業科學研究, 21(4): 486-492.]
FARQUHAR GD, SHARKEY TD, 1982. Stomatal conductance and photosynthesis[J]. Ann Rev Plant Physiol, 33(1): 317-345.
FENG HY, GU XX, TANG T, et al., 2022. Determining the effects of salinity and light on key ecophysiological traits of two nonnative mangrove species in China[J]. J Coast Res, 38(2): 361-368.
HUANG L, 2013. The seasonal dynamics of photosynthesis of Kandelia candel, Avicennia marina, Aegiceras corniculatum and? Bruguiera gymnorrhiza in Zhangjiang estuary[J]. Wetland Sci, 11(1): 82-89.[黃麗, 2013. 福建省漳江口秋茄、桐花樹、白骨壤和木欖光合作用季節動態研究[J]. 濕地科學, 11(1): 82-89.]
HUANG L, TAN FL, WU QC, et al., 2012. A study on the diurnal change of photosynthesis of Kandelia candel, Avicennia marina,and Aegiceras corniculatum at Zhangjiang estuary[J]. J Fujian For Sci Technol, 39(2): 28-34.[黃麗, 譚芳林, 吳秋城, 等, 2012. 福建漳江口3種紅樹植物光合作用日變化特性研究[J]. 福建林業科技, 39(2): 28-34.]
JINAG MY, 2018. Ecological anatomical study on secondary xylem of Sonneratia apetala and Bruguiera Sexangula? [D]. Fuzhou: Fujian Agriculture and Forestry University.[蔣夢瑩, 2018. 無瓣海桑和海蓮次生木質部的生態解剖學研究[D]. 福州: 福建農林大學.]
KAMALI B, HASHIM R, 2011. Mangrove restoration without planting[J]. Ecol Eng, 37(2): 387-391.
LELE N, KRIPA MK, PANDA M, et al., 2021. Seasonal variation in photosynthetic rates and satellite-based GPP estimation over mangrove forest[J]. Environ Monit Assess, 193(2): 1-20.
LI LF, WU XF, LIU SQ, 2015. Characteristics of photosynthesis and photosynthetic carbon fixation capacity of five mangrove tree species in Zhanjiang City[J]. Guihaia, 35(6): 825-832.[李林鋒, 吳小鳳, 劉素青, 2015. 湛江5種紅樹林樹種光合作用特性及光合固碳能力研究[J]. 廣西植物, 35(6): 825-832.]
LIAO BW, 2010. The adaptability of seedling of three mangrove species to tide-flooding and water salinity[D]. Beijing: Chinese Academy of Forestry.[廖寶文, 2010. 三種紅樹植物對潮水淹浸與水體鹽度適應能力的研究[D]. 北京: 中國林業科學研究院.]
LIAO BW, QIU FY, ZHANG LE, et al., 2010. Effect of salinity on the growth and eco-physiological characteristics of Bruguira sexangula var. rhynchopetala seedlings[J]. Acta Ecol Sin, 30(23): 6363-6371.[廖寶文, 邱鳳英, 張留恩, 等, 2010. 鹽度對尖瓣海蓮幼苗生長及其生理生態特性的影響[J]. 生態學報, 30(23): 6363-6371.]
LOPES DMS, TOGENELLA M, FALQUETO A, et al., 2019. Salinity variation effects on photosynthetic responses of the mangrove species Rhizophora mangle L. growing in natural habitats[J]. Photosynthetica, 57(4): 1142-1155.
LUO ZK, HUANG JH, SUN JX, 2007. On ecological functions and conservation of mangrove forest[J]. J Subtrop Resour Environ, 2(2): 37-47.[羅忠奎, 黃建輝, 孫建新, 2007. 紅樹林的生態學功能及其資源保護[J]. 亞熱帶資源與環境學報, 2(2): 37-47.]
L J, LI JQ, 2008. Research on the restoration of mangroves in Dongzhaigang Hainan[J]. Shangdong For Sci Technol, (3): 70-72.[呂佳, 李俊清, 2008. 海南東寨港紅樹林濕地生態恢復模式研究[J]. 山東林業科技, (3): 70-72.]
MA L, YANG SC, 2022. Growth and physiological response of Kandelia obovata and Bruguiera sexangula seedlings to aluminum stress[J]. Environ Sci Poll Res, 29(28): 43251-43266.
QIN WH, ZHANG Y,ZHU YT, et al., 2022. Variation of net photosynthetic rate of grape and its influencing factors in arid of Northwest China[J]. Guihaia, 42(12): 2157-2166.[秦文華, 張揚, 朱永泰, 等, 2022. 西北干旱區葡萄凈光合速率變化及其影響因素[J]. 廣西植物, 42(12): 2157-2166.]
RAVI S, BADER MKF, YOUNG T, et al., 2022. Are the well-fed less thirsty? Effects of drought and salinity on New Zealand mangroves[J]. J Plant Ecol, 15(1): 85-99.
REEF R, WINTER K, M0RALES J, et al., 2015. The effect of atmospheric carbon dioxide concentrations on the performance of the mangrove Avicennia germinans over a range of salinities[J]. Physiol plant, 154(3): 358-368.
SELVAM PP, GEEVARGHESE GA, RAMACHANDRAN P, et al., 2018. Spatial assessment of net canopy photosynthetic rate and species diversity in Pichavaram Mangrove Forest, Tamil Nadu[J]. Ind J Ecol, 45(4): 717-723.
SUN YW, LIAO BW, GUAN W, et al., 2015. Spatial patterns and environmental factors of rapidly degraded mangroves at Dongzhaigang Harbor in Hainan[J]. J S Chin Agric Univ, 36(6): 111-118.[孫艷偉, 廖寶文, 管偉, 等, 2015. 海南東寨港紅樹林急速退化的空間分布特征及影響因素分析[J]. 華南農業大學學報, 36(6): 111-118.]
WANG JY, 2007. Analysis on relationship among major factors of several mangroves in Dongzhai Harbor, Hainan Island[D]. Danzhou: South China University of Tropical Agriculture.[王佳燕, 2007. 海南東寨港幾種紅樹植物主要生長特征因子間的關系研究[D]. 儋州: 華南熱帶農業大學.]
WANG LR, LI Z, PU YJ, et al., 2011. Health evaluation of mangrove community in Dongzhaigang, Sanyahe, and Qingmeigang Mangrove Nature Reserves of Hainan Province[J]. J Trop Ocean, 30(2): 81-86.[王麗榮, 李貞, 蒲楊婕, 等, 2011. 海南東寨港、三亞河和青梅港紅樹林群落健康評價[J]. 熱帶海洋學報, 30(2): 81-86.]
WANG YS, 2021. Impacts, challenges and opportunities of global climate change on mangrove ecosystems [J]. J Trop Ocean, 40(3): 1-14.[王友紹, 2021. 全球氣候變化對紅樹林生態系統的影響、挑戰與機遇[J]. 熱帶海洋學報, 40(3): 1-14.]
WEI JL, WANG ZJ, 2019. Leaf anatomical structure and its ecological adaptability of four mangrove seedling species[J]. J Anhui Agric Sci, 47(18): 115-118.[韋江玲, 王增軍, 2019. 4種紅樹植物幼苗葉片解剖結構及生態適應研究[J]. 安徽農業科學, 47(18): 115-118.]
WEI L, CAI J, LIN QL, et al., 2020. Photosynthetic carbon fixation and the impact factors of Acanthus ilicifolius shrubs in mangroves of the Shenzhen Bay of China[J]. J Ocean Technol, 39(1): 50-56.[魏龍, 蔡堅, 林秋蓮, 等, 2020. 深圳灣紅樹林老鼠簕灌叢的光合固碳及其影響因子[J]. 海洋技術學報, 39(1): 50-56.]
WU R, ZHAN XF, ZHANG GX, et al., 2015. Research progress in mangrove forests in Dongzhai Harbor of Hainan Island[J]. Wetland Sci Manage, 11(4): 60-62.[吳瑞, 詹夏菲, 張光星, 等, 2015. 海南島東寨港紅樹林研究進展[J]. 濕地科學與管理, 11(4): 60-62.]
WU TT, DING S, CHEN ZZ, et al., 2020. Dynamic analysis of mangrove wetlands based on LUCC and landscape pattern change in Dongzhai Port[J]. For Res, 33(5): 154-162.[吳庭天, 丁山, 陳宗鑄, 等, 2020. 基于LUCC和景觀格局變化的海南東寨港紅樹林濕地動態研究[J]. 林業科學研究, 33(5): 154-162.]
XING JH, PAN DZ, TAN FL, et al., 2017. Effects of NaCL stress on the osmotic substance contents in Kandelia candel roots[J]. Ecol Environ Sci, 26(11): 1865-1871.[邢建宏, 潘德灼, 譚芳林, 等, 2017. NaCl脅迫對秋茄根系滲透調節物質含量的影響[J]. 生態環境學報, 26(11): 1865-1871.]
XING JH, PAN DZ, WANG LX, et al., 2019. Proteomic and physiological responses in mangrove Kandelia candel roots under short-term high-salinity stress[J]. Turk J Biol, 43(5): 314-325.
XU D, LIAO BW, ZHU NH, et al., 2014. A primary analysis on mangroves degradation in Dongzhaigang of Hainan Island[J]. Ecol Sic, 33(2): 294-300.[徐蒂, 廖寶文, 朱寧華, 等, 2014. 海南東寨港紅樹林退化原因初探[J]. 生態科學, 33(2): 294-300.]
YANG YN, LIU J, MYAT T, 2020. Monitoring and evaluation of mangrove weland pollution in Dongzhai harbor of Hainan[J]. Mar Environ Sci, 39(3): 399-406.[楊玉楠, 劉晶, Myat Thiri, 2020. 海南東寨港紅樹林濕地污染監測與評價研究[J]. 海洋環境科學, 39(3): 399-406.]
ZHANG XY, WEE KSA, TADASHI K, et al., 2021a. Differences in photosynthetic responses to simulated heat wave in seedlings of two mangrove species from different provenances[J]. Plant Sci J, 39(4): 424-432.[張小燕, Alison Kim Shan Wee, Tadashi Kajita, 等, 2021. 兩種紅樹植物不同種源幼苗對模擬高溫熱浪的光合生理響應差異[J]. 植物科學學報, 39(4): 424-432.]
ZHANG XY,WEE KSA, TADASHI K, et al., 2021b. Effects of provenance on leaf structure and function of two mangrove species: the genetic adaptation to temperature[J]. Chin J Plant Ecol, 45(11): 1241-1250.[張小燕, WEE Kim Shan Alison, KAJITA Tadashi, 等, 2021. 種源地對兩種紅樹葉片結構和功能的影響:對溫度的適應性遺傳[J]. 植物生態學報, 45(11): 1241-1250.]
ZHANG Y, CHEN GC, ZHONG CR, 2021. Research on endangered mangrove species and recovery status in China[J]. J Appl Ocean, 40(1): 142-153.[張穎, 陳光程, 鐘才榮, 2021. 中國瀕危紅樹植物研究與恢復現狀[J]. 應用海洋學學報, 40(1): 142-153.]
ZHANG YM, ZHANG L, KANG YY, et al., 2022. Biotic homogenization increases with human intervention: implications for mangrove wetland restoration[J]. Ecography: e05835.
ZHAO P, 2010. Study on genetic diversity, photosynthetic characteristics and cold resistance of Kandelia candel in the southeast coast of China[D]. Guangzhou: Guangdong Ocean University.[趙鵬, 2010. 中國東南沿海秋茄(Kandelia candel)遺傳多樣性、光合特性及耐寒性研究[D]. 廣州: 廣東海洋大學.]
ZHENG CF, CHEN W, LIU WC, et al., 2020. Recovery mechanisms of photosynthesis and sucrose metabolism in Kandelia obovata seeding from low temperature stress[J]. Chin J Ecol, 39(12): 4048-4056.[鄭春芳, 陳威, 劉偉成, 等, 2020. 低溫脅迫后紅樹植物秋茄幼苗光合特性及蔗糖代謝的恢復機制[J]. 生態學雜志, 39(12): 4048-4056.]
ZHONG CR, LIN ZW, YANG Y, et al., 2018. Preliminary study on site classification of mangrove in Hainan Island[J]. Mol Plant Breed, 16(20): 6875-6880.[鐘才榮, 林作武, 楊勇, 等, 2018. 海南島紅樹林立地類型劃分研究初探[J]. 分子植物育種, 16(20): 6875-6880.]
(責任編輯 李 莉)
