藍莓葉斑病病原菌鑒定及生物學特性
2023-06-30 12:50:23羅其鑫楊瑩李思尚曉靜侯瑞
江蘇農業科學 2023年5期
羅其鑫 楊瑩 李思 尚曉靜 侯瑞

摘要:為明確貴州省黔東南苗族侗族自治州麻江縣藍莓葉斑病病原菌種類及其生物學特性,采用組織分離法獲得了代表性病原菌株Y2,科赫氏法則證明Y2是引起藍莓葉斑病的病原菌。形態學和分子生物學綜合鑒定菌株Y2為鏈格孢菌(Alternaria alternate)。生物學特性研究結果表明,病原菌Y2在PDA培養條件下,氣生菌絲較發達,菌絲呈白色、密集,基質背面呈淡黃色,分生孢子單生,褐色,卵圓形、倒棒形,有縱橫隔膜。該菌菌絲生長的最佳培養基為察氏培養基,產孢最適培養基為WA培養基;菌絲生長的最適溫度為25 ℃,產孢最適溫度為5 ℃;菌絲對碳源葡萄糖利用效果最好,但麥芽糖能更好地促進Y2產孢;菌絲生長最適氮源為蛋白胨,產孢最適氮源為硫酸銨;全光照適宜菌絲生長,但全黑暗促進Y2產孢;菌絲生長和產孢最適pH值為7。通過Y2菌株生物學特性研究,能夠為藍莓葉斑病防治提供理論依據。
關鍵詞:藍莓;葉斑病;鏈格孢菌;生物學特性
中圖分類號:S436.639文獻標志碼:A
文章編號:1002-1302(2023)05-0146-09
藍莓學名篤斯越橘(Vaccinium uliginosum L.),屬杜鵑花科(Ericaceae)越橘亞科(Vaccination)越橘屬(Vaccinium)植物。藍莓漿果中含有豐富的花青素、鞣花酸等活性物質,可以抗衰老、減少癌癥等的發病率[1-3]。隨著我國藍莓種植規模的逐漸擴大,日趨突出的藍莓病害導致藍莓產業發展受阻[4]。目前已報道的藍莓常見病害主要有藍莓葉斑病[5]、枝枯病[6]、根癌病[7]、灰霉病[8]、間座殼芽枯病[9]和枝干潰瘍病[10]等。藍莓葉斑病已成為藍莓常見病害,西澳大利亞州報道了在藍莓上由極細鏈格孢菌(Alternaria tenuissima)引起的葉斑病[11],Norman等報道了由青枯菌(Ralstonia solanacearum)引起的藍莓青枯病[12],國內研究報道的有由根癌土壤桿菌(Agrobacterium tumefaciens)引起的藍莓根癌病[7]、灰葡萄孢菌(Botrytis cinerea)引起的藍莓灰霉病[13]和極細鏈格孢引起的藍莓葉斑病[4]。藍莓葉斑病致病菌復雜多樣,嚴雪瑞等研究發現引起藍莓葉穿孔病的致病菌為鏈格孢菌(Alternaria alternate)[14],張國輝等在貴州省黔東南州臺江縣發現了巨腔莖點霉(Phoma macrostoma)為病原的藍莓葉斑病[15]。明確致病菌種類及其生物學特性有助于該病害后續的綜合防治。
由于藍莓葉斑病病原菌的多樣性,導致不同地域葉斑病致病菌的生物學特性不同,使藍莓葉斑病的防治較為困難。本試驗對引起貴州省黔東南苗族侗族自治州麻江縣藍莓葉斑病的病原菌進行鑒定和生物學特性研究,明確其菌絲生長的最適條件和產孢情況,研究結果可為防治藍莓葉斑病提供理論依據。
1 材料與方法
1.1 試驗材料
供試發病藍莓葉片在貴州省黔東南苗族侗族自治州麻江縣藍莓栽培生產基地采集,塑封袋封裝帶回實驗室后于4 ℃冰箱保存備用。
1.2 試驗方法
1.2.1 藍莓葉斑病病原菌的分離及致病性測定
將分離純化后的菌株Y2接種至健康葉片上,致使健康藍莓葉片感病,結果發病藍莓的葉片上應能得到與初始接種的菌株性狀一致的菌落,對照組不會得到與初始菌株一致的菌落。接種前先用75%乙醇對葉片表面進行消毒,接著用無菌水反復沖洗3次,移入5%(體積分數) NaClO溶液中滅菌2 min,用無菌水清洗后晾干備用。
離體接種:用消過毒的接種針在消過毒的健康藍莓葉片表面避開葉脈刺4個小孔,于小孔上貼直徑5 mm的菌餅,覆上滅菌的濕脫脂棉,再套保鮮袋保濕,試驗每張藍莓葉片接種2個菌餅,每個菌株處理5個重復,1個對照組。
1.2.2 藍莓葉斑病病原菌的鑒定
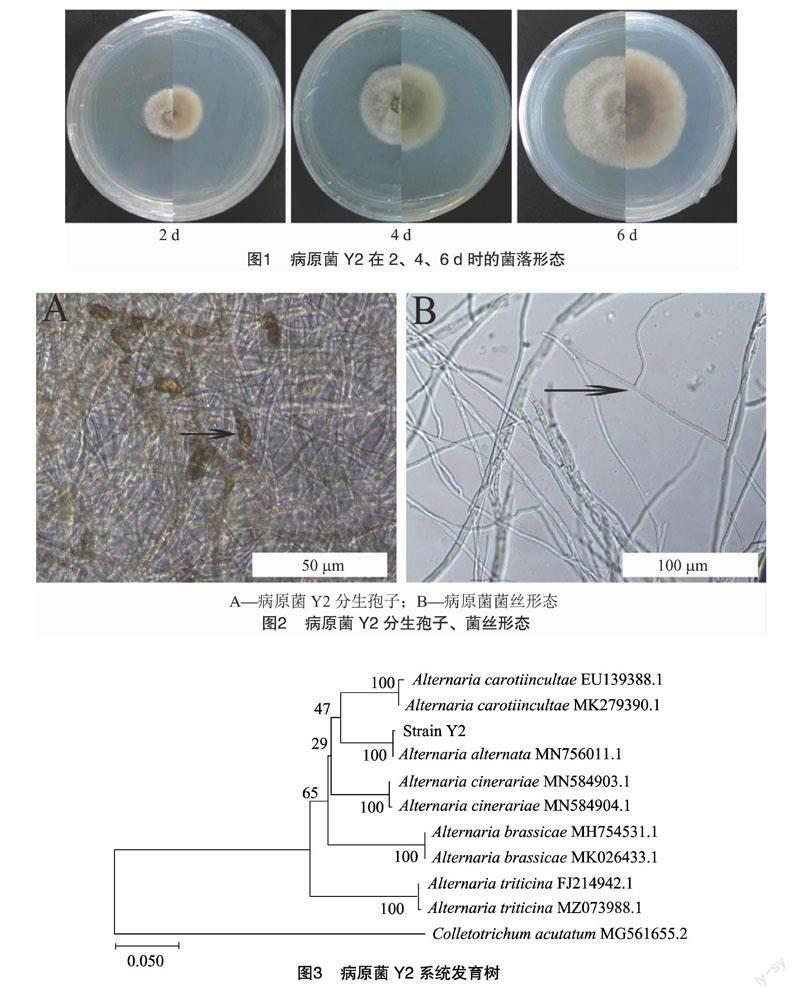
形態學鑒定:超凈操作臺中,用內徑為5 mm的打孔器在2種病原菌Y2的菌落邊緣打出菌餅,將Y2菌株的菌餅接種在PDA培養基上并置于28 ℃培養箱中2、4、6 d進行培養皿觀察拍照,觀察其菌落特點(菌絲形態、顏色、基質顏色、菌絲生長速度、菌絲密度等)。在光學顯微鏡下參照《真菌鑒定手冊》對菌絲的分生孢子梗及分生孢子進行形態學觀察,記錄其形態特征并測量分生孢子大小。
分子生物學鑒定:病原菌Y2用PDA平板進行擴繁后,刮取PDA平板上活化好的Y2菌株菌絲,用Fungal DNA Midi Kit真菌DNA提取試劑盒(OMEGA)提取病原菌Y2的DNA作為模板,采用真菌通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′) 和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)對提取的DNA進行PCR擴增,擴增產物經1.2%瓊脂糖凝膠電泳檢測條帶后,切膠回收PCR目的條帶送至重慶擎科興業生物技術有限公司進行測序。將病原菌Y2的18S RNA序列與NCBI數據庫相近序列進行比對。利用MEGA7.0的Clustal W進行多序列比對,再采用鄰接法(neighbor joining,NJ)進行菌株系統發育樹的構建。
1.2.3 藍莓葉斑病病原菌生物學特性研究
無菌環境下,截取直徑5 mm的病原菌新鮮菌餅,置于7個不同的碳源(葡萄糖、D-果糖、乳糖、D-麥芽糖、可溶性淀粉、甘露醇、蔗糖)培養基,不同的氮源(硫酸銨、尿素、甘氨酸、蛋白胨、酵母粉、牛肉膏、硝酸鈉)培養基,6個不同營養基質(PDA、燕麥、PSA、藍莓煎葉、WA、C)中培養,第3天量取菌落直徑,第5天時對培養皿拍照,30 d后測定產孢量;接種在pH值初始值分別為 5、7、9、11的PDA平板中央,第3天量取菌落直徑,第5天時對培養皿拍照,30 d后測定產孢量;置于不同溫度(5、15、25、28、35 ℃)下培養3 d,測量菌落直徑,在第5天時對培養皿拍照,30 d 后測定產孢量;置于3個光照處理(全黑暗:24 h 黑暗;全光照:24 h光照;半光照:光照12 h/黑暗 12 h)下培養,第3天量取菌落直徑,第5天時對培養皿拍照,30 d 后測定產孢量。以上所有處理均設置3個重復,菌落直徑測量采用十字交叉法,產孢量用血球計數板測定。
1.3 數據分析
利用Excel 2010、SPSS 19進行數據處理和Photoshop作圖。
2 結果與分析
2.1 藍莓葉斑病病原菌的分離及鑒定
分離得到編號為Y2的病原菌。在28 ℃恒溫環境下,病原菌Y2在PDA培養基上生長 6 d 時,病原菌的生長速度較慢,產孢,氣生菌絲較發達,菌絲呈白色、密集,基質背面呈淡黃色(圖1)。分生孢子單生,褐色,卵、圓形倒棒形,有縱橫隔膜(圖2)。
結合病原菌Y2的菌落形態和顯微特征,初步將病原菌Y2判定為鏈格孢屬(Alternaria)菌物。對病原菌Y2進行rDNA-ITS測序,將獲得的序列在NCBI數據庫進行BLAST同源性比對,比對結果表明,病原菌Y2序列與登錄號為MN756011.1的Alternaria alternate同源性最高,根據MEGA7.0中鄰接法構建病原菌Y2的系統發育樹(圖3)。系統發育樹結果表明,病原菌Y2與鏈格孢菌(Alternaria alternate) MN756011.1菌株的親緣關系最近,結合病原菌的形態特征和ITS序列特征,故最終將病原菌Y2鑒定為鏈格孢菌(Alternaria alternate)。
2.2 藍莓葉斑病病原菌Y致病性的測定
將病原菌Y2的菌餅接種到針刺傷的藍莓葉片傷口處,以無菌PDA培養基的菌餅作為對照,培養 3 d 時取下菌塊,5 d后觀察到:接種病原菌Y2的葉片在接種處有少量的氣生菌絲,葉片傷口處呈褐色且呈較規則的圓形,菌絲呈白色(圖4-A),與采集的田間病葉發病癥狀相似(圖4-C),未接種的葉片未出現發病癥狀(圖4-B),根據科赫氏法則,從發病的葉片病健交界部位組織分離得到一種分離物,和Y2為同一菌株。
[FK(W10][TPLQX44.tif][FK)]
2.3 不同碳源對菌絲生長的影響
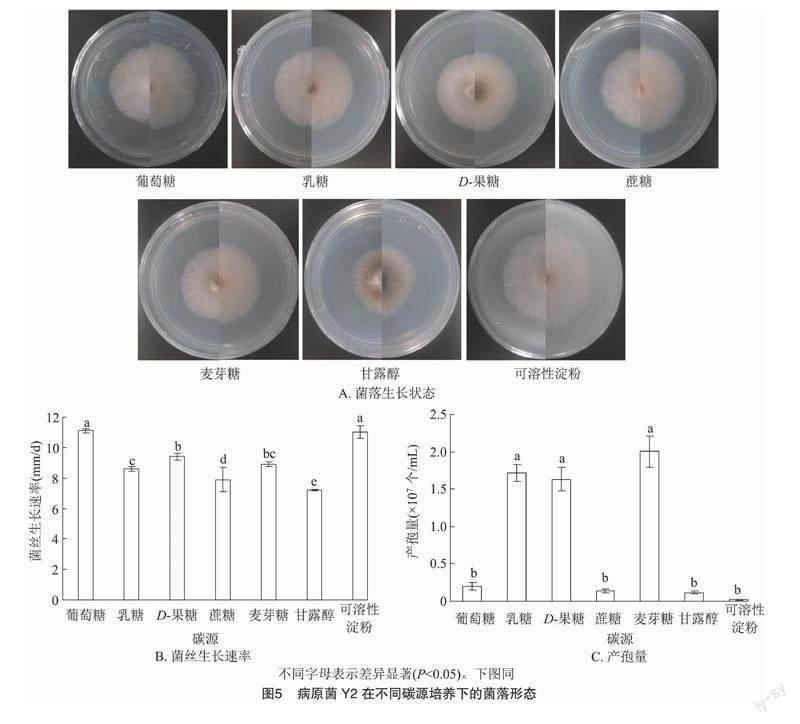
病原菌Y2在7種不同碳源培養基中的菌落形態都呈較規則的圓形,菌絲密集,有少量氣生菌絲;在甘露醇作碳源時產生較多的黃色色素,其次,D-果糖、麥芽糖2種碳源培養基培養時有少量黃色色素,其余不同碳源培養基均有微量黃色色素產生(圖5-A)。病原菌Y2在7種不同碳源培養基中菌絲生長速率存在顯著差異(P<0.05,下同),在葡萄糖、可溶性淀粉作為碳源時的菌落狀態最好,菌絲的繁殖速度最快,生長速率為11.1、11.0 mm/d;顯著高于其他碳源的生長速率;生長速率最慢的是碳源為甘露醇,為7.2 mm/d(圖5-B)。30 d后,測定其孢子濃度(圖5-C)。病原菌Y2在麥芽糖、乳糖和D-果糖為碳源時產孢量顯著高于其他碳源,麥芽糖作為碳源的培養皿中產孢子數量最多,為 2.003×107個/mL,以可溶性淀粉、葡萄糖、蔗糖和甘露醇為碳源時病原菌Y2的產孢量無顯著差異,以可溶性淀粉為碳源時最少,為0.022×107個/mL,以葡萄糖、蔗糖和甘露醇為碳源時分別為0.202×107、0.138×107、0.119×107個/mL。
2.4 不同氮源對菌絲生長的影響
病原菌Y2在7種不同氮源培養基中菌落形態較規則,均為圓形,氣生菌絲較密集,在以蛋白胨、甘氨酸、硫酸銨、尿素為培養條件下,菌落圓心部位菌絲高于周圍菌絲,形成圓形凸起,其中氮源為蛋白胨時凸起較明顯;菌落在以酵母粉、甘氨酸、尿素、牛肉膏為氮源培養時均產生黃色色素,其余條件下無色素產生,以酵母粉培養時產生較大量的黃色色素與褐色色素(圖6-A)。不同氮源處理下菌絲生長速率存在顯著差異,在以蛋白胨為氮源時菌絲生長速率顯著高于其他氮源,其菌落狀況最好,繁殖最快,生長速率為13.3 mm/d,生長速率最小的是以尿素為氮源時,其生長速率為7.7 mm/d,以硝酸鈉、甘氨酸為氮源時菌絲生長速率無顯著差異,均為 10.5 mm/d (圖6-B)。不同氮源處理下病原菌Y2產孢量有顯著差異(圖6-C),在以硫酸銨作為氮源時的病原菌Y2所產孢子量顯著高于其他氮源,為21.667×107個/mL,說明硫酸銨能誘導病原菌Y2萌發孢子,是病原菌Y2產孢的最佳的氮源。
2.5 不同pH值對菌絲生長的影響
病原菌Y2在pH 值為 5、7、9、11的環境下都能形成較規則的圓形、菌絲較為密集的菌落,但以在pH值=7環境中的菌落直徑最大,生長狀態最佳,當pH值從5到11變化時,菌絲逐漸密集,且都有黃色素產生(圖7-A)。病原菌Y2在pH值=7時生長速率顯著高于其他pH值梯度,為10.5 mm/d;生長速率最小的是在pH值=11時,為8.7 mm/d;在pH值=5、pH值=9時,其菌絲生長速率分別為9.9、9.8 mm/d,二者間無顯著差異(圖7-B),病原菌Y2在酸性條件下更適宜生長。過酸、過堿的環境均不利于病原菌Y2產孢, 而pH值=7時酸堿適中,可以促進Y2產孢,產孢量顯著高于其他pH值,為2.573×107個/mL,pH值=11時產孢量最少,只有0.074×107個/mL(圖7-C)。
2.6 不同溫度對菌絲生長的影響
病原菌Y2在5個溫度值下培養5 d后均有菌絲長出,且菌落均呈規則圓形,氣生菌絲密集,均有黃色素產生(圖8-A)。當溫度降至 5 ℃,菌株幾乎停止生長,在25 ℃下最適宜病原菌Y2生長,生長速率顯著高于其他溫度,為8.9 mm/d;在5 ℃下最不適宜生長,生長速率為2.0 mm/d;在15、28、35 ℃ 下菌落生長速率分別為5.4、7.4、3.4 mm/d (圖8-B)。低溫可以促進病原菌Y2產孢,5 ℃時產孢量顯著高于其他溫度,為4.067×107個/mL,在溫度為15 ℃和35 ℃時產孢量無顯著差異,產孢量很少或幾乎不產孢,當溫度為25 ℃和28 ℃時產孢量較多,分別為2.077×107個/mL和0.435×107個/mL(圖8-C)。
2.7 不同光照對菌絲生長的影響
在全光照、全黑暗和光暗交替3種光照條件下,病原菌Y2都能正常生長,菌落均呈規則圓形,且均有黃色色素產生,全黑暗培養下產生較大量的黃色、褐色色素;菌落氣生菌絲均致密(圖9-A)。病原菌Y2在全光照條件下生長速率顯著高于全黑暗和光暗交替條件下,為12.4 mm/d;生長速率最小是在光暗交替條件下,為10.2 mm/d;在全黑暗條件下菌絲生長速率為10.9 mm/d (圖9-B)。計算不同光照處理下致病菌Y2所產孢子量,不同光照對病原菌Y2產孢有顯著影響。全黑暗處理的病原菌Y2所產孢子數量是3個處理組中最高的,為1.62×107個/mL,光暗交替處理下的Y2孢子數目最少,為0.012×107個/mL(圖9-C)。
2.8 不同培養基對菌絲生長的影響
病原菌Y2在6種不同培養基中,28 ℃下培養5 d 后,觀察到病原菌Y2在6種不同培養基中均呈較規則圓形,氣生菌絲在PDA、PSA、察氏(C)培養條件下致密,在WA、藍莓煎汁培養基、燕麥瓊脂培養基中菌絲較稀薄;菌落在PDA、PSA、WA和燕麥瓊脂培養基中有黑色色素產生,在察氏培養基中有黃色色素產生,藍莓煎汁瓊脂培養基中無色素產生(圖10-A)。病原菌Y2在察氏培養基中生長速率顯著高于其他培養基,為17.0 mm/d;生長速率最小的是在WA培養條件下,生長速率為7.0 mm/d;在PDA、PSA、藍莓煎汁瓊脂和燕麥瓊脂培養條件下菌絲生長速率分別為10.6、11.1、8.9、11.9 mm/d (圖10-B)。統計病原菌Y2在不同培養基中所產孢子量,發現WA和燕麥培養基中的孢子數顯著高于PDA、PSA、察氏和藍莓煎汁培養基,其中WA中孢子數最多, 為4.495×107個/mL,在察氏中Y2孢子數目最少,僅僅為0.014×107個/mL,在燕麥中產孢量較多,為4.233×107個/mL,在其他基質下孢子數量差異不顯著(圖10-C)。
3 討論與結論
本研究從貴州省人工栽培藍莓園內發現典型藍莓葉斑病植株,通過組織分離法進行病原分離,科赫法則證明其致病性,根據菌落形態特征及現代分子生物技術,最終鑒定藍莓葉斑病病原菌Y2為鏈格孢菌,病原菌Y2侵染離體葉片形成的病斑為紅褐色圓形病斑,與鏈格孢菌侵染藍莓葉片形成的病斑形似;鏈格孢菌作為主要的藍莓葉斑病病原菌,在藍莓葉部病害中較為常見。鏈格孢菌還可引起多種植物病害,如煙草赤星病[16]、高粱葉斑病[17]、梨黑斑病[18]、花椒穗枯病[19]、番茄黑斑病[20]、棗果黑斑病[21]、蕎麥葉枯病[22]和玉米葉斑病[23]等。
本研究發現藍莓葉斑病病原菌-鏈格孢菌Y2,菌絲生長的最適pH值為7,與周銀麗等[24]和盧文潔等[22]研究結果一致;黎研研等[25]、周銀麗等[24]、盧文潔等[22]和劉曉琳等[21]的研究結果均表明菌絲生長的最適溫度為25 ℃,本研究得到最適氮源為蛋白胨與黎研研等[25]、王宏等[18]和劉曉琳等[21]的研究結論相同,盧文潔等[22]的研究表明,全光照條件下菌絲生長最快,本研究也得到了相同的結論。關于鏈格孢菌的生物學特性有很多不同的報道。喬鏡澄等研究了番茄黑斑病樣品中鏈格孢菌的生物學特性,結果顯示,該菌株的最適溫度為28 ℃,最適pH值為6,最佳碳源為乳糖,最佳氮源為酵母膏,PDA為其最適宜生長的培養基,菌絲在完全黑暗條件下擴展最快[20];楊曉平等報道了湖北梨黑斑病病原菌鏈格孢菌菌絲生長的最佳條件是PSA培養基[26];陳鳳美等報道了江蘇銀杏內生鏈格孢菌最適pH值為5~9,最適氮源為硝酸鈉[27];鄭肖蘭等報道了海南省南繁區玉米鏈格孢葉斑病鏈格孢菌的最適溫度為30 ℃,最適pH值為5,最適培養基是PDA+玉米綠葉煎汁,最適的氮源是纈氨酸和甘氨酸。
本試驗研究了不同處理下病原菌Y2的產孢情況,結果表明,病原菌Y2產孢的最適碳源是麥芽糖、最適氮源是硫酸銨,最適溫度為5 ℃、pH值為7,以WA為營養基質有利于病原菌Y2產孢,全黑暗促進Y2產孢。胡中會等報道了柑橘褐斑病病原菌為鏈格孢菌,其產孢的最適溫度為21 ℃,最適碳源為甘露醇,最適氮源為甘氨酸[28];鄒鳳蓮等報道了番紅花鏈格孢菌的最適溫度為28 ℃,最適碳源為蔗糖,最適氮源為牛肉浸膏[29];袁高慶等報道了毛葉棗黑斑病病原菌的最適溫度為30 ℃,最適pH值為5,PDA培養基最適宜病菌產孢[30]。說明適宜鏈格孢菌產孢的條件隨著地理環境、寄主的變化而產生較大差異。本研究對該菌菌絲生長和產孢的生物學特性的研究結果可為后續該菌的致病機制研究以及科學有效的防治提供重要參考。
本研究明確了貴州省藍莓葉斑病的病原菌為鏈格孢菌,并分析了其生物學特性。中國藍莓葉斑病的病原菌種類復雜多樣,病害防控工作的基礎原則是準確鑒別病原菌;同一病原菌的生物學特性在不同環境、不同寄主中存在差異,因此,研究病原菌Y2的生物學特性對進一步闡述葉斑病病害的發病規律和綜合防治具有的重要指導意義。
參考文獻:
[1]胡雅馨,李 京,惠伯棣. 藍莓果實中主要營養及花青素成分的研究[J]. 食品科學,2006,27(10):600-603.
[2]陳介甫,李亞東,徐 哲. 藍莓的主要化學成分及生物活性[J]. 藥學學報,2010,45(4):422-429.
[3]李亞東,張志東,吳 林. 藍莓果實的成分及保健機能[J]. 中國食物與營養,2002,8(1):27-28.
[4]余 磊,趙建榮,徐勝光,等. 云南藍莓葉斑病的病原菌鑒定[J]. 植物病理學報,2015,45(2):216-219.
[5]馮 璐,欒雨時,范永強,等. 越橘葉斑病病原菌的鑒定[J]. 東北農業大學學報,2007,38(5):614-618.
[6]余 磊,趙建榮,Impaprasert R,等. 藍莓枝枯病病原菌鑒定[J]. 植物病理學報,2013,43(4):421-425.
[7]傅俊范,彭 超,嚴雪瑞,等. 藍莓根癌病發生調查及病原鑒定[J]. 吉林農業大學學報,2011,33(3):283-286,292.
[8]嚴雪瑞,趙睿杰,周 源,等. 藍莓灰霉病菌差異性比較及藍莓品種抗病性鑒定[J]. 果樹學報,2014,31(5):912-916.
[9]嚴雪瑞,王 旭,胡夢瓊,等. 藍莓間座殼芽枯病病原菌鑒定及其生物學特性[J]. 植物病理學報,2015,45(5):556-560.
[10]徐成楠,周宗山,張紅軍,等. 藍莓枝干潰瘍病病原鑒定[J]. 植物病理學報,2012,42(5):532-535.
[11]You M P,Lanoiselet V,Wang C P,et al. First report of Alternaria leaf spot caused by Alternaria tenuissima on blueberry (Vaccinium corymbosum) in western Australia[J]. Plant Disease,2014,98(3):423.
[12]Norman D J,Bocsanczy A M,Harmon P,et al. First report of bacterial wilt disease caused by Ralstonia solanacearum on blueberries (Vaccinium corymbosum) in Florida[J]. Plant Disease,2018,102(2):438.
[13]董克鋒,姜惠鐵,高海霞. 冬暖棚有機藍莓園藍莓灰霉病發生及預防措施[J]. 中國果樹,2012(5):67-69,79.
[14]嚴雪瑞,周 源,趙睿杰,等. 藍莓葉穿孔病菌鑒定及其生物學特性研究[J]. 北方園藝,2014(16):123-127.
[15]張國輝,劉德波,宋盛英,等. 黔東南州藍莓葉部病害的種類調查和病原鑒定[J]. 中國森林病蟲,2017,36(2):42-46.
[16]彭希文,劉光珍,楊永柱,等. 云南省煙草赤星病(Tobacco brown spot)病原研究及其防治藥劑的篩選[J]. 西南農業大學學報,2000,22(2):153-156.
[17]趙艷琴,于華榮,石 凱,等. 高粱鏈格孢葉斑病病原鑒定[J]. 植物病理學報,2017,47(2):282-285.
[18]王 宏,常有宏,陳志誼. 梨黑斑病病原菌生物學特性研究[J]. 果樹學報,2006,23(2):247-251.
[19]劉 峰,楊文香,張 娜,等. 花椒枯穗病病原鑒定[J]. 植物病理學報,2013,43(3):310-313.
[20]喬鏡澄,劉 宇,馬敬昊,等. 番茄黑斑病病原菌的鑒定及生物學特性研究[J]. 江蘇農業科學,2017,45(10):94-97.
[21]劉曉琳,劉 玉,馬 榮,等. 新疆棗果黑斑病病原菌鑒定及生物學特性[J]. 西北林學院學報,2015,30(3):132-138.
[22]盧文潔,羅志明,李春花,等. 云南省蕎麥葉枯病病原菌鑒定及其生物學特性[J]. 植物保護學報,2019,46(5):1065-1072.
[23]鄭肖蘭,趙 爽,韓小雯,等. 海南省南繁區玉米鏈格孢葉斑病病原菌鑒定及其生物學特性[J]. 江蘇農業科學,2018,46(6):82-87.
[24]周銀麗,白建波,胡先奇,等. 芒果葉斑病病原菌的鑒定及其生物學特性[J]. 江蘇農業科學,2013,41(5):108-109.
[25]黎妍妍,楊 濤,賈欣欣,等. 湖北省煙草赤星病菌生物學特性研究[J]. 湖北農業科學,2018,57(22):27-31.
[26]楊曉平,胡紅菊,王友平,等. 梨黑斑病病原菌的生物學特性及其致病性觀察[J]. 華中農業大學學報,2009,28(6):680-684.
[27]陳鳳美,劉 群,蔣繼宏,等. 銀杏內生鏈格孢菌GI009生物學特性研究[J]. 西北林學院學報,2005,20(4):112-114,121.
[28]胡中會,趙立華,張 鐵. 柑橘褐斑病菌菌絲生長和孢子萌發研究[J]. 云南農業大學學報(自然科學),2015,30(1):9-12.
[29]鄒鳳蓮,汪志平,盧 鋼. 番紅花鏈格孢菌的分離及其生物學特性研究[J]. 浙江大學學報(農業與生命科學版),2006,32(2):162-167.
[30]袁高慶,賴傳雅,黃麗華. 毛葉棗黑斑病的病原鑒定及其生物學特性研究[J]. 中國農學通報,2003,19(1):44-47.
收稿日期:2022-04-12
基金項目:貴州省科技計劃項目(編號:黔科合支撐[2017]2567、黔科合平臺人才[2018]5781);貴州大學SRT計劃[編號:貴大SRT(2019)252號]。
作者簡介:羅其鑫(1999—),女,貴州貴陽人,碩士研究生,研究方向為森林病理學。E-mail:452450608@qq.com。
通信作者:侯 瑞,博士,副教授,主要從事森林病理學等研究工作。E-mail:jiayouhourui123@163.com。
