
轉錄因子NbNAC062及其納米藥物對PVY侵染的抑制作用
2023-06-11 21:30:56曲瀟玲宋麗云張道順丁程瀛沈廣材張友臣張曉亮焦裕冰李瑩楊金廣申莉莉
中國煙草科學 2023年2期
曲瀟玲 宋麗云 張道順 丁程瀛 沈廣材 張友臣 張曉亮 焦裕冰 李瑩 楊金廣 申莉莉
摘? 要:前期研究表明NbNAC062能夠抑制PVY早期侵染,本研究通過構建NbNAC062敲除突變體和過表達植株進一步明確NbNAC062的抗病毒功能,并利用異硫氰酸熒光素(fluorescein-5-isothiocyanate,FITC)標記的殼聚糖季銨鹽(chitosan quaternary ammonium salt,HACC)包被NbNAC062質粒,制備HACC-NbNAC062納米藥物。激光共聚焦顯微鏡對納米藥物進行示蹤;透射電鏡和激光粒子分析儀對其表征進行分析。通過GFP熒光差異、qRT-PCR和蛋白免疫印跡檢測病毒含量來探明納米藥物對PVY侵染的影響。結果顯示,敲除組病毒GFP熒光增強,而過表達組病毒GFP熒光減弱,PVY CP含量與上述結果一致。納米藥物粒徑集中分布在18~32 nm之間;Zeta電位為+41.8 mV。浸潤納米藥物HACC-NbNAC062后48 h,在細胞內觀察到FITC-HACC(綠色熒光)與RFP-NbNAC062(紅色熒光);接種PVY-GFP后5、7、9 d,NbNAC062-HACC施藥組的PVY CP mRNA水平較對照組分別下調17.41%、47.81%、13.03%;第7天施藥組PVY CP蛋白水平明顯低于對照組,病毒熒光強度顯著暗于對照組。上述研究結果說明HACC-NbNAC062納米藥物成功遞送了NbNAC062,并發揮了其對PVY初期侵染的抑制作用。
關鍵詞:NbNAC062;馬鈴薯Y病毒;敲除;過表達;HACC-NbNAC062納米藥物
中圖分類號:S435.72? ? ? ? ? ? ? ? ? ? 文獻標識碼:A? ? ? ? ? ? ? ? ? ? 文章編號:1007-5119(2023)02-0035-08
Abstract: Previous studies have shown that NbNAC062 inhibits early PVY infection. In this study, NbNAC062 knockout mutants and overexpression plants were used to further clarify the antiviral function of NbNAC062. The NbNAC062 plasmid was coated with fluorescein isothiocyanate (FITC)-labeled chitosan quaternary ammonium salt (HACC) to prepare HACC-NbNAC062 nanomedicine. Laser confocal microscopy was used to trace the nanomedicine and its characteristics were analyzed by transmission electron microscopy and laser particle analyzer. The viral content was determined GFP fluorescence difference, qRT-PCR and Western blot to explore the effects of nanomedicine on PVY infection. The results showed that the viral GFP fluorescence in the knockout group was enhanced, while the viral fluorescence in the overexpression group was weakened. The PVY CP content was consistent with the above results. The diameter of HACC-NbNAC062 nanomedicine was 18~32 nm; the Zeta potential was +41.8 mV. FITC-HACC (green fluorescence) and RFP-NbNAC062 (red fluorescence) were observed in the plant cells at 48 h after infiltrated with the HACC-NbNAC062 nanomedicine. Comparing with the control group, the PVY CP mRNA levels in the NbNAC062-HACC group were down-regulated by 17.41%, 47.81%, and 13.03% respectively 5, 7, and 9 days after PVY-GFP inoculation. The protein level was significantly lower than that of the control group at 7 days after viral inoculation, and the GFP fluorescence intensity was significantly darker than that of the control group. The above research results indicated that the HACC-NbNAC062 nanomedicine successfully delivered NbNAC062 and exerted its inhibitory effect on the initial infection of PVY.
Keywords: NbNAC062; potato virus Y; gene knockdown; over-expression; HACC-NbNAC062 nanomedicine
馬鈴薯Y病毒(potato virus Y,PVY)能侵染包括茄科、藜科、豆科在內的34屬170余種植物,是煙草上最具破壞性的病毒[1]。自苗期開始噴施抗病毒劑預防是病害的主要控制策略之一,但目前生產上缺少靶向高效藥劑。培育抗病品種是目前最有效的防治措施,而植物抗病基因的挖掘和功能闡明可為植物病害的防治提供新的種質資源。
NAC作為植物特有的轉錄因子,在植物整個生命周期中發揮重要作用[2],不僅調控分生組織形成、側根形成、花器官發育、果實成熟及葉片衰老[3-6]等生長發育過程;還參與低溫、冷害、干旱、鹽害等非生物脅迫下的抗逆反應及發育進程[7-8]。此外,一些NAC轉錄因子還受病菌和病毒侵染的誘導,調控植物的防衛反應。例如,擬南芥AtNAC062能與病程相關蛋白基因PR1、PR2、PR5結合,激發抗性反應,抑制丁香假單胞菌(Pseudomonas syringae)侵染[9];玉米ZmNAC41和ZmNAC100可防御炭疽病菌(Colletotrichum graminicola)侵染[10]。煙草NbNAC089和百合LrNAC35有助于誘導植株對黃瓜花葉病毒(cucumber mosaic virus,CMV)和煙草花葉病毒(tobacco mosaic virus,TMV)的防御反應[11-12]。
將抗病蛋白核酸遞送進入細胞是發揮其抗性作用的前提。納米材料的高通量應用能夠將生物分子快速簡單地引入植物細胞,而無需昂貴且費力的生物分子轉移技術[13-15]。研究發現,殼聚糖季銨鹽(quaternary ammonium salt of chitosan,HACC)可有效包裹核酸使其免受酶的降解,并有效進入細胞,是目前核酸傳遞中廣泛應用的納米材料[16-18]。本課題組前期研究表明,HACC能有效遞送抗病蛋白NbMLP28質粒,增強植物對病毒侵染的抗性[19];本氏煙轉錄因子NbNAC062能通過促進細胞生存信號而抑制PVY病毒的早期侵染增殖[20],但尚未在敲除突變體和過表達植株中驗證。基于敲除突變體和組成型過表達植株的靶標特異性和穩定性,本文以本氏煙為材料,構建NbNAC062敲除突變體和過表達植株,明確其在PVY侵染過程中的作用;創制殼聚糖-DNA質粒納米藥物并檢測其對PVY的防效,以期為PVY的靶向防控提供參考。
1? 材料與方法
1.1? 供試植株、病毒、質粒和試劑
供試煙苗均為5~6葉期溫室土培本氏煙(N. benthamiana),培養溫度(25±1)℃,光照16 h/d,光照強度2000 lx,相對濕度65%;供試病毒為保存于三生NN煙(N. tabacum var. Samsun NN)活體上的PVY,及中國農業科學院煙草研究所病毒組自制侵染性克隆PVY-GFP[21],于–80 ℃保存。
pEarleyGate100-RFP-NbNAC062質粒由煙草所病毒組制備[20],pORE-Cas9質粒由西南大學夏慶友老師惠贈[22];殼聚糖季銨鹽(chitosan quaternary ammonium salt,HACC)和熒光素-5-異氰酸酯(fluorescein-5-isothiocyanate,FITC)購自上海源葉生物科技有限公司,二甲基亞砜(dimethyl sulfoxide,DMSO)購自Sigma-Aldrich公司。測序由派森諾生物科技有限公司完成。
1.2? NbNAC062敲除突變體和過表達植株構建
根據NbNAC062基因序列設計2個sgRNA位點(表1)。將單鏈Oligo DNA退火形成的雙鏈DNA,Bsa I酶切pORE-Cas9產生的線性化表達載體,兩者經T4 DNA連接酶連接后,轉化至大腸桿菌,篩選卡那霉素抗性陽性轉化子,提取質粒,將其轉化至農桿菌,篩選陽性轉化子[2]。根據NbNAC062序列設計含XbaⅠ、EcoRⅠ酶切位點的引物Fu- NbNAC F/R(表1)。擴增NbNAC062并將其連接入Fu46-RFP,構建入門載體Fu46::RFP::NbNAC062;利用LR反應,將其與pEarleyGate100重組構建NbNAC062表達載體,提取質粒并轉化農桿菌。制備農桿菌懸液后,利用葉盤法轉染本氏煙,通過Bar抗性篩選陽性小芽,生根培養[20]。
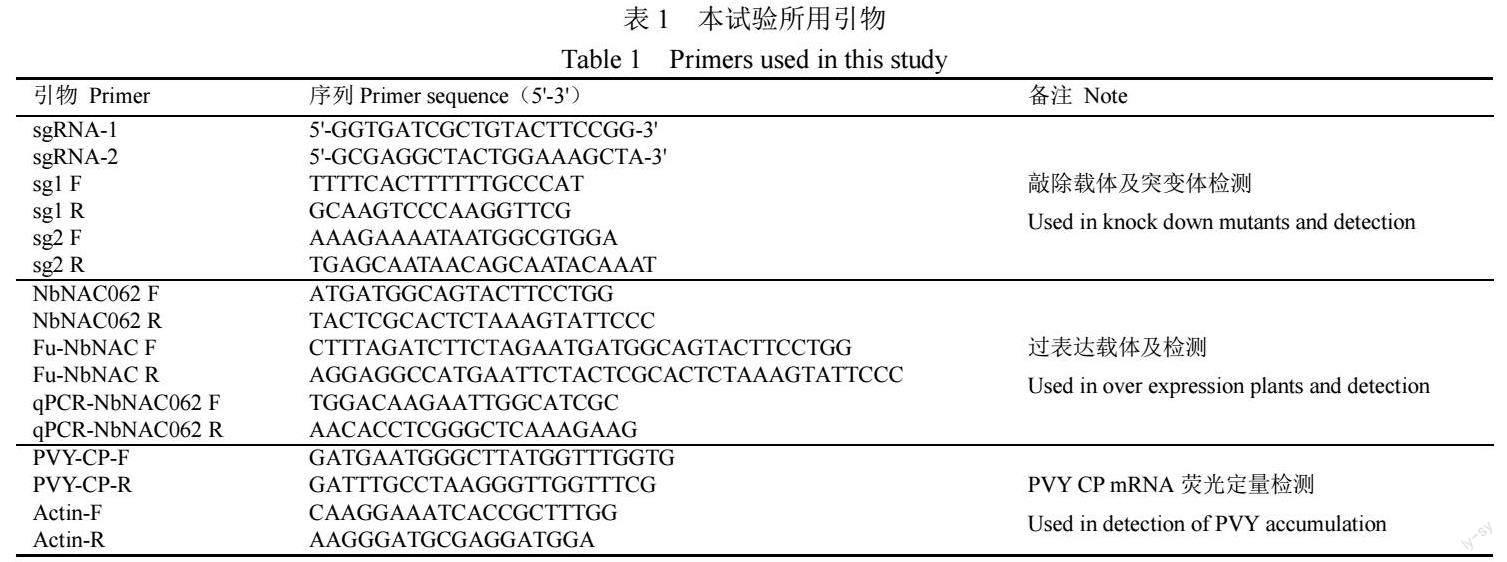
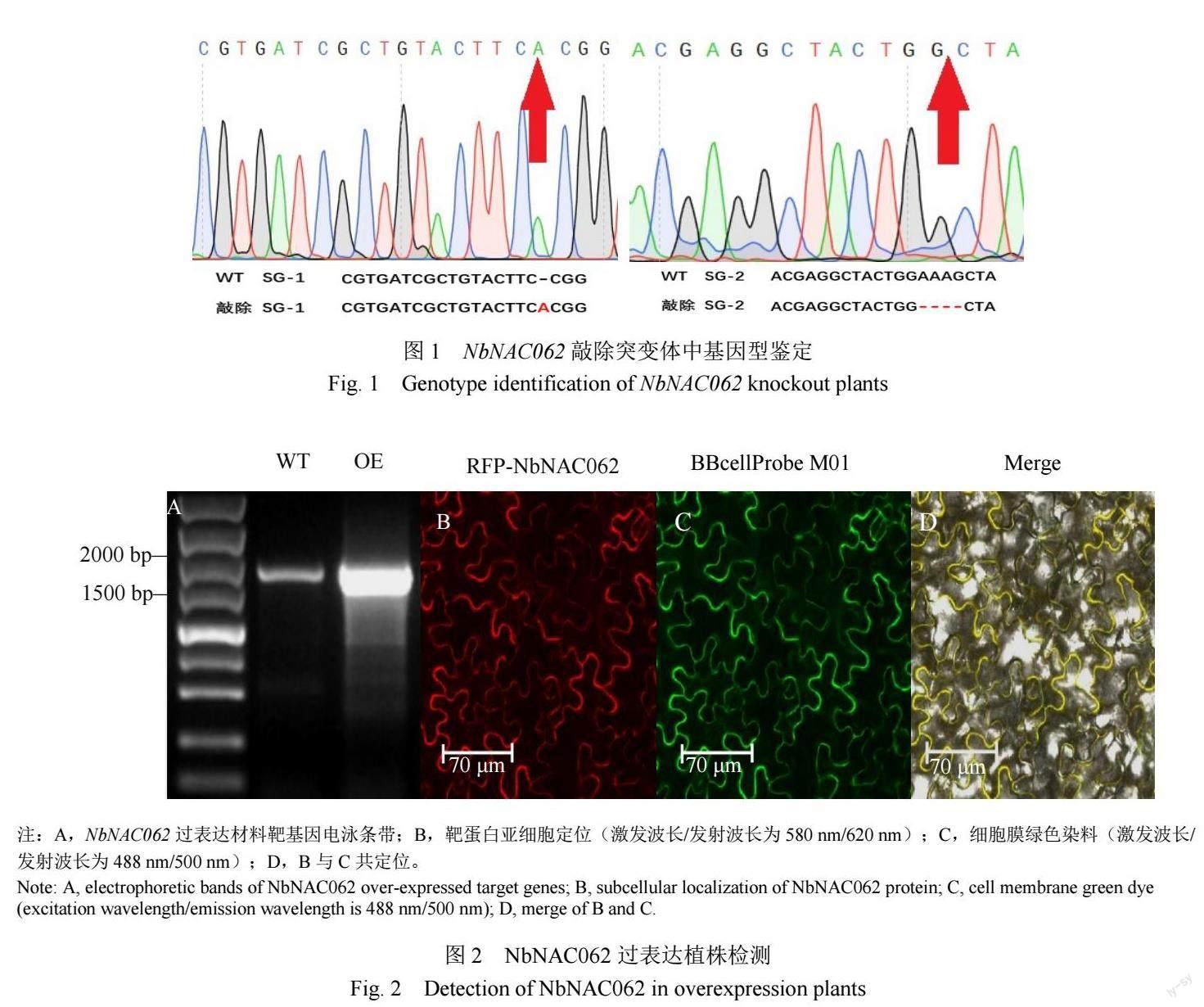
提取敲除突變體葉片總DNA,分別利用sg1 F/R和sg2 F/R檢測引物(表1),進行PCR擴增,測序比對靶基因有效突變株。提取過表達植株葉片總DNA,利用NbNAC062 F/R引物(表1)進行PCR擴增,瓊脂糖凝膠電泳檢測NbNAC062表達量。以細胞膜綠色染液BBcellProbe M01(488 nm/500 nm, Bestbio)為對照,在激光共聚焦顯微鏡下觀察NbNAC062的亞細胞定位[20]。
1.3? NbNAC062敲除突變體和過表達植株的表型及對PVY侵染的影響
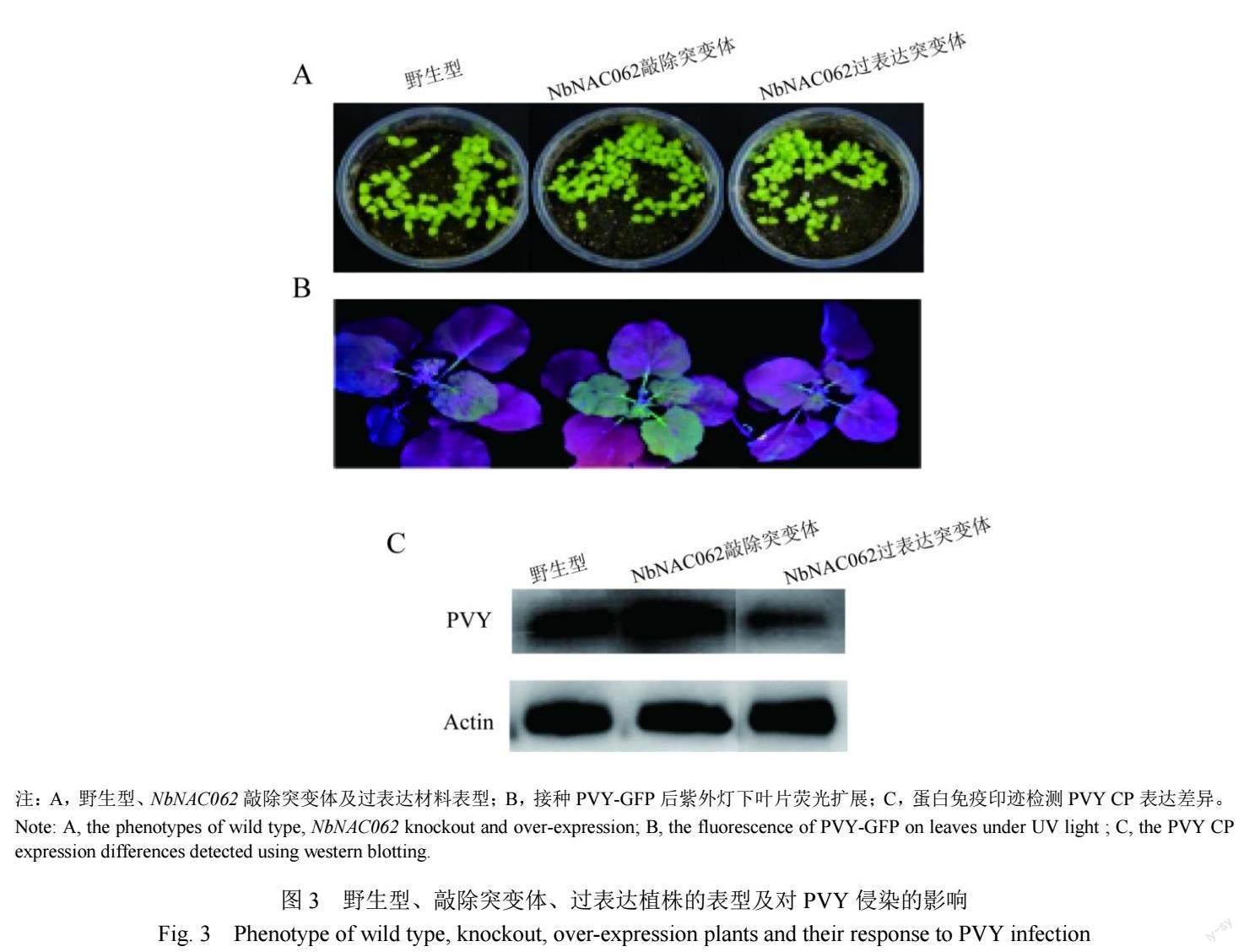
將野生型本氏煙、NbNAC062敲除突變體和過表達植株,分別套袋收種和檢測后,于營養基質土中播種,溫室培養。觀察出苗率、苗期及成株葉片和花器官表型。5~6葉期時,浸潤接種PVY-GFP侵染性克隆。每處理15株,3次重復。接種后7 d在紫外燈下觀察葉片上病毒熒光強度及擴展情況;提取各處理葉片總蛋白,通過Western blot檢測PVY CP蛋白積累差異。
1.4? FITC標記HACC
將10 mg HACC和50 mL ddH2O置于燒杯中,于磁力攪拌器上,800 r/min、25 ℃攪拌至顆粒融化,配置0.2 mg/mL的HACC溶液,4 ℃保存。將20 mg FITC和20 mL DMSO液體置于燒杯中,300 r/min、25 ℃、黑暗條件下攪拌至粉末融化,配置1 mg/mL FITC溶液,4 ℃避光保存。在鋁箔紙包裹的滅菌錐形瓶中,加入等體積的HACC與FITC,25 ℃避光輕輕攪拌3 h后,將混合溶液倒入預先煮沸(10 min)且降至室溫的8000~14000 Da透析袋中,封口后于ddH2O中,4 ℃黑暗透析3 d,制備FITC-HACC溶液[19]。
1.5? NbNAC062納米藥物制備、表征檢測及表達示蹤
將pEarleyGate100-RFP-NbNAC062質粒(200 ng/?L)與HACC溶液(0.2 mg/mL),分別按3∶1、2∶1、1∶1、1∶2、1∶3、1∶4、1∶5(V∶V)混勻后,經55 ℃水浴1 min,渦旋振蕩30 s、靜置10 min,制備殼聚糖-DNA納米藥物HACC-NbNAC062。通過瓊脂糖凝膠電泳,檢測包裹效率。以FITC-HACC溶液替代HACC溶液,制備FITC標記的納米藥物FITC-HACC-NbNAC062。透射電鏡下檢測藥物的微觀形態和大小;激光粒度分析儀檢測其粒徑分布和Zeta電位。于本氏煙下表皮上浸潤納米藥物FITC-HACC-NbNAC062,每片葉200 ?L,48 h后在激光共聚焦顯微鏡下檢測藥物的表達和示蹤[19]。
1.6? HACC-NbNAC02納米藥物對PVY的防效檢測
取長勢均勻的本氏煙兩組,于下表皮浸潤納米藥物FITC-HACC-NbNAC062(200 ?L/葉),以浸潤FITC-HACC的為對照組,每處理移栽15株,3次重復。溫室培養12 h后,分別浸潤接種PVY-GFP侵染性克隆。接種后1、3、5、7、9、11、13、15 d,取接種葉進行qRT-PCR檢測PVY CP mRNA含量或通過Western blot檢測PVY CP蛋白積累。另取部分植株于接種后7、8、10、11、13、15 d,在紫外燈下持續觀察葉片熒光情況[19]。
1.7? 實時熒光定量PCR(Quantitative Real-time PCR)
按照制造商的說明利用TRIzol(Vazyme, 南京)法提取植物總RNA和反轉錄試劑盒合成cDNA(Vazyme, 南京),利用Applied Biosystems 7500快速實時PCR系統(Applied Biosystems,Waltham,MA,USA),使用SYBR Premix Ex TaqTM試劑盒(Vazyme,南京)進行qRT-PCR。β-肌動蛋白基因用作內源對照,使用引物PVY-CP-F和PVY-CP-R檢測病毒外殼蛋白表達的變化。采用–2-△△CT法計算?目的基因的相對表達量,每處理3個生物學重復。
1.8? 蛋白質印跡(Western Blotting)
從本氏煙中提取植物總蛋白質,并與2×蛋白質上樣緩沖(含DTT)液等體積混合后,將蛋白質樣品在95 ℃下孵育3 min,置于12% SDS-聚丙烯酰胺凝膠上分離。然后通過電轉印儀將分離的蛋白質轉移到硝酸纖維素膜上。PVY CP抗體(SRA20001,Agdia,USA)和β-肌動蛋白(CW0264M,CWBIO,北京)抗體用于蛋白印跡。
2? 結? 果
2.1? NbNAC062敲除與過表達材料檢測
在NbNAC062敲除突變體中,PCR分別擴增sg-1、sg-2靶序列。測序結果比對顯示(圖1),sg-1中在第18 bp位置處,插入了1 bp的腺嘌呤脫氧核糖核苷酸(A);sg-2中從第14 bp位置起,缺失了3 bp的腺嘌呤脫氧核糖核苷酸(A)和1 bp的鳥嘌呤脫氧核糖核苷酸(G),依次為AAAG,該突變體為有效敲除,可以篩選純合子作為后續試驗的材料。
在NbNAC062過表達植株中,擴增NbNAC062靶基因,瓊脂糖凝膠電泳檢測顯示,在1944 bp位置出現預期的電泳條帶,且過表達植株的條帶亮度顯著強于野生型(圖2A),說明該材料已過表達NbNAC062。激光共聚焦顯微鏡觀察發現,過表達植株中,顯示紅色熒光的RFP-NbNAC062蛋白(圖2B)與顯示綠色熒光的細胞膜染液BBcellProbe M01(圖2C)發生共定位,產生黃色熒光(圖2D),說明過表達植株NbNAC062蛋白定位于細胞膜,這一結果與之前檢測到的NbNAC062在正常情況下定位于細胞膜相一致[24]。
2.2? NbNAC062敲除與過表達材料表型及對PVY侵染的影響
野生型、敲除、過表達本氏煙材料種子,播種后第10天出苗率、幼苗及開花結果成株表型,三者均無明顯差異(圖3A),說明正常情況下NbNAC062基因的敲除或過表達對植株的生長發育無顯著影響。對5~6葉期野生型、敲除突變體和過表達本氏煙葉片浸潤接種PVY-GFP后7 d,綠色熒光顯示,相較于野生型對照組,敲除組病毒熒光增強,而過表達組病毒熒光減弱(圖3B);PVY CP蛋白表達量亦顯示NbNAC062敲除促進了病毒積累,而過表達抑制了病毒積累(圖3C)。這說明本氏煙轉錄因子NbNAC062對PVY侵染具有一定的抑制作用,可用于納米藥物制備。
2.3? NbNAC062-FITC-HACC藥物示蹤
抗病毒藥物充分發揮作用的前提是藥物進入植物細胞內。如圖4,殼聚糖-DNA納米藥物NbNAC062-FITC-HACC浸潤本氏煙后48 h,激光共聚焦顯微鏡下顯示,紅色熒光標記的融合蛋白RFP-NbNAC062與綠色熒光標記的殼聚糖季銨鹽FITC-HACC發生共定位,產生黃色熒光。說明pEarleyGate100-RFP-NbNAC062質粒可被殼聚糖季銨鹽HACC遞送進入細胞內,并發生表達。
2.4? NbNAC062-HACC納米藥物表征測定
瓊脂糖凝膠電泳檢測HACC對NbNAC062的最大載藥量,顯示pEarleyGate100-RFP-NbNAC062質粒(200 ng/?L)與HACC溶液按照體積比1∶2混均,包裹效率較好(圖5A)。透射電鏡下(TEM),納米藥物NbNAC062-HACC呈近球形的顆粒狀,微觀形態良好(圖5B);激光粒度分析儀測定顯示,藥物顆粒集中分布在18~32 nm之間,粒徑較均勻(圖5C);納米藥物NbNAC062-HACC的Zeta電位為+41.8 mV(圖5D)。說明形態和大小均符合納米藥物的表征要求,且藥物溶液體系穩定性良好,可以開展藥物抑制PVY侵染增殖的室內防效試驗。
2.5? NbNAC062納米藥物對馬鈴薯Y病毒初期侵染的防效檢測
qRT-PCR檢測結果顯示(圖6A),相較于HACC對照組,PVY接種后1、3、5、7、9、11、15 d,NbNAC062-HACC納米藥物處理組的CP表達分別下調15.44%、27.00%、17.41%、47.81%、13.03%、10.98%、7.13%,其中5、7、9 d,藥物處理組的病毒CP表達下調達到了顯著或極顯著水平,接種后7 d的Western blot檢測亦顯示(圖6B),藥物處理組的PVY CP蛋白表達顯著低于對照組。接種后7~11 d的植株紫外熒光亦顯示(圖6C),在7~8 d,NbNAC062-HACC藥物處理組的PVY熒光強度顯著低于HACC對照組,10 d之后熒光強度差異不明顯,與PVY CP表達量檢測相符,顯示植株在接種PVY前預先施用納米藥物,能顯著抑制或延緩PVY的早期積累和擴展。
3? 討? 論
煙草NAC轉錄因子可以調控葉片衰老、參與鹽脅迫和響應病毒侵染[6-7,12,20]。本課題組前期研究表明本氏煙NbNAC062能促進細胞生存而抑制病毒的早期侵染增殖[20]。這一結論在本研究中,通過創制敲除突變體和過表達植株進一步得到驗證,即敲除組對PVY敏感性上升,而過表達組則抑制病毒侵染增殖(圖3)。本氏煙NbNAC062對病毒侵染增殖的抑制作用,與煙草NbNAC089和百合LrNAC35抑制CMV和TMV侵染[11-12],及擬南芥AtNAC089抵御車前草花葉病毒(plantago asiatica mosaic virus,PlAMV)的侵染[9,23]相一致;但與番茄卷葉病毒(tomato leaf curl virus,TLCV)可利用植株的SINAC1基因與自身的復制增強蛋白REn結合,促進自身DNA復制,加速病毒增殖[24]相反。雖然,目前的研究尚不能明確是不同寄主的不同NAC轉錄因子,還是不同病毒類型(TMV、CMV、PVY均為RNA病毒,TLCV為DNA病毒)導致了這種差異。但推測,寄主在通過一些NAC類轉錄因子抑制病毒侵染的同時,一些病毒也會進化出適應途徑,通過主動的誘導激活和劫持利用,而實現自身增殖。此外,本研究中敲除和過表達NbNAC062,正常條件下均不影響植株的生長發育(圖3);但作為重要的試驗材料,后續還應進行干旱、鹽害、冷害等逆境脅迫試驗,進一步闡釋其在煙草抗逆中的功能。
細胞壁不僅是植物的保護屏障,還是外源生物分子進入植物體的主要運輸障礙,限制了功能性外源DNA的傳遞。相較于傳統的病毒類載體,單壁碳納米管、介孔二氧化硅、殼聚糖季銨鹽等無細胞毒性的納米材料是核酸藥物傳遞的安全、有效載體。殼聚糖衍生物殼聚糖季銨鹽(HACC)是核酸傳遞的納米載體,具有生物相容性、可降解性及廣譜抗菌性[16-17],可有效包裹核酸,使DNA免受DNA酶的降解并有效進入細胞[18],目前已應用于醫藥、食品、環保、農業等研究領域。本研究創制的殼聚糖?-DNA納米藥物NbNAC062-HACC,其球形顆粒的形態和大小均符合納米藥物要求[25-26],且藥物穩定性良好,能在細胞內穩定表達(圖4、5);接種病毒前預先浸潤施用NbNAC062-HACC納米藥物,能顯著抑制和延緩PVY的初期侵染和增殖(圖6)。但作為藥物開發,仍需改進施藥方法。NbNAC062作為抑制PVY侵染的功能核酸,要充分發揮其作用,可嘗試制備DNA水凝膠的分子納米藥物[27-28],以更好地穿越核膜,發揮其轉錄因子的調控作用。
4? 結? 論
本研究表明,在正常條件下,敲除和過表達轉錄因子NbNAC062對本氏煙生長發育無顯著影響;在PVY侵染脅迫下,過表達NbNAC062轉基因植株對病毒侵染初期具有抑制作用。殼聚糖-DNA納米藥物NbNAC062-HACC表征和穩定性良好,且能在細胞內表達;預先施用能抑制和延緩PVY在本氏煙中的積累和擴展。
參考文獻
[1]朱賢朝,王彥亭,王智發. 中國煙草病害[M]. 北京:中國農業出版社,2002.
ZHU X C, WANG Y T, WANG Z F. Tobacco disease of China[M]. Beijing: China Agriculture Press, 2002.
[2]馬雪祺,陰艷紅,馮婧嫻,等. 植物NAC轉錄因子研究進展[J]. 植物生理學報,2021,57(12):2225-2234.
MA X Q, YIN Y H, FENG J X, et al. Research progress of NAC transcription factors in plant[J]. Plant Physiology Journal, 2021, 57(12): 2225-2234.
[3]KOU X, LIU C, HAN L, et al. NAC transcription factors play an important role in ethylene biosynthesis, reception and signaling of tomato fruit ripening[J]. Molecular Genetics and Genomics, 2016, 291(3): 1205-1217.
[4]RUSHTON P J, BOKOWIEC M T, HAN S, et al. Tobacco transcription factors: novel insights into transcriptional regulation in the solanaceae[J]. Plant Physiology, 2008, 147(1): 280-295.
[5]ODA-YAMAMIZO C, MITSUDA N, SAKAMOTO S, et al. The NAC transcription factor ANAC046 is a positive regulator of chlorophyll degradation and senescence in Arabidopsis leaves[J]. Scientific Reports, 2016, 6(1): 23609.
[6]LI W, LI X X, CHAO J T, et al. NAC family transcription factors in tobacco and their potential role in regulating leaf senescence[J]. Frontiers in Plant Science, 2018, 9: 1900.
[7]LIU Q L, XU K D, ZHAO L J, et al. Overexpression of a novel Chrysanthemum NAC transcription factor gene enhances salt tolerance in tobacco[J]. Biotechnology Letters, 2011, 33(10): 2073-2082.
[8]JIAN W, ZHENG Y X, YU T T, et al. SlNAC6, A NAC transcription factor, is involved in drought stress response and reproductive process in tomato[J]. Journal of Plant Physiology, 2021, 264: 153483.
[9]GAYRAL M, ARIAS GAGUANCELA O, BASQUEZ E, et al. Multiple ER-to-nucleus stress signaling pathways are activated during Plantago asiatica mosaic virus and Turnip mosaic virus infection in Arabidopsis thaliana[J]. The Plant Journal, 2020, 103(3): 1233-1245.
[10]SEO P J, KIM M J, PARK J Y, et al. Cold activation of a plasma membrane-tethered NAC transcription factor induces a pathogen resistance response in Arabidopsis[J]. The Plant Journal, 2010, 61(4): 661-671.
[11]VOITSIK A M, MUENCH S, DEISING H B, et al. Two recently duplicated maize NAC transcription factor paralogs are induced in response to Colletotrichum graminicola infection[J]. Bmc Plant Biology, 2013, 13(1): 85-100.
[12]LI F F, SUN H J, JIAO Y B, et al. Viral infection-induced endoplasmic reticulum stress and a membrane-associated transcription factor NbNAC089 is involved in resistance to virus in Nicotiana benthamiana[J]. Plant Pathology, 2018, 67: 233-243.
[13]DEMIRER G S, ZHANG H, MATOS J L, et al. High aspect ratio nanomaterials enable delivery of functional genetic material without DNA integration in mature plants[J]. Nature Nanotechnology, 2019, 14(5): 456-464.
[14]THAGUN C, HORII Y, MORI M, et al. Non-transgenic gene modulation via spray delivery of nucleic acid/peptide complexes into plant nuclei and chloroplasts[J]. ACS nano, 2022, 16(3): 3506-3521.
[15]WANG J W, CUNNINGHAM F J, GOH N S, et al. Nanoparticles for protein delivery in planta[J]. Current Opinion in Plant Biology, 2021, 60: 102052.
[16]YANG Y, YANG S, WANG Y, et al. Anti-infective efficacy, cytocompatibility and biocompatibility of a 3D-printed osteoconductive composite scaffold functionalized with quaternized chitosan[J]. Acta Biomaterialia, 2016, 46: 112-128.
[17]MORI T, OKUMURA M, MATSUURA M, et al. Effects of chitin and its derivatives on the proliferation and cytokine production of fibroblasts in vitro[J]. Biomaterials, 1997, 18(13): 947-951.
[18]LI G F, WANG J C, FENG X M, et al. Preparation and testing of quaternized chitosan nanoparticles as gene delivery vehicles[J]. Applied Biochemistry and Biotechnology, 2015, 175(7): 3244-3257.
[19]ZHANG D S, SONG L Y, LIN Z L, et al. HACC-based nanoscale delivery of the NbMLP28 plasmid as a crop protection strategy for viral diseases[J]. ACS Omega, 2021 6(49): 33953-33960.
[20]曲瀟玲,焦裕冰,羅健達,等. 本氏煙NbNAC062的克隆及對馬鈴薯Y病毒侵染的抑制作用[J]. 中國農業科學,2021,54(19):4110-4120.
QU X L, JIAO Y B, LUO J D, et al. Cloning of Nicotiana benthamiana NAC062 and its inhibitory effect on potato virus Y infection[J]. Scientia Agricultura Sinica, 2021, 54(19): 4110-4120.
[21]SUN H J, SHEN L L, QIN Y X, et al. CLC-Nt1 affects potato virus Y infection via regulation of endoplasmic reticulum luminal Ph[J]. New Phytologist, 2018, 220: 539-552.
[22]GAO J P, WANG G H, MA S Y, et al. CRISPR/Cas9-mediated targeted mutagenesis in Nicotiana tabacum[J]. Plant Molecular Biology, 2015, 87(1/2): 99-110.
[23]SUN D, ZHANG X, ZHANG Q, et al. Comparative transcriptome profiling uncovers a Lilium regale NAC transcription factor, LrNAC35, contributing to defence response against cucumber mosaic virus and tobacco mosaic virus[J]. Molecular plant pathology, 2019, 20(12): 1662-1681.
[24]SELTH L A, DOGRA S C, RASHEED M S, et al. A NAC domain protein interacts with tomato leaf curl virus replication accessory protein and enhances viral replication[J]. The Plant Cell, 2005, 17(1): 311-325.
[25]DEMIRER G S, ZHANG H, GOH N S, et al. Carbon nanotube–mediated DNA delivery without transgene integration in intact plants[J]. Nature Protocol, 2019, 14(10): 1-24.
[26]MITTER N, WORRALL E A, ROBINSON K E, et al. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses[J]. Nature Plants, 2017, 3(2): 16207.
[27]陶晴,卞曉軍,張彤,等. DNA水凝膠的制備及應用[J]. 生物工程學報,2021,37(9):3162-3178.
TAO Q, BIAN X J, ZHANG T, et al. Preparation and application of DNA hydrogels: a review[J]. Chinese Journal of Biotechnology, 2021, 37(9): 3162-3178.
[28]HU Q Q, LI H, WANG L H, et al. DNA nanotechnology-enabled drug delivery systems[J]. Chemical Reviews, 2019, 119: 6459-6506.
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
河北科技師范學院學報(2021年1期)2021-05-10 03:34:20
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
電源技術(2017年1期)2017-03-20 13:37:59
海峽科技與產業(2016年3期)2016-05-17 04:32:12
食品界(2016年4期)2016-02-27 07:36:46
應用化工(2014年7期)2014-08-09 09:20:21
