利用CRISPR/Cas9 技術(shù)定向編輯加工番茄 SlACS2
2023-06-26 02:02:52劉江娜張西英白云鳳張愛萍
西北農(nóng)業(yè)學報 2023年6期
劉江娜,張西英,白云鳳,張愛萍
(新疆生產(chǎn)建設(shè)兵團 第六師農(nóng)業(yè)科學研究所,新疆五家渠 832100)
番茄(Solanumlycopersicum)屬于呼吸躍變型果實,在成熟過程中乙烯產(chǎn)量驟增,果實容易成熟過度而變軟,致使耐壓性能變差、貯藏期縮短、抗病原菌能力降低,嚴重影響到生產(chǎn)、貯運及加工品質(zhì)。因此,探索能夠抑制加工番茄果實過度軟化、提高耐貯性、便利儲存運輸?shù)姆椒@得尤為重要[1-3]。 利用基因工程手段調(diào)節(jié)番茄過熟軟化、改善耐貯性取得一定進展, 途徑主要有兩條:(1)通過抑制乙烯合成,來提高果實耐受“成熟過渡” 的能力。乙烯的生物合成遵循甲硫氨酸 (Methionine,Met)→S-腺苷甲硫氨酸(S-Adenosyl methionine,SAM)→1- 氨基環(huán)丙烷 1-羧酸(1-aminocyclopropane-1-carboxylic acid,ACC)→乙烯的途徑。將 ACC 合酶(ACC synthase,ACS)[4-5]和ACC氧化酶(ACC oxidase,ACO)/乙烯形成酶(ethylene-forming enzyme,EFE) 反義基因[6-7]或 RNAi 基因[8-9]導入番茄,均可抑制乙烯合成,并已培育出耐貯運基因工程新品種‘華番 1 號’[10]。(2)抑制細胞壁果膠的降解。多聚半乳糖醛酸酶(polygalacturonase,PG) 是一種細胞壁蛋白,能將果實中果膠的 主要成分多聚半乳糖醛酸降解為半乳糖醛酸,使細胞結(jié)構(gòu)解體,導致果實軟化。把 PG 反義基因?qū)敕?能抑制 PG 表達,提高番茄硬度,延緩過熟軟化[11]。但因其是在植物原有基因組上整合了新序列,易引起民眾的安全性 疑慮,增加轉(zhuǎn)基因作物評價和應用成本。
近年來,以多種新型高效的DNA靶向內(nèi)切酶為基礎(chǔ)建立的基因組編輯技術(shù),是對植物自身的基因進行定點修飾,提高了育種效率,規(guī)避了常規(guī)轉(zhuǎn)基因可能產(chǎn)生的不良效應。基因編輯可分為鋅指核酸酶(Zinc-finger nuclease,ZFN)、類轉(zhuǎn)錄激活因子效應物核酸酶(trans cription activator-like effector nuclease,TALEN)和CRISPR/Cas(clustered regulatory interspaced short palindromic repeat(CRISPR)/CRISPR-associated(Cas))技術(shù)。ZFN和 TALEN是利用蛋白質(zhì)識別特定的基因組靶位點,針對每一個突變位點需要構(gòu)建兩個相應的核酸酶,操作繁瑣; CRISPR/Cas是利用更簡單的核苷酸互補配對方式識別特定的基因組靶位點,比ZFN和TALEN編輯效率高,構(gòu)建更簡單。CRISPR/Cas9 系統(tǒng)的首篇報道發(fā)表于《科學》雜志上,研究發(fā)現(xiàn) CRISPR/Cas9 系統(tǒng)可以在體外切割雙鏈 DNA,實現(xiàn)了在活細胞中進行基因編輯[12]。隨后因其操作較為簡單、效率高、應用更為廣泛,相繼在甜橙[13-15]、葡萄[16-18]、獼猴桃[19]、芥藍[20]、甘藍[21]等園藝作物中成功實現(xiàn)了基因編輯。目前,學者利用CRISPR/Cas9技術(shù)已對番茄的多個基因,如轉(zhuǎn)錄因子RIN[22]、RNA編輯因子SlORRM4[23]、果實軟化相關(guān)基因α-Man[24]及生長相關(guān)的DELLA基因[25]進行了編輯,也為提高番茄耐貯性提供了新思路[26]。
ACS是乙烯合成的限速酶。番茄是呼吸躍變型果實,其ACS(SlACS)由多基因家族編碼,SlACS1A和SlACS6主導系統(tǒng)Ⅰ乙烯的合成,負責番茄果實在呼吸躍變前產(chǎn)生基礎(chǔ)水平的少量乙烯,SlACS2和SlACS4主導系統(tǒng)Ⅱ乙烯的合成,在果實成熟過程中伴隨呼吸躍變產(chǎn)生大量乙烯,催熟果實。系統(tǒng)Ⅱ乙烯具有自催化機制,使得番茄成熟過程難以控制,造成果實過熟軟化、腐爛變質(zhì)。番茄1-氨基環(huán)丙烷1-羧酸ACC(1-aminocyc lopropane- 1-carboxylic acid)是合成乙烯的直接前體,由ACC合酶(ACC synthase)SlACS催化生成。番茄SlACS2催化系統(tǒng)Ⅱ乙烯合成前體ACC的生成,是系統(tǒng)Ⅱ乙烯合成的重要限速酶。
利用CRISPR/Cas9技術(shù)敲除SlACS2基因,通過農(nóng)桿菌介導的遺傳轉(zhuǎn)化獲得ACS2突變體,不僅有利于研究SlACS2基因?qū)Ψ压麑嵆墒旒百A藏性能等方面的影響,進而培育耐貯藏品種,也為改良番茄的其他性狀建立新途徑,具有重要的生物學意義及商業(yè)價值。
1 材料與方法
1.1 試驗材料、載體及菌株
1.1.1 植物材料、菌株及試劑 試驗所用材料為第六師農(nóng)業(yè)科學研究所自育加工番茄(Solanumlycopersicum)品種‘新番72號’(650444),以期子葉為外植體,利用農(nóng)桿菌介導的遺傳轉(zhuǎn)化方法進行轉(zhuǎn)化。
大腸桿菌(Escherichiacoli)DH5α 與根癌農(nóng)桿菌(Agrobacterium tumefaciens)EHA105 由中國農(nóng)業(yè)科學院蔬菜花卉研究所農(nóng)業(yè)農(nóng)村部園藝作物生物學與種質(zhì)創(chuàng)制重點實驗室保存,中間載體SIU6b::sgRNA購自GenScript 公司、PCAMBIA2300,PCB-DT1TA-SIU6P載體、表達載體sgRNA骨架載體pCAMBIA2300_SlU6p_sgRNA(CP185)和CRISPR/Cas9雙元載體(CP178)由中國農(nóng)科院蔬菜花卉研究所崔霞研究員惠贈。
1.1.2 試劑 限制性內(nèi)切酶Eco31Ⅰ、AscⅠ、XbaⅠ、SalⅠ、EcoRⅠ和 T4連接酶購于 NEB (New England Biolabs) 公司; KOD Fx DNA 聚合酶購于 Toyobo,日本; 凝膠回收和質(zhì)粒提取試劑盒、DNA Marker、逆轉(zhuǎn)錄試劑盒等均購于寶生物工程(大連) 有限公司(TaKaRa) ; PCR 引物(表 1) 、序列測定等由華大基因完成。
1.2 方 法
1.2.1 特異性靶向SlACS2基因的sgRNA設(shè)計與合成 番茄SlACS2基因的克隆:根據(jù)NCBI已報道的SlACS2序列(GenBank ACCESSION:NM_001247249.2),利用Primer5.0 設(shè)計合成特異性引物SlACS2-F/SlACS2-R(表1),采用天根植物總RNA提取試劑盒提取番茄總RNA,利用非變性的1.2%瓊脂糖凝膠,電泳檢測所提RNA質(zhì)量,以‘新番72’號總RNA為模板,采用寶生物PrimeScriptTMRT-PCR Kit試劑盒反轉(zhuǎn)錄合成cDNA第一鏈,以合成的cDNA鏈為模板對SlACS2基因進行擴增,擴增產(chǎn)物以1.0%瓊脂糖凝膠電泳進行檢測,對獲得的目的條帶,用PCR產(chǎn)物純化試劑盒進行回收,并送公司測序。對測序結(jié)果進行分析,明確其序列。

表1 本研究中所用引物Table 1 Primers used in this study
靶向番茄SlACS2基因的sgRNA的設(shè)計:SlACS2-sgRNA的設(shè)計,具體步驟為: 打開靶點預測網(wǎng)站(https:/ /crispr.dbcls.jp/),利用靶位點在線設(shè)計工具“CRISPRdirect”,在文本框中輸入基因序列,選擇番茄物種,即可得出該基因序列所有的靶位點,綜合考慮各項參數(shù)后選取幾組靶位點。 長19nt的寡核苷酸sgRNA核心序列按GG(N)19NGG序列進行設(shè)計。
構(gòu)建sgRNA的寡聚核苷酸: 根據(jù)選擇的sgRNA核心序列,設(shè)計正義鏈引物和反義鏈引物,兩條引物的5′端添加含有核酸內(nèi)切酶AarⅠ識別位點的序列ATATCACCTGCACACTTTGG,以與載體質(zhì)粒的黏性末端互補。正義鏈引物在添加序列的3′端為編輯位點197~215的序列GGATTAAGAGAAACCCAAA,編輯位點的3′端添加GTTTCAGAGCTATGCTGGAA (SEQ ID NO:5);反義鏈引物在添加序列的3′端為編輯位點276~254的反向互補序列TTGCCAAC- TTTCAAGATTA,編輯位點的3′端添加CCAAACTACACTGTTAGATTC(SEQ ID NO:6)。
1.2.2 植物表達載體的構(gòu)建 以CP185為模板,用正義鏈引物(SEQ ID NO:5)和反義鏈引物(SEQ ID NO:6)進行PCR擴增,得到含有雙sgRNA的DNA片段并連接至pEasy-blunt,測序無誤后,用AarI酶切連接至CP178載體上,形成植物表達載體pSlACS2-DsgRNA(圖1)。植物表達載體上的Cas9基因受35S驅(qū)動,兩條sgRNA基因均受番茄U6啟動子驅(qū)動。構(gòu)建好的植物表達載體通過熱激法轉(zhuǎn)化到大腸桿菌DH5 菌株中擴增,通過測序驗證載體序列正確,再通過凍融法轉(zhuǎn)化農(nóng)桿菌LBA4404菌株,以備侵染利用。

圖1 植物表達載體pSlACS2-DsgRNA的構(gòu)建Fig.1 Construction of plant expression vector pSlACS2-DSGRNA
1.2.3 農(nóng)桿菌介導的加工番茄‘新番72號’遺傳轉(zhuǎn)化及抗性植株的獲得 稱取0.6 g籽粒飽滿的‘新番72號’種子,在超凈工作臺內(nèi)用75%的酒精處理30 s,無菌水沖洗2遍,倒掉無菌水再加入10%的漂白水,放置在搖床上晃動1 h左右,至種子泛白,取出種子用無菌水沖洗5遍,放置 在 4 ℃冰箱中冷藏12 h,取出處理好的種子(盡量控干多余水分)接種至1/2MS培養(yǎng)基中(不含蔗糖),每瓶接種50~60粒種子,培養(yǎng)溫度為25~28 ℃、光照度1 800 lx、光照時間16 h,培養(yǎng)6 d獲得無菌苗,剪取6 d苗齡的加工番茄無菌苗子葉為外置體在預培養(yǎng)基A1中預培養(yǎng)1 d(黑暗條件)。
取-80 ℃條件下保存的含有pSlACS2-DsgRNA質(zhì)粒的農(nóng)桿菌甘油菌,在固體YEB培養(yǎng)基平板上劃線(Rif 50 mg/L 與Kan 50 mg/L),倒置于28 ℃條件下培養(yǎng)48 h,挑取含目的片段的農(nóng)桿菌單菌落,接種于5 mL 的YEB 液體培養(yǎng)基中(含有50 mg/mL Kan和100 mg/mL Rif),置于搖床內(nèi)200 r/min培養(yǎng)至OD≈1.0,取1 mL轉(zhuǎn)入相同YEB培養(yǎng)基50 mL,培養(yǎng)至OD600=0.7,將預培養(yǎng)后的外植體在農(nóng)桿菌懸浮液中輕微震蕩15 min,之后,用濾紙吸干多余菌液,28 ℃共培養(yǎng)2 d(黑暗條件),轉(zhuǎn)入抗性培養(yǎng)基A2(MS+玉米素(2 mg/L)+吲哚乙酸(1 mg/L)+卡那霉素(100 mg/L)+特美汀(300 mg/L)進行抗性篩選培養(yǎng),每隔10~15 d將外植體轉(zhuǎn)移至新的篩選培養(yǎng)基進行篩選培養(yǎng),將獲得的抗性不定芽轉(zhuǎn)移至不定芽伸長培養(yǎng)基A3繼續(xù)培養(yǎng)15~20 d,轉(zhuǎn)入生根培養(yǎng)基A4中進行生根培養(yǎng)至成株。
培養(yǎng)基具體配方如下:A1預培養(yǎng)培養(yǎng)基的配方為MS+玉米素(1 mg/L)+吲哚乙酸 (1 mg/L)。A2分化培養(yǎng)基的配方為MS+玉米素(2 mg/L)+吲哚乙酸(1 mg/L)+卡那霉素(100 mg/L)+特美汀(300 mg/L)。A3不定芽伸長培養(yǎng)基配方MS+玉米素(2 mg/L)+吲哚乙酸(1 mg/L)+卡那霉素(50 mg/L)+特美汀(300 mg/L)。A4生根培養(yǎng)基的配方為MS+吲哚乙酸(1 mg/L)+卡那霉素(50 mg/L)+特美汀(300 mg/L)。
1.2.4 轉(zhuǎn)基因植株的分子檢測 以加工番茄‘新番72號’野生型和抗性再生植株幼嫩葉片為材料,使用天根植物基因組DNA提取試劑盒提取T0代植株葉片DNA,利用Cas9特異引物擴增載體特異片段,擴增出目的片段的即為轉(zhuǎn)基因陽性植株。
2 結(jié)果與分析
2.1 SlACS2基因的克隆與植物表達載體的構(gòu)建
以加工番茄‘新番72號’RNA為模板,以 SlACS2-F/SlACS2-R為引物,進行RT-PCR 擴增,對其擴增產(chǎn)物進行測序獲得SlACS2的cDNA序列,大小為1 458 bp。將測序結(jié)果與網(wǎng)上公布的序列,通過DNAstar進行比對分析二者相似度達到100%,證明克隆的基因是所要獲得的目標基因。對其gDNA比對結(jié)果發(fā)現(xiàn),SlACS2由3個內(nèi)含子和4個外顯子組成,根據(jù)CRISPR-Cas9靶點設(shè)計原則,利用CRISPRdirect (https://crispr.dbcls.jp/),在SlACS2第2外顯子上篩選到2個靶位點(圖2-A),sgRNA靶位點序列1和2在番茄基因組1號染色體,之后設(shè)計一對包含 2 個 Target 和 Aar I 酶切位點的引物(SEQ ID NO:5+ SEQ ID NO:6),以 CP185載體為模板,擴增 sg RNA1_U6-26t_Sl U6p_sg RNA2 片段并連接至pEasy-blunt,測序無誤后,然后用 Aar I (Thermo, ER1581)酶切sg RNA1_U6-26t_Sl U6p_sg RNA2 片段和終載體CP178。最后用T4DNA 連接酶(NEB,M0202)進行連接, 將連接成功的sg RNA1_U6-26t_Sl U6p_sg RNA2 表達盒組裝到CP178載體上,構(gòu)建成終載體pSlACS2-DsgRNA(圖2-B)。菌液 PCR 和測序獲得正確載體,并用于加工番茄的遺傳轉(zhuǎn)化。表1為構(gòu)建 CRISPR/Cas9 載體所用的引物。
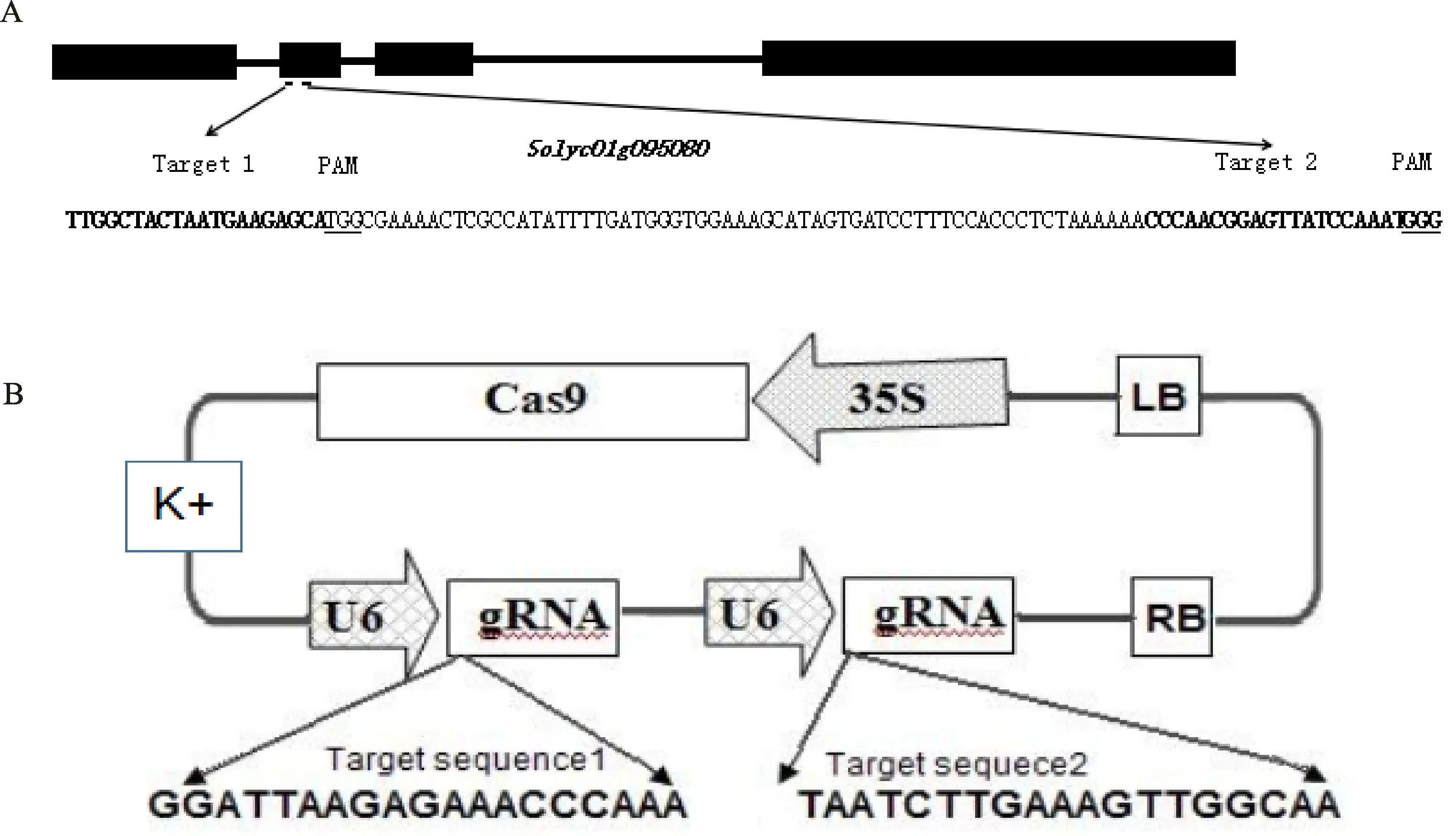
A.雙gRNA靶位點在 SlACS2中的位置;B.pSlACS2-DsgRNA-T1質(zhì)粒T-DNA區(qū)示意圖
2.2 農(nóng)桿菌介導的加工番茄‘新番72號’子葉遺傳轉(zhuǎn)化
以加工番茄子葉為外植體,經(jīng)過種子萌發(fā)后(圖3-A),取無菌培養(yǎng)的番茄子葉剪切后于預培養(yǎng)培養(yǎng)基上預培養(yǎng)1 d(黑暗條件),再置于含基因編輯表達載體 pSlACS2-DsgRNA的農(nóng)桿菌28 ℃共培養(yǎng)2 d(黑暗條件)(圖3-B),轉(zhuǎn)入抗性培養(yǎng)基進行抗性篩選培養(yǎng),獲得抗性愈傷組織(圖3-C),培養(yǎng)20~45 d誘導出抗性不定芽(圖3-D),再在芽伸長培養(yǎng)基中篩選培養(yǎng)15 d(圖3-E),最終轉(zhuǎn)入生根培養(yǎng)基中進行生根培養(yǎng),獲得卡那抗性的再生植株。
2.3 T0代陽性植株分子鑒定
對獲得的31株抗性再生植株編號,每株取 0.2 g嫩葉提取基因組總DNA后,利用特異引物Cas9-F和Cas9-R進行PCR 擴增,以植物編輯表達載體質(zhì)粒為陽性對照,結(jié)果表明,其中22株擴增出了大小為841 bp(與陽性對照一致)的條帶,野生株未擴增出目的條帶(圖4),初步表明已將Cas9基因轉(zhuǎn)化到受體細胞中。

M.DL2000 maker;WT.野生型;P.陽性質(zhì)粒;1~31.抗性植株編號
2.4 SlACS2突變體植株的基因突變類型鑒定
為進一步獲得純合突變植株,提取T1代植株的葉片DNA,以(xp3196+xp3199)作為靶位點上下游特異引物進行PCR 擴增,然后將對應的PCR產(chǎn)物回收轉(zhuǎn)化大腸桿菌DH5 ,進行測序。使用 DNAMAN 對測序結(jié)果進行比對分析,結(jié)果發(fā)現(xiàn)共有6種突變類型,主要以堿基的缺失和插入為主,未發(fā)現(xiàn)有堿基替換,除GE1和GE3僅在一處靶位點發(fā)生堿基的缺失外,其余均同時在2個編輯位點發(fā)生堿基的缺失或插入,具體編輯類型如圖5-A所示,其中GE1由于在靶位點2處缺失7個核苷酸,造成其在第113位和114位氨基酸發(fā)生缺失,同時自第116位氨基酸后發(fā)生移碼(如圖5-B);GE2因在在靶位點1處缺失1個核苷酸致使自第98至112位氨基酸發(fā)生移碼,同時靶位點2處缺失7個核苷酸,造成其在第113位和114位氨基酸發(fā)生缺失,同時自第116位氨基酸后發(fā)生移碼;GE3在靶位點1處缺失4個堿基,致使GE4在靶位點1處缺失5個核苷酸同時靶位點2處缺失7個核苷酸,GE5在靶位點1處有1個單堿基的插入同時靶位點2處缺失7個核苷酸,GE6在靶位點1處缺失9個核苷酸同時靶位點2處缺失7個核苷酸。
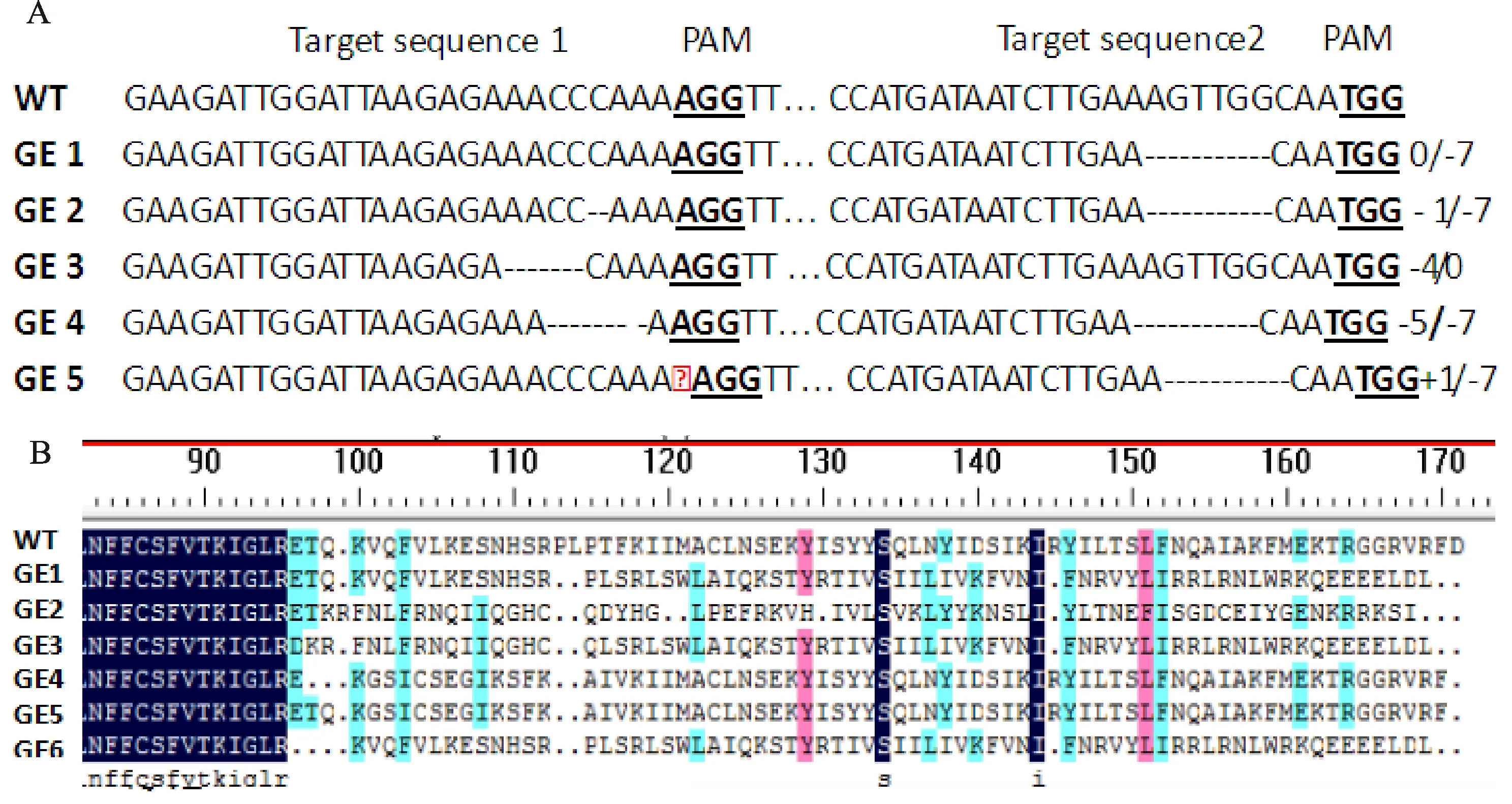
A. SlACS2突變體與野生型核苷酸序列比對; B. SlACS2突變體與野生型氨基酸序列比對A.Comparison of SlACS2 mutants with wild-type nucleotide sequences;B.Comparison of SlACS2 mutants with wild-type amino acids圖5 SlACS2突變體與野生型核苷酸及氨基酸序列比對分析Fig.5 Comparative analysis of nucleotide and amino acid sequences between SlACS2 mutant and wild-type
3 討 論
ACS作為乙烯形成的關(guān)鍵酶[27],由多基因家族編碼。通過抑制ACS的表達抑制乙烯的產(chǎn)生,從而調(diào)控果實成熟的研究方法,多集中于反義RNA的研究[28-29],其效果顯著。將反義ACS基因轉(zhuǎn)入番茄子葉,得到了乙烯合成受抑制的轉(zhuǎn)基因番茄,在沒有乙烯合成的情況下,番茄果實的成熟衰老受抑制,果實不變紅,硬度較大,耐失水,在室溫下可貯藏2~3個月[30];Oeller等[4]將ACS的基因ACS2反向插入載體并轉(zhuǎn)化番茄后,大大降低了乙烯合成量,成功控制了果實的成熟;熊愛生等[31]將ACC氧化酶和ACC合成酶(ACS)反義 RNA融合基因?qū)敕?經(jīng)過4代選擇獲得了表現(xiàn)良好的轉(zhuǎn)基因材料,乙烯釋放量僅為野生型材料的9.5%, 達到阻遏番茄果實中乙烯的形成的目的,顯著地提高番茄果實的耐貯存性能;生吉萍等[32]發(fā)現(xiàn)轉(zhuǎn)翻譯ACS的番茄果實在沒有乙烯存在的情況下繼續(xù)完成生長發(fā)育,并緩慢進入成熟衰老階段;綜上所述,雖然將外源基因?qū)敕迅纳颇唾A性取得了一定的進展,比常規(guī)育種縮短了育種周期,拓寬了基因來源,但由于番茄基因組上整合的是外源基因,易使民眾產(chǎn)生安全性疑慮,影響了基因工程育種效率及推廣應用。
CRISPR-Cas9 系統(tǒng)對目標基因進行精確的原位編輯后,不必再通過轉(zhuǎn)基因調(diào)控目標基因功能。由于sgRNA-Cas9基因的整合位點和編輯位點是分離的,因此可通過遺傳分離剔除sgRNA-Cas9基因,比常規(guī)轉(zhuǎn)基因技術(shù)安全可靠;編輯修飾的基因只有少數(shù)核苷酸改變,與自然突變或人工理化誘變本質(zhì)相同,而在效率和可控性方面則遠遠優(yōu)于誘變育種。番茄基因組較小,基因組測序已經(jīng)完成(Tomato Genome Consortium,2012),遺傳轉(zhuǎn)化體系也已成熟。迄今利用 CRISPR-Cas9 系統(tǒng)對番茄的ARGONAUTE7基因進行了編輯,獲得了復葉畸形突變體[33],但鮮見改良番茄農(nóng)藝性狀的報道。番茄是二倍體自花授粉作物,利用 CRISPR-Cas9 系統(tǒng)原位定點修飾乙烯代謝相關(guān)基因以遲滯番茄過熟腐爛,并通過自交分離剔除sgRNA-Cas9基因,獲得無轉(zhuǎn)基因序列的耐貯運新品種,無疑對新疆番茄的生產(chǎn)加工業(yè)具有重要的應用價值,也將為利用 CRISPR-Cas9 系統(tǒng)改良番茄其他農(nóng)藝性狀、培育無轉(zhuǎn)基因序列的新品種探索新途徑。
利用 CRISPR/Cas9技術(shù)進行單個位點的編輯,如果編輯位點發(fā)生堿基替換,而替換的堿基僅僅造成同義突變,該編輯則是無效編輯;如果編輯位點發(fā)生了堿基插入或缺失,插入或缺失的堿基數(shù)為3的倍數(shù),盡管會造成氨基酸數(shù)量的改變,但不一定會造成移碼突變,編碼的蛋白質(zhì)仍然保持原有功能。為了增加基因編輯的有效性,本研究采用了雙位點編輯策略,2個編輯位點的PAM之間相隔79 bp。在產(chǎn)生的突變體中,有單位點突變,也有雙位點突變,包括雙位點均發(fā)生堿基缺失,但兩個位點間的序列沒有完全刪除。先前有文獻報道,兩個基因編輯位點之間相隔66 bp,得到了中間片段刪除的突變類型[33]。大片段序列的刪除,無疑會減少翻譯失活蛋白的能量消耗。兩個基因編輯位點之間的序列能否刪除,除了與位點之間的距離相關(guān)外,是否還有其他影響因素,尚需進一步探討。
