木霉D5菌株的鑒定及其殺線活性測定
2023-06-05 20:42:23陳瑞蓮毛維興劉永紅張樹武徐秉良
植物保護 2023年3期
陳瑞蓮 毛維興 劉永紅 張樹武 徐秉良


摘要
通過形態學和分子生物學方法鑒定了1株殺線木霉D5菌株的種類,并評價了其孢子懸浮液和發酵液對禾谷孢囊線蟲Heterodera?avenae?2齡幼蟲的毒殺活性。結果表明,木霉D5菌株為綠色木霉Trichoderma?viride(ITS序列登錄號:OL661629;TEF序列登錄號:ON241311)。濃度為1.0×107?cfu/mL的D5菌株孢子懸浮液和發酵液原液處理2齡幼蟲72?h后,線蟲死亡率分別為85.20?%和86.57?%。在溫度為20~80℃范圍內以及pH為2~10時,發酵液均具有較高殺線活性。
關鍵詞
綠色木霉;?鑒定;?禾谷孢囊線蟲;?發酵液;?殺線活性
中圖分類號:
S?476
文獻標識碼:?A
DOI:?10.16688/j.zwbh.2022184
Identification?of?Trichoderma?strain?D5?and?determination?of?its?nematicidal?activity
CHEN?Ruilian1,?MAO?Weixing1,?LIU?Yonghong1,?ZHANG?Shuwu1,2*,?XU?Bingliang1*
(1.?College?of?Plant?Protection,?Gansu?Agricultural?University,?Lanzhou?730070,?China;?2.?Biocontrol
Engineering?Laboratory?of?Crop?Diseases?and?Pests?of?Gansu?Province,?Lanzhou?730070,?China)
Abstract
The?species?of?Trichoderma?spp.?D5?strain?was?identified?by?morphology?and?molecular?biology?methods,?and?also?the?nematicidal?activity?of?its?spore?suspension,?fermentation?broth?were?evaluated?in?the?present?test.?The?results?showed?that?the?Trichoderma?spp.?D5?strain?was?identified?as?Trichoderma?viride?(ITS?sequence?accession?number:?OL661629;?TEF?sequence?accession?number:?ON241311).?The?mortality?of?the?secondstage?juveniles?of?nematodes?were?85.20?%?and?86.57?%?72?h?after?treatment?with?the?concentration?of?1.0×107?cfu/mL?spore?suspension?and?fermentation?broth?of?D5?strain,?respectively.?The?highest?nematicidal?activity?of?fermentation?broth?was?found?at?the?temperature?from?20?to?80℃?and?the?pH?value?from?2?to?10.
Key?words
Trichoderma?viride;?identification;?Heterodera?avenae;?fermentation?broth;?nematicidal?activity
禾谷孢囊線蟲Heterodera?avenae?寄主范圍廣泛,可危害多種禾本科作物及牧草[1]。近年來危害面積不斷擴大,已經成為麥類作物的主要病害之一,每年造成經濟損失近千億元[2]。目前,對于禾谷孢囊線蟲的防治主要以化學防治為主,但化學農藥的大量使用不僅造成環境污染、農藥殘留,同時造成線蟲抗藥性逐漸增強。因此,開發高效安全的生物殺線劑是當前研究的主導方向。
木霉是一類廣泛存在于土壤中的習居菌[34]。Harman等報道,木霉至少對18個屬20余種病原真菌和多種病原細菌具有拮抗作用[5]。同時,已有研究表明,多種木霉對線蟲具有毒殺和寄生作用,如深綠木霉Trichoderma?atroviride、哈茨木霉T.harzianum、絨毛木霉T.tomentosum、綠木霉?T.virens、棘孢木霉?T.asperellum?和長枝木霉?T.longibrachiatum等對植物寄生線蟲具有抑制作用,其發酵液對線蟲卵和幼蟲體壁有消解作用,可抑制或延遲卵孵化,并使幼蟲體內空泡化[67]。此外,Siddiqui等[8]利用綠色木霉、哈茨木霉、康氏木霉T.koningii和擬康氏木霉T.pseudokoningii防治爪哇根結線蟲Meloidogyne?javanica,發現其發酵液能明顯降低線蟲卵孵化率并提高對2齡幼蟲的致死率;馬金慧等[9]研究發現哈茨木霉TR12發酵液處理南方根結線蟲48?h致死率可達100?%。此外,毛維興[10]報道了溫度和酸堿度等不同外界因子對長枝木霉T.longibrachiatum?T6菌株發酵液毒殺禾谷孢囊線蟲Heterodera?avenae?2齡幼蟲的殺線活性具有不同程度的影響;彭雙等[11]研究發現溫度、酸堿度和儲藏時間等不同外界因子對枯草芽胞桿菌Bacillus?subtilis和蠟樣芽胞桿菌B.cereus發酵液殺松材線蟲Bursaphelenchus?xylophilus活性也有不同程度的影響。但是有關本課題組分離的木霉D5菌株的種類及外界因素對其殺線活性影響尚未有報道。
因此,本試驗以前期分離的1株木霉D5菌株為研究對象,通過形態學和分子生物學鑒定了其種類,測定了其對禾谷孢囊線蟲2齡幼蟲的毒殺活性及溫度和酸堿度對其殺線穩定性的影響,以便為新型、高效的微生物殺線劑的研制、開發和應用提供理論基礎。
1?材料與方法
1.1?試驗材料
木霉D5菌株由甘肅農業大學植物保護學院植物病毒學和分子生物學實驗室分離并保存。供試線蟲采自河南省小麥禾谷孢囊線蟲發生較嚴重的地塊。
1.2?試驗方法
1.2.1?木霉菌株D5形態學鑒定
將木霉D5菌株接種于PDA平板上,置于25℃、光周期L∥D=16?h∥8?h的條件下培養2?d后,顯微鏡下觀察其菌絲形態,培養4?d后待其大量產孢時,觀察和記錄其菌落形態、顏色。同時,在光學顯微鏡下拍照觀察其分生孢子梗和分生孢子形態特征,并測量其分生孢子大小,每個重復隨機測量100個分生孢子,共重復3次并取其平均值。
1.2.2?木霉菌株D5分子生物學鑒定
采用CTAB法提取木霉D5菌株基因組DNA,并利用通用引物?ITS1/ITS4和TEFF/TEFR(表1)擴增?ITS和TEF片段,引物合成及PCR產物測序均由生工生物工程(上海)股份有限公司完成。然后,將測序獲得的ITS和TEF序列提交NCBI網站進行BLAST比對分析,利用MEGA?7軟件和鄰接法構建系統發育樹(bootstrap=1?000),確定其分類地位,并結合形態學鑒定結果確定其種類。
1.2.3?木霉D5菌株分生孢子懸浮液和發酵液制備
參考張樹武等[12]的方法制備木霉D5菌株分生孢子懸浮液和發酵液。利用血球計數板制備濃度為1.0×104~1.0×107?cfu/mL的分生孢子懸浮液。然后,在PDB培養基?(60?mL,pH=7)?中加入1?mL的1.0×107?cfu/mL?D5菌株分生孢子懸浮液,經發酵培養7?d后,4℃,12?000?r/min離心20?min,利用0.22?μm孔徑的細菌過濾器去除孢子,即獲得無菌發酵液原液,置于4℃冰箱保存備用。
1.2.4?禾谷孢囊線蟲分離及木霉D5菌株殺線活性測定
參考張樹武等[13]的方法,采用“漂浮法”分離禾谷孢囊線蟲孢囊和2齡幼蟲,并利用無菌水配制?(200±10)條/mL的2齡幼蟲懸浮液。
參考張樹武等[14]的方法,利用96孔細胞培養板測定孢子懸浮液和發酵液的殺線活性,每孔加入10?μL?2齡幼蟲懸浮液和90?μL木霉D5菌株孢子懸浮液或發酵液,共5個處理(1.0×104~1.0×107?cfu/mL的分生孢子懸浮液和發酵液原液)和1個對照(無菌水),每個處理和對照均為16個重復。然后,分別置于25℃培養箱并每隔24?h在光學顯微鏡下統計死亡蟲數,連續觀察3?d,計算線蟲死亡率并進行方差分析。
1.2.5?溫度對木霉D5菌株發酵液殺線活性的影響
參考張艷軍等[15]的方法略作修改。利用恒溫水浴鍋將50?mL木霉D5發酵液分別置于20、40、60、80℃和100℃下處理30?min,然后放置冰上待其冷卻至室溫后參照1.2.4方法測定殺線活性。試驗以4℃保存的木霉D5菌株發酵液作為陽性對照,以無菌水作為陰性對照,每個處理和對照均重復3次。
1.2.6?酸堿度對木霉D5菌株發酵液殺線活性的影響
參考陳超等[16]的方法略作修改。利用鹽酸和氫氧化鈉分別將木霉發酵液pH調至2、4、6、8、10和12,4℃放置24?h后,再利用鹽酸和氫氧化鈉調至pH=7后參照1.2.4方法測定殺線活性。試驗以4℃保存的木霉D5菌株發酵液作為陽性對照(自然pH),以無菌水作為陰性對照,每個處理和對照均重復3次。
1.3?數據處理
數據采用Excel?2010軟件進行統計分析和作圖,并采用SPSS?23軟件和Duncan氏新復極差法進行多重比較(P<0.05)。
2?結果與分析
2.1?木霉D5菌株形態學鑒定
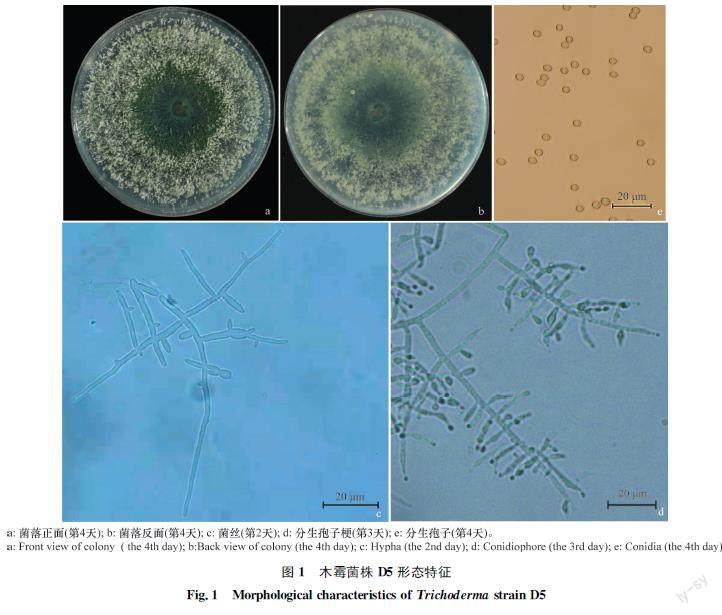
木霉D5菌株在PDA平板上培養初期(1~2?d)形成白色絨狀菌落,有少量白色氣生菌絲,隨著培養時間增加,菌落逐漸增大,并在菌落表面產生大量綠色分生孢子。培養4?d后,菌落長滿整個平皿,菌落背面中央為綠色。菌絲無色,具有隔膜;分生孢子梗無色,有對生或互生分支,分支角度為銳角或近于直角,在分支頂端形成瓶狀小梗,小梗頂端簇生分生孢子;分生孢子多為卵圓形,無色,大小為(2.8×3.5)μm~(2.8×4.2)μm。結合其菌落、菌絲、分生孢子梗和分生孢子形態特征,并參考楊合同主編的《木霉分類與鑒定》[17],將木霉D5菌株初步鑒定為綠色木霉Trichoderma?viride。
2.2?木霉D5菌株分子生物學鑒定
凝膠電泳結果表明,經ITS和TEF引物擴增分別得到大小約為600?bp(圖2a)和300?bp的條帶(圖2b),并經BLAST序列比對發現木霉D5菌株ITS和TEF序列與綠色木霉相似性分別為98.7?%(KY673213.1)和99.83?%(AM498595.1),且ITS(圖3)和TEF(圖4)分別與綠色木霉(KF881775.1)和綠色木霉(AM498579.1)聚為一支。因此,結合形態學特征和分子生物學檢測將木霉D5菌株鑒定為綠色木霉Trichoderma?viride(ITS序列GenBank登錄號:OL661629;TEF序列GenBank登錄號:ON241311)。
2.3?綠色木霉D5菌株孢子懸浮液及發酵液殺線活性測定
不同濃度綠色木霉D5菌株孢子懸浮液和發酵液對禾谷孢囊線蟲2齡幼蟲均具有顯著的毒殺作用。處理24?h后2齡幼蟲開始出現死亡,且隨著孢子懸浮液濃度的升高其殺線活性逐漸增強,1.0×107?cfu/mL孢子懸浮液和發酵原液處理72?h時線蟲死亡率高,分別為85.20?%和86.57?%,且無顯著差異(表2)。
2.4?溫度對綠色木霉D5菌株發酵液殺線活性的影響
試驗表明,綠色木霉D5菌株發酵液具有較好的溫度穩定性,在20~80℃的溫度區間均具有較高的殺線活性且與對照相比無顯著差異,但當溫度繼續升高至100℃時,其殺線活性顯著降低,處理線蟲72?h死亡率為77.74?%,與對照相比降低6.85?%(表3)。
2.5?pH對綠色木霉D5菌株發酵液殺線活性的影響
綠色木霉D5菌株發酵液具有較好的酸堿穩定性,發酵液在pH?2~10條件下處理后,對禾谷孢囊線蟲2齡幼蟲均具有較好的毒殺作用,處理線蟲72?h后死亡率均高于77?%,且與對照相比無顯著差異,但是當發酵液在pH?12條件下處理后,其殺線活性顯著降低,處理線蟲72?h后,線蟲死亡率僅為38.96?%(表4)。
3?結論與討論
木霉作為一種分布廣泛的生防菌,其對植物寄生線蟲毒殺和寄生作用方面已有相關報道,如張樹武等[1819]的研究表明,長枝木霉T6菌株對禾谷孢囊線蟲的孢囊、卵及2齡幼蟲均具有較高的防治效果,其孢子懸浮液(1.5×107?cfu/mL)處理2齡幼蟲72?h后死亡率可達91.3?%,說明木霉在防治禾谷孢囊線蟲方面有巨大的應用前景[20]。本試驗鑒定了一株對禾谷孢囊線蟲2齡幼蟲具有較好毒殺作用的綠色木霉D5菌株,發現其發酵液對2齡幼蟲的毒殺活性高于孢子懸浮液,并且其發酵液殺線活性在溫度和不同酸堿度等因素的脅迫下具有較好的穩定性。
譚可菲等[21]的研究結果表明,不同溫度(30~100℃)對生防菌嗜松青霉Penicillium?pinophilum?SneF8發酵液毒殺南方根結線蟲活性無顯著影響,24?h后其校正死亡率均大于86?%。本試驗中綠色木霉D5菌株發酵液在20~80℃均具有較高的殺線活性,但當溫度升高至100℃時,殺線活性降低,這種差異可能是菌株來源以及不同發酵方式引起的。此外,胡紅濤等[22]研究發現生防菌弗氏鏈霉菌Streptomyces?fradiae?K7發酵液在80℃處理后對黃瓜枯萎病菌Fusarium?oxysporum和立枯絲核菌Rhizoctonia?solani的抑菌效果無顯著差異。趙雅等[23]的研究發現,貝萊斯芽胞桿菌B.?velezensis?HNQ8菌株發酵液在pH?4~10時,具有良好的穩定性,對立枯絲核菌具有較高的抑菌活性,但是強酸性或強堿性環境均會造成其抑菌活性降低,本研究中,綠色木霉D5菌株發酵液具有良好的酸堿穩定性(pH?2~10),這種差異可能是菌株不同造成的。
本研究鑒定了一株具有殺線活性的綠色木霉D5菌株,其發酵液具有較好的殺線活性且在20℃~80℃和pH?2~12時具有殺線穩定性。目前已從木霉菌中提取的殺線活性物質主要有木霉菌素[24]、乙酸[25]和膠酶毒素[26]等,如Yang等[27]研究發現木霉菌YMF1.00416能夠產生對秀麗隱桿線蟲?Caenorhabditis?elegans具有較強致死作用的揮發性有機物,且其對秀麗隱桿線蟲處理48?h后,致死率高達85?%以上。但是,有關綠色木霉D5菌株殺線活性物質鑒定及其殺線作用機制還有待進一步研究。
參考文獻
[1]?HOLGADO?R,?STOEN?M,?MAGNUSSON?C,?et?al.?The?occurrence?and?hosts?of?cereal?cyst?nematodes?(Heterodera?spp.)?in?Norway?[J].?International?Journal?of?Nematology,?2003,?13(1):?119.
[2]?NICOL?J,?RIVOAL?R,?TAYLOR?S,?et?al.?Global?importance?of?cyst?(Heterodera?spp.)?and?lesion?nematodes?(Pratylenchus?spp.)?on?cereals:?distribution,?yield?loss,?use?of?host?resistance?and?integration?of?molecular?tools?[J].?Nematology?Monographs?and?Perspectives,?2003,?2:?119.
[3]?SAMUELS?G?J.?Trichoderma:?A?review?of?biology?and?systematics?of?the?genus?[J].?Mycological?Research,?1996,?100(8):?923935.
[4]?張茹,?李金華,?柴兆祥,?等.?甘肅河西馬鈴薯根際生防木霉菌對接骨木鐮刀菌的拮抗篩選及鑒定[J].?草業學報,?2009,?18(2):?138145.
[5]?HARMAN?G?E,?HOWELL?C?R,?VITERBO?A.?et?al.?Trichoderma?species?opportunistic,?avirulent?plant?symbionts?[J].?Nature?Reviews?Microbiology,?2004,?2(1):?4356.
[6]?焦俊,?韓冰潔,?王媛媛,?等.?毒殺南方根結線蟲的木霉種類鑒定及活性研究[J].?植物保護,?2015,?41(2):?6469.
[7]?ZHANG?Shuwu,?GAN?Yantai,?XU?Bingliang.?Biocontrol?potential?of?a?native?species?of?Trichoderma?longibrachiatum?against?Meloidogyne?incognita?[J].?Applied?Soil?Ecology,?2015,?94:?2129.
[8]?SIDDIQUI?I?A,?AMERZAREEN,?ZAKI?M?J,?et?al.?Use?of?Trichoderma?species?in?the?control?of?Meloidogyne?javanica,?root?knot?nematode?in?okra?and?mungbean?[J].?Journal?of?Biological?sciences,?2001,?4(7):?846848.
[9]?馬金慧,?朱萍萍,?茆振川,?等.?哈茨木霉菌株TRI2的鑒定及其對黃瓜根結線蟲的防治作用[J].?中國農學通報,?2014,?30(22):?263269.
[10]毛維興.?長枝木霉T6菌株培養條件優化及關鍵殺線活性蛋白分析[D].?蘭州:?甘肅農業大學,?2019.
[11]彭雙,?閆淑珍,?陳雙林.?具殺線蟲活性植物內生細菌的篩選和活性產物[J].?微生物學報,?2011,?51(3):?368376.
[12]張樹武,?徐秉良,?古麗君,?等.?長枝木霉對禾谷胞囊線蟲卵的寄生和毒殺作用及其機制[J].?植物保護學報,?2013,?40(4):?320326.
[13]張樹武,?徐秉良,?薛應鈺,?等.?長枝木霉對禾谷胞囊線蟲的寄生和致死作用[J].?微生物學報,?2014,?54(7):?793802.
[14]張樹武,?徐秉良,?薛應鈺,?等.?長枝木霉對南方根結線蟲致死和寄生作用的顯微觀察及測定[J].?植物保護,?2013,?39(4):?4651.
[15]張艷軍,郭榮.?煙草赤星病菌拮抗菌的篩選鑒定及發酵液穩定性分析[J].?浙江師范大學學報(自然科學版),?2019,?42(2):?179183.
[16]陳超,?龐立,?張敏,?等.?獼猴桃葡葡座腔菌拮抗內生菌的篩選及其發酵液穩定性研究[J].?食品科技,?2018,?43(3):?813.
[17]楊合同.?木霉分類與鑒定[M].?北京:?大地出版社,?2009.
[18]張樹武,?劉佳,?徐秉良,?等.?長枝木霉對禾谷胞囊線蟲寄生和致死作用的顯微觀察及測定[J].?應用生態學報,?2013,?24(10):?29552960.
[19]張樹武,?徐秉良,?薛應鈺,?等.?長枝木霉對小麥禾谷孢囊線蟲的致死作用[J].?應用生態學報,?2014,?25(7):?20932098.
[20]SHARON?E,?BAREYAL?M,?CHET?I,?et?al.?Biological?control?of?the?rootknot?nematode?Meloidogyne?javanica?by?Trichoderma?harzianum?[J].?Phytopathology,?2001,?91(7):?687693.
[21]譚可菲,?段玉璽,?陳立杰,?等.?根結線蟲生防菌SneF8發酵代謝產物穩定性研究[J].?江蘇農業科學,?2009(3):?121123.
[22]胡紅濤,?劉振香,?徐文鳳,?等.?生防菌弗氏鏈霉菌K7所產抑菌物質的穩定性研究[J].?黑龍江農業科學,?2019(7):?8588.
[23]趙雅,?張岱,?楊志輝,?等.?貝萊斯芽胞桿菌HNQ8菌株發酵液穩定性測定及抑菌活性成分分析[J].?微生物學通報,?2020,47(2):?490499.
[24]YANG?Zhongshan,?GUO?Hongli,?ZHAO?Peiji,?et?al.?Nematicidal?activity?of?Trichoderma?spp.?and?isolation?of?an?active?compound?[J].?World?Journal?of?Microbiology?and?Biotechnology,?2010,?26(12):?22972302.
[25]DJIAN?C,?PIJAROWSKI?L,?PONCHET?M,?et?al.?Acetic?acid:?a?selective?nematicidal?metabolite?from?culture?filtrates?of?Paecilomyces?lilacinus?(Thom)?Samson?and?Trichoderma?longibrachiatum?Rifai?[J].?Nematologica,?1991,?37(1):?101112.
[26]ANITHA?R,?MURUGESAN?K.?Production?of?gliotoxin?on?natural?substrates?by?Trichoderma?virens?[J].?Journal?of?Basic?Microbiology,?2005,?45(1):?1219.
[27]YANG?Zhongshan,?YU?Zefen,?LEI?Liping,?et?al.?Nematicidal?effect?of?volatiles?produced?by?Trichoderma?sp.?[J].?Journal?of?AsiaPacific?Entomology,?2012,?15(4):?647650.
(責任編輯:田?喆)
