中國4省份葡萄霜霉病菌群體對烯酰嗎啉的抗性及適合度
2023-06-05 20:42:23劉曉慧周連柱孔繁芳王忠躍黃曉慶張昊
植物保護 2023年3期
劉曉慧 周連柱 孔繁芳 王忠躍 黃曉慶 張昊
摘要
為明確我國山西、山東、河北和云南省葡萄霜霉病菌對烯酰嗎啉的抗性以及田間病菌群體的適合度,本研究采用TaqmanMGB技術檢測了2020年-2021年采自上述4省份292株葡萄霜霉病菌對烯酰嗎啉的抗性頻率;采用葉盤法測定了田間抗性菌株與敏感菌株的適合度。結果表明,上述4個地區的平均烯酰嗎啉抗性頻率及抗性等位基因頻率分別為64.0%和74.7%,其中山東省蓬萊區最高(91.2%和96.3%),河北省廊坊市(73.8%和85.0%)和云南省賓川縣(75.0%和82.6%)次之,山西省清徐縣(16.7%和34.7%)最低。適合度分析發現,田間抗性菌株的產孢能力明顯高于敏感菌株,但二者的復合適合度指數無顯著差異。上述結果表明,4省份霜霉病菌群體均對烯酰嗎啉產生了不同程度的抗性,田間抗性菌株與敏感菌株相比無明顯適合度變化。
關鍵詞
葡萄霜霉病菌;?烯酰嗎啉;?抗藥性;?適合度
中圖分類號:
S?436.631.1
文獻標識碼:?A
DOI:?10.16688/j.zwbh.2022161
Resistance?of?Plasmopara?viticola?populations?to?dimethomorph?in?four?provinces,?China?and?their?fitness
LIU?Xiaohui,?ZHOU?Lianzhu,?KONG?Fanfang,?WANG?Zhongyue,?HUANG?Xiaoqing*,?ZHANG?Hao*
(State?Key?Laboratory?for?Biology?of?Plant?Diseases?and?Insect?Pests,?Institute?of?Plant?Protection,
Chinese?Academy?of?Agricultural?Sciences,?Beijing?100193,?China)
Abstract
In?order?to?clarify?the?resistance?of?Plasmopara?viticola?field?populations?to?dimethomorph?in?Shanxi,?Shandong,?Hebei?and?Yunnan?provinces?of?China?and?their?fitness,?the?dimethomorphresistance?of?292?P.viticola?isolates?collected?from?2020?to?2021?was?determined?by?TaqmanMGB?realtime?PCR,?and?the?fitness?of?resistant?and?sensitive?isolates?was?determined?on?detached?leaves.?The?results?showed?that?the?average?resistance?frequency?and?resistance?allele?frequency?of?P.viticola?from?the?four?provinces?were?64.0%?and?74.7%,?respectively.?Among?them,?the?average?resistance?frequency?and?resistance?allele?frequency?in?the?populations?of?Penglai,?Shandong?province?were?the?highest?(91.2%?and?96.3%),?followed?by?those?in?Langfang?of?Hebei?province?(73.8%?and?85.0%)?and?Binchuan?of?Yunnan?province?(75.0%?and?82.6%),?and?the?lowest?was?detected?in?Qingxu?of?Shanxi?province?(16.7%?and?34.7%).?The?sporulation?capacity?of?the?resistant?isolates?was?significantly?higher?than?that?of?the?sensitive?ones,?but?there?was?no?significant?difference?in?the?composite?fitness?index?between?them.?The?results?indicated?that?the?resistance?of?P.viticola?populations?to?dimethomorph?in?the?four?provinces?has?developed?to?various?degrees,?but?no?significant?difference?existed?in?the?fitness?between?sensitive?and?resistant?isolates.
Key?words
Plasmopara?viticola;?dimethomorph;?fungicide?resistance;?fitness
葡萄是一種營養價值很高的水果,在我國大部分地區均有種植[1]。葡萄霜霉病作為葡萄上危害最為嚴重的一種卵菌病害,具有很強的流行性[23]。由于主栽葡萄品種對霜霉病菌缺乏抗性,在環境條件適宜的情況下,葡萄霜霉病大面積發生,嚴重影響葡萄品質與產量[45]。目前葡萄霜霉病的防治仍以化學防治為主[6],生產上使用較多的藥劑有甲霜靈、嘧菌酯、烯酰嗎啉和霜脲氰等。但藥劑的頻繁使用易導致病原菌抗藥性的產生和發展,增加防治難度和成本。
烯酰嗎啉屬于羧酸酰胺類殺菌劑[5],主要通過干擾纖維素的生物合成,影響細胞壁的沉積,阻礙病菌孢子囊壁的形成[7],對單軸霉屬Plasmopara引起的霜霉病和疫霉菌Phytophthora引起的疫病具有很好的防治效果[8]。自1996年在我國注冊登記以來,廣泛用于葡萄霜霉病的防治[9]。2010年,Sun等[10]測定了我國7省11個地區葡萄霜霉病菌Plasmopara?viticola對烯酰嗎啉的敏感性,發現所有菌株均表現敏感。但隨著烯酰嗎啉的大量、頻繁使用,2014年首次在我國廣西資源縣檢測到了烯酰嗎啉抗性葡萄霜霉菌株[11]。2018年,周連柱[12]測定了我國18個葡萄主產區的葡萄霜霉病菌對烯酰嗎啉的抗性,發現霜霉病菌對烯酰嗎啉的整體抗性頻率較低(37.2%),但不同地區間差異較大,河北省廊坊市等部分地區抗性頻率較高,最高為97.8%。
病原菌在田間能否形成穩定的抗性群體,受抗性菌株的生存適合度、殺菌劑選擇壓力以及環境條件等因素的影響[1314]。CorioCostet等[15]發現,對甲氧基丙烯酸酯類殺菌劑(Quinone?outside?inhibitor,?QoI)敏感及抗性的葡萄霜霉病菌,在無藥劑選擇壓條件下,二者的適合度無顯著差異,具有相當的生存競爭力。王文橋等[14]發現,實驗室條件下經紫外誘導獲得的烯酰嗎啉抗性突變體的適合度低于其原始敏感菌株,并且抗性菌株經繼代培養后,其產孢量減少,適合度顯著下降,生存競爭力下降。但目前田間葡萄霜霉病菌對烯酰嗎啉抗性群體的適合度變化情況尚不明確。開展田間抗性菌株適合度研究對明確田間病原菌群體抗藥性發展趨勢及制定合理的抗藥性治理策略具有重要意義。
我國不同地區氣候條件差異較大,葡萄霜霉病發生情況也不同,導致地區間用藥方案存在差異[16]。山西省清徐縣在葡萄成熟季節(7月-9月)降水集中,葡萄霜霉病發生較為嚴重,但當地整體用藥水平較低,其中,烯酰嗎啉用藥歷史較短,用藥頻率低,每個生長季節使用1~2次,病原菌抗藥性頻率較低[9,17];山東煙臺地區葡萄種植歷史長,葡萄霜霉病發生嚴重,每個生長季需要進行10多次的殺菌劑噴霧,其中烯酰嗎啉用藥頻次高達4~6次[6],存在較高的藥劑選擇壓力[9,18];河北廊坊、昌黎地區在葡萄霜霉病的防治過程中存在輕防重治的現象,每個生長季節使用烯酰嗎啉3~5次,并存在縮短施藥間隔期和增大用藥量等現象,病菌對烯酰嗎啉抗性較高[9,19];云南賓川地區葡萄生長期長,防治霜霉病過程中存在連續單一使用烯酰嗎啉的情況,每個生長季節使用次數2~4次,導致病菌對烯酰嗎啉抗性日益嚴重[9,2022]。上述4省份是我國重要的葡萄種植區域,為進一步明確上述地區葡萄霜霉病菌對烯酰嗎啉抗性頻率以及田間抗性菌株適合度的變化,我們重點進行了以下兩方面的研究:1)?測定不同葡萄產區霜霉病菌對烯酰嗎啉的抗性;2)?分別測定抗性菌株和敏感菌株的侵染率、病斑面積和產孢能力3個適合度參數,比較兩種病原菌群體的適合度差異。為殺菌劑的合理使用提供科學指導,為制定科學合理的葡萄霜霉病菌抗藥性治理策略提供依據。
1?材料與方法
1.1?供試材料
供試菌株分別來自2020年和2021年從河北省廊坊市廣陽區、山東省煙臺市蓬萊區、山西省太原市清徐縣和云南省大理市賓川縣4個主要葡萄產區采集的霜霉病樣。采樣時盡量采集發病初期幼嫩葉片,確保霜霉病菌活力。每個產區選擇2~3個自然村,每個自然村采集50個樣品,每個采樣點采集3~5片病葉,各取樣點之間至少間隔50?m以上。將采集的病樣進行保濕培養及病原菌的分離和純化,共收集到292株葡萄霜霉病菌。
供試葡萄品種為‘里扎馬特,采自中國農業科學院植物保護研究所廊坊中試基地葡萄園。
供試培養基為1%水瓊脂培養基(1%WA)。
1.2?試驗方法
1.2.1?葡萄霜霉病菌的純化與擴繁
將采集的病葉用滅菌水淋洗3~4遍,去除表面雜質,用剪刀剪去葉緣枯黃組織,將病葉背面向上放置于放有兩層濕濾紙的培養皿中,在病葉和培養皿內噴水保濕,放于溫度21℃、相對濕度100%、L∥D=16?h∥8?h的人工氣候培養箱中培養3?d左右,待其長出新鮮霉層。
采集‘里扎馬特當年生副梢頂端第4~5片健康嫩葉作為葡萄霜霉病菌的接種材料。將上述葉片用清水沖凈表面塵土,使用1%次氯酸鈉消毒30?s后用無菌水浸泡2~3次,用無菌吸水紙吸干表面液體后,使用滅菌打孔器打制成直徑1.5?cm的葉盤。將葉盤背面向上置于1%?WA培養基上,每皿10個葉盤。挑取培養出的單個霜霉病菌斑,配制成濃度為1?×?106個/mL孢子懸浮液,每葉盤接種20?μL,放于溫度21℃、相對濕度100%、L∥D=16?h∥8?h的人工氣候培養箱中培養。
1.2.2?葡萄霜霉病菌的DNA提取
使用OMAGE公司生產的真菌DNA試劑盒(D339002)提取帶菌葉盤的DNA,首先向裝有帶菌葉盤的凍存管中加入800?μL?Buffer?FG1,放入液體研磨儀中研磨2?min,確保組織充分破碎,然后按照試劑盒步驟提取。
1.2.3?葡萄霜霉病菌對烯酰嗎啉抗藥性檢測
采用王喜娜[11]建立的TaqmanMGB分子檢測技術檢測葡萄霜霉病菌對烯酰嗎啉抗性。利用核酸濃度測定儀(Nano?Vue?Plus)對所提取DNA進行濃度測定,選擇質量較高的樣品。使用引物Cesa3F(5′GCACAAACACGACAATGTAGACAA3′)和Cesa3R?(5′CGGCTGCTACCTTTACGGCAAA3′),探針Cesa3pR(5′FAMAGCAACGAGCTGAAMGB3′)和Cesa3pS?(5′VICCAGCAACGAGCCGAMGB3′)進行熒光定量PCR擴增。試驗選取抗性、敏感菌株的DNA和ddH2O分別作為陽性、陰性和空白對照。由于葡萄霜霉病菌對烯酰嗎啉的抗性是由隱性等位基因控制,只有當該基因位點的兩個等位基因均發生了純合突變時,病原菌才表現為抗性。因而,當菌株僅具有敏感探針Cesa3pS擴增曲線而無抗性探針Cesa3pR擴增曲線時,則判定該菌株為敏感純合子;當菌株同時具有敏感探針Cesa3pS和抗性探針Cesa3pR擴增曲線時,則判定該菌株為敏感雜合子;當菌株僅具有抗性探針Cesa3pR擴增曲線時,則判定該菌株為抗性菌株。根據檢測結果統計病原菌群體的抗性頻率和抗性等位基因頻率,進而分析病原菌的抗性情況及抗性發展趨勢。其中,抗性頻率=抗性菌株數/總菌株數×100%;抗性等位基因頻率=(抗性菌株數×2+敏感雜合菌株數)/總菌株數×100%。
1.2.4?葡萄霜霉病菌抗烯酰嗎啉菌株適合度測定
采用葉盤法[2325]測定葡萄霜霉病菌對烯酰嗎啉抗性菌株及敏感菌株的侵染率(infection?frequency)、病斑面積(lesion?area)和產孢能力(sporulation?capacity)3個參數,以評估其復合適合度指數(composite?fitness?index)[26]。將135個烯酰嗎啉抗性菌株及121個敏感菌株的孢子囊懸浮液(1×105個/mL)分別接種到離體葡萄葉盤背面,每個葉盤上接種20?μL菌液,每皿放置10個葉盤,每菌株設置3個重復,每菌株共接種30個葉盤。置于培養箱中黑暗處理24?h后,用滅菌濾紙條將葉盤表面殘余液滴吸去,將平板放于溫度21℃、L∥D=16?h∥8?h、相對濕度100%的人工氣候培養箱中培養,7?d后調查。
侵染率(%)為發病葉盤數占總葉盤數的比率;病斑面積(cm2/葉)為每菌株在葉盤上的發病面積;產孢能力(個/cm2·mL)為單位體積下每單位面積產生的孢子囊數量;復合適合度指數=侵染率×病斑面積×產孢能力。
1.2.5?數據分析
利用Excel?2019對抗性頻率、適合度數據進行整理,計算各產區葡萄霜霉病菌抗性頻率及抗性等位基因頻率,并結合Rstudio軟件中fifer、tidyverse、RColorBrewer等軟件包進行葡萄霜霉病菌對烯酰嗎啉抗性頻率及抗性等位基因頻率的精確檢驗,利用GraphPad?5.0繪圖軟件對抗性頻率及抗性等位基因頻率數據進行作圖;利用SPSS?25.0統計軟件對適合度數據進行t檢驗,比較是否存在顯著性差異,利用GraphPad?5.0繪圖軟件對抗性菌株、敏感菌株的侵染率、病斑面積和產孢能力數據進行作圖、利用R語言4.2.0中tidyverse等軟件包對復合適合度指數數據進行作圖。
2?結果與分析
2.1?葡萄霜霉病菌對烯酰嗎啉的抗性情況
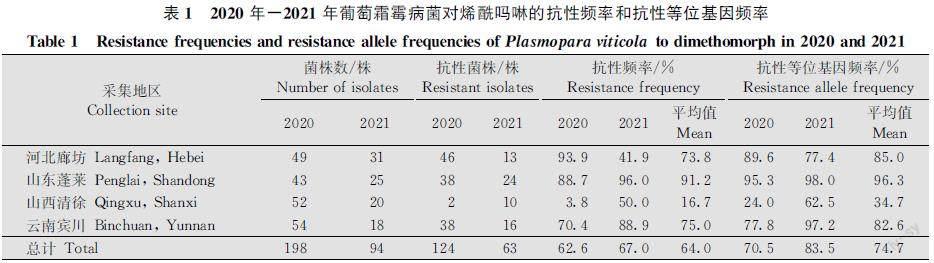
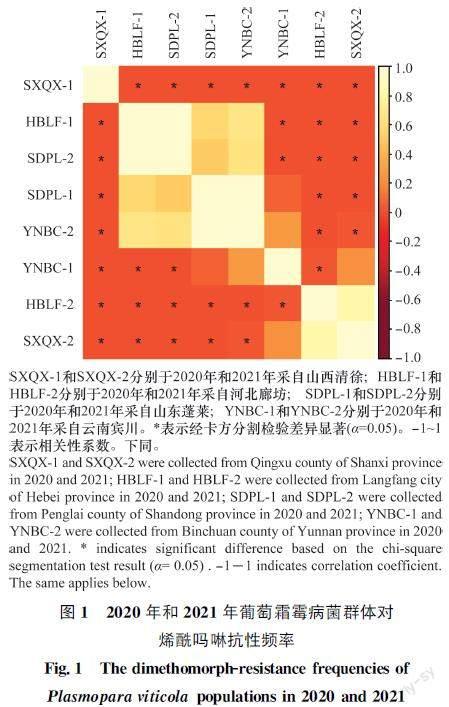
2020年和2021年采自我國河北省廊坊市、山東省蓬萊區、山西省清徐縣和云南省賓川縣4個地區的292株葡萄霜霉病菌對烯酰嗎啉的整體抗性頻率達到64.0%,2020年和2021年采集菌株的抗性頻率分別為62.6%和67.0%(表1)。其中,2020年河北省廊坊市葡萄霜霉病菌抗性頻率最高,達到93.9%,山西省清徐縣葡萄霜霉病菌抗性頻率顯著低于其他3個地區,僅為3.8%;2021年山東省蓬萊區葡萄霜霉病菌抗性頻率高達96.0%,山西省清徐縣(50%)和河北省廊坊市(41.9%)抗性頻率顯著低于其他2個地區(圖1)。
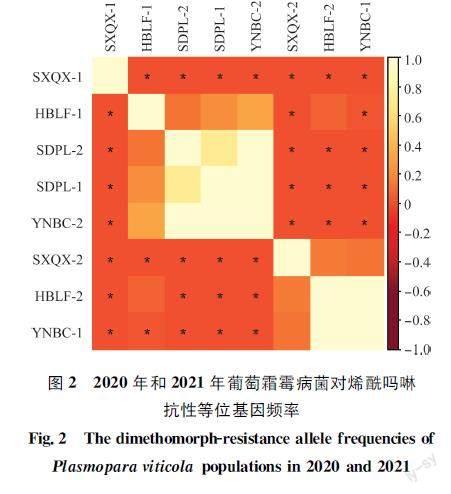
292株葡萄霜霉病菌對烯酰嗎啉整體抗性等位基因頻率為74.7%,2020年和2021年整體抗性等位基因頻率分別為70.5%和83.5%(表1),其中2020年山東省蓬萊區病菌抗性等位基因頻率高達95.3%,山西省清徐縣病菌抗性等位基因頻率為24.0%,顯著低于其他3個地區(圖2);2021年山東省蓬萊區和云南省賓川縣抗性等位基因均超過95%,河北省廊坊市次之(77.4%),山西省清徐縣最低(62.5%)。
與2020年相比,2021年各地區的抗藥性發生情況不同。2021年山西省清徐縣葡萄霜霉病菌對烯酰嗎啉的抗性頻率明顯升高,河北省廊坊市顯著降低(P<0.05);而對于抗性等位基因頻率,2021年山西省清徐縣及云南省賓川縣均明顯升高(P<0.05),其他無顯著變化(P>0.05)。
2.2?葡萄霜霉病菌抗烯酰嗎啉菌株的適合度
采用葉盤法測定292株葡萄霜霉病菌的侵染率、病斑面積和產孢能力3個適合度指標過程中,36株葡萄霜霉病菌被污染,最終得到烯酰嗎啉抗性菌株135株和敏感菌株121株。
侵染率測定結果(圖3a)顯示,2020年和2021年抗性菌株的侵染率均值分別為(88±16)%和(88±14)%,敏感菌株的侵染率均值分別為(86±18)%和(85±19)%,抗性菌株與敏感菌株侵染率均無顯著差異(P>0.05)。
病斑面積測定結果(圖3b)顯示,2020年和2021年的抗性菌株引起的病斑面積分別為(4.16±0.8)cm2/葉和(3.76±1.22)cm2/葉,而敏感菌株引起的病斑面積分別為(3.72±0.72)cm2/葉和(3.55±1.26)cm2/葉,抗性菌株和敏感菌株兩年的病斑面積相當(P>0.05)。產孢能力測定結果(圖3c)顯示,培養7?d后,抗性菌
株和敏感菌株的孢子萌發率均達到100%,其中2020年和2021年抗性菌株的產孢能力分別為(7?819±4?821)個/(cm2·mL)和(6?906±5?107)個/(cm2·mL),敏感菌株的產孢能力(5?133±2?327)個/(cm2·mL)和(4?482±1?679)個/(cm2·mL),抗性菌株兩年的產孢量均顯著高于敏感菌株(P<0.05),表明抗性菌株具有較強的產孢能力。
根據以上3個參數,按照Tooley等[26]建立的公式分別計算出每個菌株的復合適合度指數(圖4)。結果顯示,2020年和2021年抗性菌株的復合適合度指數分別為(27?385±18?357)和(17?354±9?524),2020年和2021年敏感菌株的復合適合度指數分別為(18?000±9?803)和(11?131±7?737),兩年抗性菌株和敏感菌株的復合適合度指數均變化不大(P>0.05),表明烯酰嗎啉抗性菌株與敏感菌株之間沒有明顯的適合度差異。
3?結論與討論
葡萄霜霉病在世界主要葡萄產區嚴重發生[27]。該病流行性極強,一旦暴發,所造成的損失慘重[16],嚴重影響著葡萄質量和產量的提升。化學防治因具見效快、效果好的優點,成為病害防治過程中必不可少的防控手段。大量內吸性殺菌劑,如甲霜靈、嘧菌酯、烯酰嗎啉[2829]等雖具有較好的防治效果,但藥劑的大量頻繁使用加速了病原菌抗藥性的產生和發展,使葡萄生產蒙受了重大損失[912]。烯酰嗎啉因對葡萄霜霉病具有很好的防治效果,且與甲霜靈等苯基酰胺類殺菌劑不存在交互抗性,近些年在生產上廣泛使用[5],葡萄霜霉病菌對其抗性發展迅速[9,30]。
研究發現,葡萄霜霉病菌對羧酸酰胺類殺菌劑的抗性與其纖維素合酶第1?105位的兩個SNP突變(G1105S和G1105V)有關[8,31]。但G1105V突變僅在世界上的少數幾個地點被檢測到,并且抗性頻率很低,歐洲大部分地區及日本等主要以G1105S為主,目前在我國也僅檢測到了G1105S突變[9,30]。基于這一原理,王喜娜等[11]建立了快速檢測葡萄霜霉病菌對烯酰嗎啉抗性的TaqmanMGB分子檢測技術,該技術比葉盤法更為簡便、精確,顯著縮短了檢測時間,提高了檢測精度,為田間葡萄霜霉病菌對烯酰嗎啉抗性的快速檢測及監測提供了技術保障。
本研究發現,我國山西、河北、山東、云南省4個地區均檢測到了葡萄霜霉病菌烯酰嗎啉抗性菌株。其中,山東省蓬萊區、河北省廊坊市和云南省賓川縣的菌株對烯酰嗎啉抗性較為嚴重(兩年的平均抗性頻率均在70%以上),而山西清徐縣的抗性頻率較低(16.7%),這可能與不同地區每個生長季節烯酰嗎啉的用藥次數及用藥量等密切相關[9]。Huang等[9]調查發現2014年-2016年山西省清徐縣每個生長季節用藥次數為1~2次,總用藥量為0.03~0.06?kg/hm2,用藥頻次和用藥量均較少;而山東省蓬萊區、河北省廊坊市和云南省賓川縣每個葡萄生長季烯酰嗎啉用藥次數較高(3~5次),平均用藥量超過1?kg/hm2,高于烯酰嗎啉防治葡萄霜霉病的推薦劑量(0.3~0.5?kg/hm2),長期大量、頻繁的藥劑使用可能導致病原菌產生了抗性,并且伴隨著抗藥性的產生,種植戶為了達到良好的防治效果只得增加用藥量,從而加劇了上述地區霜霉病菌抗藥性的發展。與周連柱2018年對上述4個地區葡萄霜霉病菌烯酰嗎啉的抗性檢測結果相比,山東省蓬萊區、山西省清徐縣和云南省賓川縣三地區葡萄霜霉病菌的烯酰嗎啉抗性頻率和抗性等位基因頻率均呈現上升趨勢,導致該結果的原因可能是檢測地區近兩年葡萄生長季節平均降雨量較常年偏多10.3%~35%[3233],葡萄霜霉病發生較為嚴重,從而導致各產區殺菌劑的使用量隨之增加。雖然山西省清徐縣菌株抗性頻率較低,但抗性等位基因頻率呈逐年上升趨勢,表明病原菌具有一定的抗性發展趨勢,生產上應注意藥劑的合理使用。另外,病原菌抗藥性的產生會引起藥效下降[34],具體藥劑的防治效果需進一步評估,從而為田間精準用藥及制定更科學的抗性治理對策提供依據。
同時,明確無藥劑選擇壓力下葡萄霜霉病菌抗性菌株與敏感菌株適合度變化狀況,對制定科學的抗藥性治理方案也至關重要。本研究選擇測定了采自我國山西省清徐縣、云南省賓川縣、山東省蓬萊區及河北廊坊市4個不同葡萄產區的256個田間菌株中對烯酰嗎啉敏感菌株與抗性菌株在無藥條件下的侵染率、病斑面積和產孢能力,發現抗性菌株的侵染率、病斑面積與敏感菌株不存在顯著差異(P>0.05),而產孢能力顯著高于敏感菌株(P<0.05),生存繁殖能力較強。此外,還發現抗性菌株的復合適合度指數高于敏感菌株,但兩者無顯著差異(P>0.05),該結果與Blum[35]的研究結果一致,原因可能是自然選擇導致適合度顯著下降的抗性菌株被淘汰,從而使兩類菌株表現出適合度相當的狀況。另外,通過連續兩年適合度分析,發現侵染率、病斑面積和產孢能力變化趨穩,可能是由于只有當葡萄霜霉病菌發生隱性純合突變時才表現對烯酰嗎啉的抗性,延緩了病菌對烯酰嗎啉抗性的發展,導致短時間內病原菌群體抗藥性及適合度無太大變化。但上述試驗是在實驗室條件下進行的,給予了病原菌較為適宜的溫度、濕度等生長環境,與田間自然條件下病菌的適合度反應有所差異,一定程度上該方法仍存在缺陷[36]。為此,尚需進一步研究抗性菌株適合度的穩定性以及其與敏感菌株的競爭能力等,以制定更為科學、精準的抗藥性治理策略。
綜上所述,本研究明確了我國河北省廊坊市、山東省蓬萊區、山西省清徐縣和云南省賓川縣4個葡萄產區霜霉病菌對烯酰嗎啉的抗性情況和適合度現狀,為葡萄霜霉病的田間防治以及病原菌的抗藥性治理提供了理論依據。結合國際殺菌劑抗性委員會CAA工作組的相關建議,對于山東省蓬萊區等抗性較嚴重的產區,應避免烯酰嗎啉等羧酸酰胺類藥劑單獨使用,建議與霜脲氰、氰霜唑、氟吡菌胺·霜霉威等其他不同作用機制的藥劑桶混使用[37];在山西省清徐縣等抗性較低的產區,應將羧酸酰胺類殺菌劑與不同作用機理的殺菌劑交替或混合使用,禁止羧酸酰胺類殺菌劑作鏟除性使用,限制每個生長季使用羧酸酰胺類殺菌劑不超過3次,以避免抗藥性發展和擴散[12,37]。
參考文獻
[1]?WONG?F?P,?BURR?H?N,?WILCOX?W?F.?Heterothallism?in?Plasmopara?viticola?[J].?Plant?Pathology,?2001,?50(4):?427432.
[2]?葉正和,?王文相,?張愛芳,?等.?我國卵菌病害化學防治概況[J].?安徽農業科學,?2000,?28(4):?530533.
[3]?司乃國,?劉君麗,?馬學明.?卵菌病害的化學防治現狀與防治策略[J].?農藥,?2000,?39(2):?710.
[4]?SPENCERPHILIPS?P?T?N,?GISI?U,?LEBEDA?A.?Advances?in?downy?mildew?research?[M].?Netherlands:?Kluwer?Academic?Publishers,?2002:?119159.
[5]?朱書生,?盧曉紅,?陳磊,?等.?羧酸酰胺類(CAAs)殺菌劑研究進展[J].?農藥學學報,?2010,?12(1):?112.
[6]?高琪,?李興紅,?劉梅,?等.?我國葡萄園病害發生危害及防治用藥情況調查[J].?中國果樹,?2021(9):?97102.
[7]?FABRITIUS?A?L,?JUDELSON?H?S.?Matingtype?loci?segregate?aberrantly?in?Phytophthora?infestans?but?normally?in?Phytophthora?parasitica:?implications?for?models?of?matingtype?determination?[J].?Current?Genetics,?1997,?32(1):?6065.
[8]?BLUM?M,?WALDNER?M,?GISI?U.?A?single?point?mutation?in?the?novel?PvCesA3?gene?confers?resistance?to?the?carboxylic?acid?amide?fungicide?mandipropamid?in?Plasmopara?viticola?[J].?Fungal?Genetics?and?Biology,?2010,?47(6):?499510.
[9]?HUANG?Xiaoqing,?WANG?Xina,?KONG?Fanfang,?et?al.?Detection?and?characterization?of?carboxylic?acid?amideresistant?Plasmopara?viticola?in?China?using?a?TaqManMGB?realtime?PCR?[J].?Plant?Disease,?2020,?104(9):?23382345.
[10]SUN?Haiyan,?WANG?Hancheng,?STAMMLER?G,?et?al.?Sensitivity?of?Chinese?isolates?of?Plasmopara?viticola?to?metalaxyl?and?dimethomorph?[J].?Journal?of?Phytopathology,?2010,?158(6):?450452.
[11]王喜娜.?我國主要葡萄產區霜霉菌對烯酰嗎啉和嘧菌酯的抗藥性分析[D].?北京:?中國農業科學院,?2017.
[12]周連柱.?我國主要葡萄產區霜霉病菌對五種主要殺菌劑的抗藥性檢測[D].?北京:?中國農業科學院,?2019.
[13]劉召陽,?王帥,?高宇琪,?等.?2株不同地理來源的蘋果樹腐爛病菌對甾醇生物合成抑制劑類殺菌劑的交互抗藥性及生物適合度分析[J].?西北林學院學報,?2020,?35(2):?119124.
[14]王文橋,?劉國容,?張小風,?等.?葡萄霜霉病菌和馬鈴薯晚疫病菌對三種殺菌劑的抗藥性風險研究[J].?植物病理學報,?2000,?30(1):?4852.
[15]CORIOCOSTET?M?F,?DUFOUR?M?C,?CIGNA?J,?et?al.?Diversity?and?fitness?of?Plasmopara?viticola?isolates?resistant?to?QoI?fungicides?[J].?European?Journal?of?Plant?Pathology,?2011,?129(2):?315329.
[16]王忠躍.?中國葡萄病蟲害與綜合防控技術[M].?北京:?中國農業科學技術出版社,?2010:?70.
[17]王喜娜,?王敏,?孔繁芳,?等.?山西省清徐縣葡萄霜霉菌對烯酰嗎啉的抗藥性分析[J].?植物保護,?2018,?44(1):?139142.
[18]李寶燕,?石潔,?寧爽,?等.?山東煙臺葡萄霜霉病菌對烯酰嗎啉的抗藥性分析[J].?中國果樹,?2018(1):?6769.
[19]劉秀香.?河北昌黎釀酒葡萄病蟲害發生與防治[J].?現代農村科技,?2015(8):?2829.
[20]劉彥均,?涂芥兵,?楊愛兵.?云南賓川葡萄產業現狀、問題與發展建議[J].?農業工程技術,?2021,?41(14):?7.
[21]蘇月.?賓川縣葡萄產業發展現狀及問題探析[J].?南方農業,?2019,?13(12):?115117.
[22]杜飛,?繆錢江,?梅馨月,?等.?葡萄霜霉病菌對主要殺菌劑的抗性監測[C]∥中國植物病理學會2015年學術年會論文集.?北京:?中國農業出版社,?2015:?92.
[23]STAHLECSECH?U,?GISI?U,?SOZZI?D.?Determination?of?the?sensitivity?of?Plasmopara?viticola?to?phenylamides?[J].?EPPO?Bulletin,?2010,?22(2):?314316.
[24]馮夏蓮,?何承忠,?張志毅,?等.?植物遺傳多樣性研究方法概述[J].?西南林學院學報,?2006,?26(1):?6974.
[25]趙美琦,?趙幀梅,?馬占鴻,?等.?水稻稻瘟病品種小種組合寄生適合度測定方法的研究[J].?中國農業大學學報,?1997,?2(5):?5157.
[26]TOOLEY?P?W,?SWEIGARD?J?A,?FRY?W?E.?Fitness?and?virulence?of?Phytophthora?infestans?isolates?from?sexual?and?asexual?populations?[J].?Phytopathology,?1986,?76(11):?12091212.
[27]郭俊強,?張曉月,?王榮花,?等.?葡萄品種霜霉病抗性的田間自然鑒定[J].?西北農業學報,?2021,?30(6):?914920.
[28]TOFFOLATTI?S?L,?VENTURINI?G,?CAMPIA?P,?et?al.?Sensitivity?to?cymoxanil?in?Italian?populations?of?Plasmopara?viticola?oospores?[J].?Pest?Management?Science,?2015,?71(8):?11821188.
[29]黃云霄,?李敏,?潘學軍,?等.?幾種殺菌劑對葡萄霜霉病的防治效果[J].?農藥,?2018,?57(11):?836839.
[30]ZHANG?Hao,?KONG?Fanfang,?WANG?Xina,?et?al.?Tetraprimer?ARMS?PCR?for?rapid?detection?and?characterisation?of?Plasmopara?viticola?phenotypes?resistant?to?carboxylic?acid?amide?fungicides?[J].?Pest?Management?Science,?2017,?73(8):?16551660.
[31]AOKI?O,?KAWAGOE?Y,?FUJIMORI?N,?et?al.?Monitoring?of?a?single?point?mutation?in?the?PvCesA3?allele?conferring?resistance?to?carboxylic?acid?amide?fungicides?in?Plasmopara?viticola?populations?in?Yamanashi?prefecture,?Japan?[J].?Plant?Health?Progress,?2015,?16(2):?8487.
[32]佚名.?《2020年中國氣候公報》速覽[N/OL].?中國氣象報,?20210210.?DOI:?10.28122/n.cnki.ncqxb.?2021.000182.
[33]王純枝,?趙秀蘭.?2021年秋季氣象條件對農業生產的影響評價[J].?中國農業氣象,?2022,?43(3):?240243.
