東海海域帶魚的個體繁殖力
2023-06-21 07:45:44玄文丹徐開達(dá)周永東
廣東海洋大學(xué)學(xué)報 2023年3期
謝 玉,朱 凱,玄文丹,徐開達(dá),周永東
(1.浙江海洋大學(xué)海洋與漁業(yè)研究所,浙江 舟山 316022;2.浙江省海洋水產(chǎn)研究所/農(nóng)業(yè)農(nóng)村部重點漁場漁業(yè)資源科學(xué)觀測實驗站/浙江省海洋漁業(yè)資源可持續(xù)利用技術(shù)研究重點實驗室/浙江省海水增養(yǎng)殖重點實驗室,浙江 舟山 316021)
繁殖為魚類生活史的重要組成部分,對維持其群體數(shù)量穩(wěn)定及種群延續(xù)起至關(guān)重要作用[1]。繁殖力變動是物種或種群適應(yīng)環(huán)境變動的結(jié)果,任何影響魚類繁殖力的因素均為影響魚類種群數(shù)量變動的因素[2]。因此,研究魚類繁殖力對于正確估測魚類種群數(shù)量及其變動規(guī)律,開展?jié)O業(yè)管理和合理利用漁業(yè)資源有現(xiàn)實意義[3]。
關(guān)于帶魚(Trichiurus japonicus)個體繁殖力研究雖有一些報道[4-8],但年份大多較為久遠(yuǎn),國內(nèi)關(guān)于帶魚個體繁殖力的報道至少已有18 a[9]。隨著海洋環(huán)境及捕撈強(qiáng)度的變化,帶魚漁獲個體的性成熟逐漸提早、初次性成熟最小肛長趨小[8,10]。這些生物學(xué)特征的改變可能會引起帶魚繁殖生物學(xué)發(fā)生相應(yīng)變化[8]。本研究通過分析東海海域帶魚個體繁殖力與其生物學(xué)特征的關(guān)系,揭示目前東海海域帶魚繁殖生物學(xué)的變動規(guī)律,為今后東海海域帶魚的繁殖力監(jiān)測研究及其資源管理提供理論依據(jù)。
1 材料與方法
1.1 樣品來源與處理
從2020年10月—2021年10月東海海域拖網(wǎng)和帆張網(wǎng)漁獲中隨機(jī)取樣83 尾成熟度為Ⅲ期以上的雌性帶魚,測定其肛長(Anal length,L)、體質(zhì)量(Body mass,m)和卵巢質(zhì)量(Ovary mass,mo),肛長精確到1 mm,體質(zhì)量精確到0.1 g,卵巢質(zhì)量精確到0.000 1 g[11]。卵粒計數(shù)采取質(zhì)量法[12],即取0.2~0.5 g(前、中、后部卵粒混合)卵巢組織,固定于體積分?jǐn)?shù)10%的甲醛溶液中,置于培養(yǎng)皿中,將卵粒均勻離散,用計數(shù)器統(tǒng)計卵粒數(shù)。再隨機(jī)挑選50粒完整卵粒在顯微鏡下測量卵徑,取平均值。
1.2 數(shù)據(jù)處理
計算絕對繁殖力(F,粒)、肛長相對繁殖力(FL,粒/mm)、體質(zhì)量相對繁殖力(Fm,粒/g)、性腺成熟系數(shù)(Gonado somatic index,GSI)、肥滿度(K,g/mm3):
式中,mo為卵巢質(zhì)量;L為肛長,m為體質(zhì)量。
運(yùn)用SPSS 和Excel 2016,根據(jù)文獻(xiàn)[13],對個體繁殖力指標(biāo)(F、FL、Fm)分別與5個生物學(xué)指標(biāo)(L、m、mo、GSI 和K)所組成的15 對因素進(jìn)行相關(guān)分析,分別采用線性函數(shù)、二次函數(shù)、復(fù)合函數(shù)、對數(shù)函數(shù)、指數(shù)函數(shù)、逆函數(shù)和冪函數(shù)對所測定數(shù)據(jù)進(jìn)行擬合,選出最佳擬合回歸方程,用多元逐步回歸分析方法,建立東海海域帶魚個體繁殖力與其生物學(xué)指標(biāo)的多元回歸方程,分析個體繁殖力(F、FL、Fm)與L、m、mo、GSI、K的關(guān)系以及東海帶魚的繁殖特性。
2 結(jié)果與分析
2.1 東海海域帶魚的體型參數(shù)
83 尾雌性東海海域帶魚的肛長為210.0~373.0 mm,平均(289.04±41.00)mm,優(yōu)勢肛長組為230~330 mm,占91.60%,體質(zhì)量 為121.20~694.90 g,平均(375.40±146.95)g,優(yōu)勢體質(zhì)量組為200~600 g,占93.98%(圖1)。對其肛長與體質(zhì)量數(shù)據(jù)進(jìn)行函數(shù)擬合,可知兩者呈冪函數(shù)關(guān)系,關(guān)系式為m=2×10-5L2.9795(R2=0.940,n=83),即83 尾雌性東海海域帶魚的體質(zhì)量隨其肛長的增長而不斷增大,增加速度隨肛長的增長而增大(圖2)。
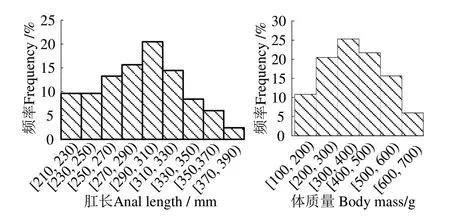
圖1 雌性帶魚個體肛長與體質(zhì)量分布Fig.1 Distribution of anal length and body mass of female Trichiurus japonicus
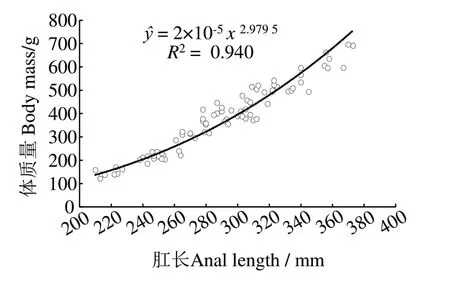
圖2 雌性帶魚肛長與體質(zhì)量的關(guān)系Fig.2 Relationship between anal length and body mass of female Trichiurus japonicus
2.2 個體繁殖力的分布
83 尾雌性帶魚的個體絕對繁殖力為3 001~78 265 粒,平均(27 750.0±16 255.6)粒,其中81.9%集中在10 000~50 000 粒;肛長相對繁殖力12.4~240.1 粒/mm,平均(91.9 ± 45.6)粒/mm,其中95.2%集中在25~175 粒/mm;體質(zhì)量相對繁殖力13.9~152.9 粒/g,平均(72.7 ± 27.6)粒/g,其中93.8%集中在20~120 粒/g。整體上,帶魚F與Fm分布較集中,而FL分布較分散(圖3)。

圖3 東海帶魚個體繁殖力分布Fig.3 Distribution of Trichiurus japonicus individual fecundity in the East China Sea
2.3 個體繁殖力與生物學(xué)指標(biāo)的關(guān)系
2.3.1 相關(guān)分析 魚類個體繁殖力與生物學(xué)指標(biāo)間往往存在某種相關(guān)關(guān)系[13]。相關(guān)分析表明,F(xiàn)、FL與K相關(guān)性不顯著(P>0.05),與其他指標(biāo)間均呈極顯著正相關(guān)(P<0.01),其中F與m相關(guān)系數(shù)最高,L次之,F(xiàn)L與mo相關(guān)系數(shù)最高,m次之;Fm與mo和GSI 呈極顯著正相關(guān)(P<0.01),與K呈顯著負(fù)相關(guān)(P<0.05),與mo的相關(guān)系數(shù)最高,GSI次之,與其他指標(biāo)之間的相關(guān)性不顯著(P>0.05)(表1)。

表1 帶魚個體繁殖力與5個生物學(xué)指標(biāo)間的相關(guān)分析Table 1 Correlation between individual fecundity and 5 biological indicators of Trichiurus japonicus
2.3.2 最佳回歸分析 相關(guān)關(guān)系往往表現(xiàn)為一定的曲線相關(guān)。個體繁殖力(F、FL、Fm)與其生物學(xué)指標(biāo)(L、m、mo、GSI 和K)之間的線性函數(shù)、二次函數(shù)、復(fù)合函數(shù)、對數(shù)函數(shù)、指數(shù)函數(shù)、逆函數(shù)和冪函數(shù)擬合方程見表2。

表2 個體繁殖力與5個生物學(xué)指標(biāo)的最佳擬合方程Table 2 Best fitting equations for individual fecundity and 5 biological indicators
因F、FL與K相關(guān)性不顯著(P>0.05),故東海帶魚F與L、m、mo和GSI分別呈冪函數(shù)關(guān)系、指數(shù)函數(shù)關(guān)系、二次函數(shù)關(guān)系和二次函數(shù)關(guān)系,其中F與L的關(guān)系最為密切;FL與L、m、mo和GSI分別呈冪函數(shù)關(guān)系、二次函數(shù)關(guān)系、對數(shù)函數(shù)關(guān)系和二次函數(shù)關(guān)系,其中FL與mo的關(guān)系最為密切。又因Fm與L和m相關(guān)性不顯著(P>0.05),故Fm與mo、GSI 和K分別呈二次函數(shù)關(guān)系、對數(shù)函數(shù)關(guān)系和二次函數(shù)關(guān)系,其中FL與mo的關(guān)系最為密切。
2.3.3 逐步回歸分析 個體繁殖力與單一生物學(xué)指標(biāo)的擬合方程雖可反映出個體繁殖力與某一生物學(xué)指標(biāo)的局部關(guān)系,但相關(guān)性分析可知,自變量生物學(xué)指標(biāo)L與m(P<0.01),mo與m和GSI 之間均存在極強(qiáng)的多重共線性(P<0.01),為更好探索東海海域帶魚個體繁殖力與其生物學(xué)指標(biāo)之間的關(guān)系,建立個體繁殖力與生物學(xué)指標(biāo)的多元回歸方程:
對于F,m、mo和K為回歸方程的自變量,通過方程(1)可得,F(xiàn)分別隨m和mo的增大而增大,與m和mo呈正相關(guān);F隨K的增大而減小,與K負(fù)相關(guān),與表1 結(jié)果基本一致,不同的是逐步回歸方程舍棄了L、GSI兩個生物學(xué)指標(biāo),這是因為自變量生物學(xué)指標(biāo)m與L、GSI之間存在極強(qiáng)的多重共線性(P<0.01)。
對于FL,mo和L為回歸方程中的自變量,通過方程(2)可得,F(xiàn)L分別隨mo和L的增大而增大,與mo和L呈正相關(guān),與表1 結(jié)果基本吻合,不同的是逐步回歸方程舍棄了m、GSI 兩個生物學(xué)指標(biāo),這是因為自變量生物學(xué)指標(biāo)mo與m、GSI 之間存在極強(qiáng)的多重共線性(P<0.01)。
對于Fm,m和K為回歸方程中的自變量,通過方程(3)可得,F(xiàn)m隨mo的增大而增大,與mo呈正相關(guān);Fm隨K的增大而減小,與K呈負(fù)相關(guān),與表1 結(jié)果基本一致,不同的是逐步回歸方程舍棄了GSI 這一生物學(xué)指標(biāo),這是因為自變量生物學(xué)指標(biāo)mo與GSI 之間存在極強(qiáng)的多重共線性(P<0.01)。
2.4 卵徑分布
從83 尾帶魚樣品中隨機(jī)挑選50 粒完整卵進(jìn)行測量,共測得4 150 個卵徑數(shù)據(jù)。圖4 表明,卵徑為452~2 063 μm,平均(922.94 ± 318.68)μm,其中94.0%的卵徑集中分布在400~1 400 μm,即Ⅲ~Ⅴ期帶魚卵徑僅一個波峰,推測東海海域帶魚為單批次產(chǎn)卵。
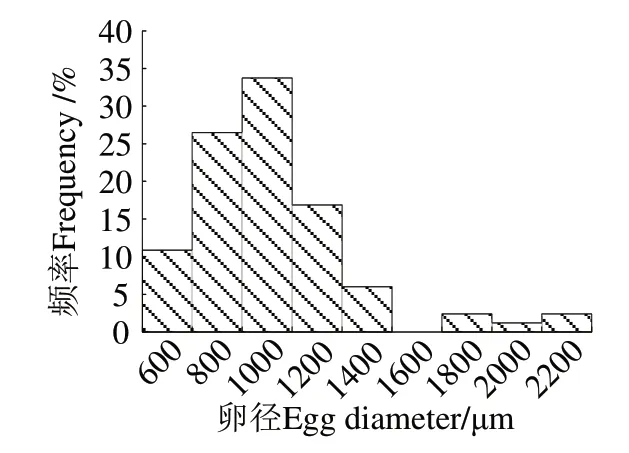
圖4 卵徑頻率分布Fig.4 Frequency distribution of egg diameter
表3可見,性成熟度為Ⅲ期的帶魚有16尾,占整體19.28%,卵徑為452~822 μm,平均(640.63 ±126.14)μm;Ⅳ期有29尾,占整體34.9%,卵徑為570~1 617 μm,平均(835.24±208.22)μm;Ⅴ期有38尾,占整體45.78%,卵徑為589~2 063 μm,平均(1 108.74±331.47)μm。由此可得,隨著性腺成熟度的增長,卵徑隨之增大,且性腺成熟度的優(yōu)勢期為Ⅴ期。
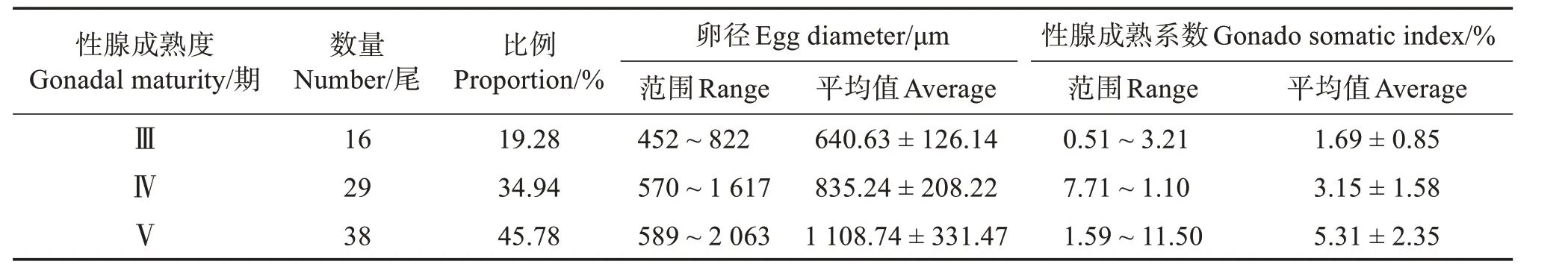
表3 各期成熟度的卵徑和性成熟系數(shù)分布Table 3 Distribution of egg diameter and gonado somatic index at each maturity stage
3 討論
3.1 帶魚生物學(xué)特征
比較不同時期帶魚產(chǎn)卵群體的肛長組成發(fā)現(xiàn),1960—1993 年肛長范圍、均值和優(yōu)勢肛長組整體呈下降趨勢,可能由捕撈壓力的增大以及海洋環(huán)境的變化所致。帶魚為維持種族的繁衍和生存,被迫進(jìn)行自身的適應(yīng)性調(diào)節(jié),具體表現(xiàn)為產(chǎn)卵群體的小型化[10]。而1996—2021 年肛長范圍、均值和優(yōu)勢肛長組整體呈上升趨勢(表4),可能與1995 年后的伏季休漁制度有關(guān),經(jīng)過幾十年的伏季休漁,帶魚產(chǎn)卵群體小型化的趨勢可能得到延緩,具體原因但還需對帶魚產(chǎn)卵群體的肛長組成進(jìn)行繼續(xù)監(jiān)測與分析。
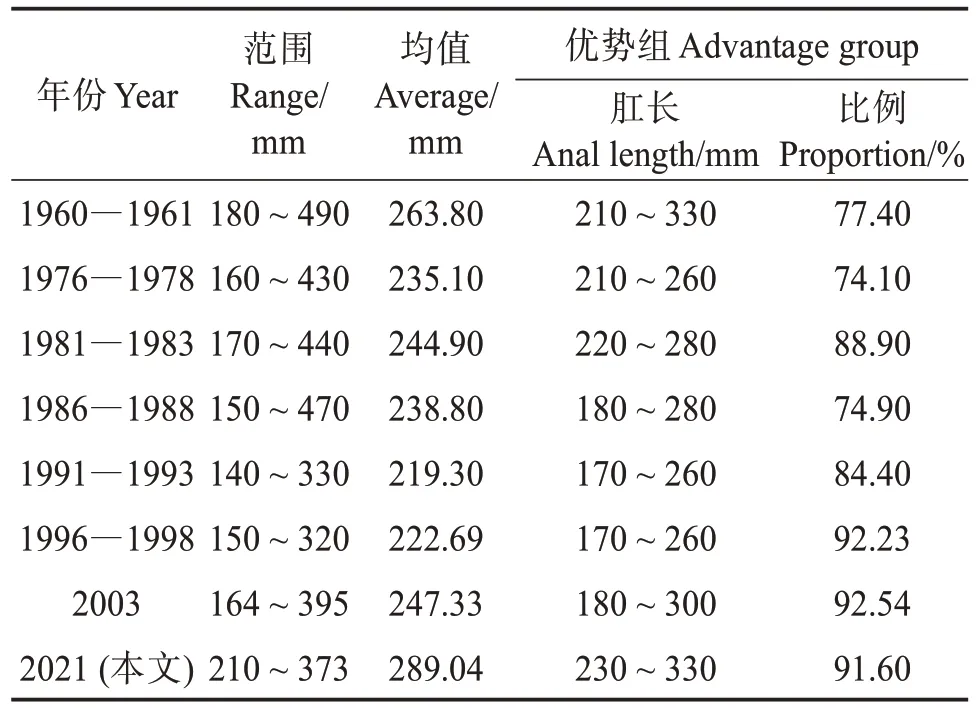
表4 不同時期產(chǎn)卵群體肛長組成Table 4 Anal length composition of spawning population in different periods
3.2 帶魚個體繁殖力
個體相對繁殖力可用來比較規(guī)格不同的不同種或不同種群魚的繁殖力,體現(xiàn)了魚的繁殖策略[2]。個體相對繁殖力高,魚所產(chǎn)卵體積小、數(shù)量多,每個卵成功發(fā)育為成體的機(jī)會少,如大黃魚(Pseudosciaena crocea)、鰻鱺(Anguilla)等,通過產(chǎn)生大量的卵來抵御環(huán)境壓力;個體相對繁殖力低,表示魚所產(chǎn)卵體積大、數(shù)量少,卵含有的卵黃多,有更多的營養(yǎng)供仔魚利用,使每個卵成功發(fā)育為成體的機(jī)會增多,如鮭鱒魚類[11]。帶魚的個體相對繁殖力與大黃魚(P.crocea)[12]、海鰻(Muraenesox cinereus)[13]相比較低,卻與烏蘇里白鮭(Coregonus ussuriensis)[14]、歐白鮭(Coregonus albula)[15]、高白鮭(Coregonus peled)[16]類似,推測帶魚的繁殖策略趨向于通過提高卵的質(zhì)量來保證種的延續(xù)。
對于許多魚類,不同種群或群體繁殖力不同[11]。本研究的帶魚(T.japonicus)個體絕對繁殖力為3 001~78 265 粒,平均27 749.95 粒,而東海海域帶魚(Trichiurus haumela)[9]的個體絕對繁殖力大于本研究,為9 674~159 142 粒,均值為39 233 粒,表明帶魚科魚類同屬不同種之間個體絕對繁殖力相差較大,可能與其性成熟年齡、個體規(guī)格、卵徑、棲息水域環(huán)境及營養(yǎng)狀況等因素有關(guān)[14]。
3.3 個體繁殖力與生物學(xué)指標(biāo)的關(guān)系
魚類個體繁殖力是表征種群動態(tài)的重要指標(biāo),不僅與生物學(xué)特征、外界環(huán)境條件、營養(yǎng)狀況有關(guān),還與生物學(xué)指標(biāo)顯著相關(guān)[2,17]。通過比較不同魚類個體繁殖力與生物學(xué)指標(biāo)之間的關(guān)系,可發(fā)現(xiàn)魚類個體繁殖力與生物學(xué)指標(biāo)間關(guān)系往往隨著魚類種類的不同而有所不同[18]。以常規(guī)生物學(xué)指標(biāo)體長(肛長)、體質(zhì)量、卵巢質(zhì)量和性腺成熟系數(shù)為例,如銀鯧(Stromateoides argenteus)的F和FL與體長呈冪函數(shù)關(guān)系,與體質(zhì)量、卵巢質(zhì)量和性腺成熟系數(shù)呈線性函數(shù)關(guān)系;Fm與卵巢質(zhì)量和性腺成熟系數(shù)呈線性函數(shù)關(guān)系[19]。海鰻(M.cinereus)的F和FL與肛長呈二次函數(shù)關(guān)系,與體質(zhì)量、卵巢質(zhì)量和性腺成熟系數(shù)呈冪函數(shù)關(guān)系;Fm與性腺成熟系數(shù)呈冪函數(shù)關(guān)系[13]。鳳鱭(Coilia mystus)的F和FL與體長、體質(zhì)量、卵巢質(zhì)量和性腺成熟系數(shù)均呈冪函數(shù)關(guān)系;Fm與卵巢質(zhì)量和性腺成熟系數(shù)分別呈指數(shù)函數(shù)和冪函數(shù)關(guān)系[20]。刀鱭(Coilia nasus)的F與體長、體質(zhì)量、卵巢質(zhì)量和性腺成熟系數(shù)分別呈二次函數(shù)、冪函數(shù)、指數(shù)函數(shù)和二次函數(shù)關(guān)系;FL與體長、體質(zhì)量、卵巢質(zhì)量和性腺成熟系數(shù)分別呈冪函數(shù)、冪函數(shù)、二次函數(shù)和二次函數(shù)關(guān)系;Fm與卵巢質(zhì)量和性腺成熟系數(shù)均呈二次函數(shù)關(guān)系[21]。本研究結(jié)果與其他魚類之間既有相同之處,又存在一定區(qū)別:東海海域帶魚(T.japonicus)的Fm與肛長(體長)和體質(zhì)量無相關(guān)性,與銀鯧(S.argenteus)[19]、海鰻(M.cinereus)[13]、鳳鱭(C.mystus)[20]、刀鱭(C.nasus)[21]的結(jié)果一致;東海海域帶魚(T.japonicus)的F與肛長、體質(zhì)量、卵巢質(zhì)量和性腺成熟系數(shù)分別呈冪函數(shù)、指數(shù)函數(shù)、二次函數(shù)和二次函數(shù)關(guān)系,F(xiàn)L與肛長、體質(zhì)量、卵巢質(zhì)量和性腺成熟系數(shù)分別呈冪函數(shù)、二次函數(shù)、對數(shù)函數(shù)和二次函數(shù)關(guān)系,F(xiàn)m與卵巢質(zhì)量和性腺成熟系數(shù)分別呈二次函數(shù)和對數(shù)函數(shù)關(guān)系,與其他魚類[13,19-21]不同。
由表1 可知,F(xiàn)與肥滿度無顯著相關(guān)性,而在多元逐步回歸分析中,F(xiàn)與肥滿度呈顯著負(fù)相關(guān),并舍棄了肛長和性腺成熟系數(shù)這兩個生物學(xué)指標(biāo);FL舍棄了體質(zhì)量和性腺成熟系數(shù)這兩個生物學(xué)指標(biāo);Fm舍棄了性腺成熟系數(shù)這一生物學(xué)指標(biāo)。這是由于各指標(biāo)之間的關(guān)系錯綜復(fù)雜,從單一指標(biāo)看,原本與個體繁殖力不相關(guān)的指標(biāo),在多元逐步回歸分析中由于其它指標(biāo)的關(guān)系而呈現(xiàn)一定的相關(guān)性,同時,原本與個體繁殖力關(guān)系密切的指標(biāo),當(dāng)多元逐步回歸方程中入選指標(biāo)與個體繁殖力的相關(guān)性更顯著時,該單一指標(biāo)可能會被舍棄[20,22]。因此,在分析個體繁殖力與生物學(xué)指標(biāo)的關(guān)系時,除考慮單一指標(biāo)外,還需進(jìn)行多元回歸分析,以準(zhǔn)確說明個體繁殖力與生物學(xué)指標(biāo)之間的本質(zhì)聯(lián)系[22]。
根據(jù)個體繁殖力與生物學(xué)指標(biāo)的關(guān)系可快速計算出個體繁殖力,對預(yù)測種群的繁殖能力、指導(dǎo)漁業(yè)生產(chǎn)具有重要作用[18]。根據(jù)F、FL、Fm的多元逐步回歸方程,東海海域帶魚的F主要受體質(zhì)量、卵巢質(zhì)量和肥滿度的影響,F(xiàn)L主要受卵巢質(zhì)量和肛長的影響,F(xiàn)m主要受卵巢質(zhì)量和肥滿度的影響,故帶魚個體繁殖力與卵巢質(zhì)量的關(guān)系最為密切,因而適宜用個體繁殖力與卵巢質(zhì)量的關(guān)系對東海海域帶魚種群繁殖力進(jìn)行預(yù)測,以了解其種群動態(tài)。這與長江口鳳鱭產(chǎn)卵群體繁殖力特征研究結(jié)果[23]相同。
3.4 產(chǎn)卵類型分析
魚類產(chǎn)卵類型可分為一次性產(chǎn)卵和分批產(chǎn)卵,有關(guān)帶魚的產(chǎn)卵類型,目前尚存諸多爭議,有些研究認(rèn)為帶魚卵巢是一次成熟[5],也有學(xué)者根據(jù)生殖季節(jié)的卵巢組織學(xué)觀察,認(rèn)為帶魚屬多次排卵[6]。在魚類繁殖生物學(xué)中,常用卵徑頻率分布判斷魚類的產(chǎn)卵類型[7],本研究并未發(fā)現(xiàn)東海帶魚卵巢內(nèi)已經(jīng)形成一定數(shù)量規(guī)模的小卵徑魚卵,且卵徑分布圖近似單峰型,故推測本研究采集的性成熟帶魚應(yīng)為一次性產(chǎn)卵的魚類。
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
現(xiàn)代畜牧科技(2021年4期)2021-07-21 06:13:00
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
福建基礎(chǔ)教育研究(2019年1期)2019-09-10 07:22:44
中國生殖健康(2019年2期)2019-08-23 08:12:08
福建基礎(chǔ)教育研究(2019年1期)2019-05-28 08:39:49
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
數(shù)學(xué)物理學(xué)報(2017年2期)2017-06-05 09:12:30
汽車觀察(2016年3期)2016-02-28 13:16:26
醫(yī)學(xué)研究雜志(2015年11期)2015-06-10 06:44:03
