溫度脅迫對農作物和入侵雜草的生長影響機制
2023-07-10 07:59:14梅英皓孔方麗張軼釩祁珊珊戴志聰杜道林
雜草學報 2023年1期
梅英皓 孔方麗 張軼釩 祁珊珊 戴志聰 杜道林


摘要:在全球氣候變化的背景下,我國極端氣候出現的次數比以往更加頻繁,而極端的高溫或低溫氣候都對農作物的生長發育有負面影響,嚴重影響了我國農作物的產量。溫度的改變加快了入侵雜草的遷移和演替,農田系統中的入侵雜草和農作物競爭資源并在競爭中獲得優勢,最終對農業生產造成極大的威脅和危害。本文綜述了溫度脅迫對農作物和入侵雜草生長的影響,并闡述了在溫度改變背景下農作物系統可能面臨的困境,最后展望了未來在溫度變化背景下的農作物-雜草系統相關研究的方向。
關鍵詞:農作物;入侵雜草;極端氣候;溫度脅迫;農作物-雜草系統
中圖分類號:S451;Q945.78文獻標志碼:A文章編號:1003-935X(2023)01-0001-10
Effect Mechanistic of Temperature Stress on Growth of Crops and Invasive Weeds
MEI Ying-hao1, KONG Fang-li1, ZHANG Yi-fan1, QI Shan-shan2, DAI Zhi-cong1, DU Dao-lin1
(1.School of Environment and Safety Engineering,Jiangsu University,Zhenjiang 212013,China;
2.School of Agricultural Engineering,Jiangsu University,Zhenjiang 212013,China)
Abstract:In the context of global climate change,the frequency of extreme climate in China is more frequent than before,and the extreme high temperature or low temperature climate negatively impacts on the growth and development of crops,and seriously affectes their yield. The change in climate temperature accelerates the migration and succession of invasive weeds which compete with crops for resources and usually gain advantages in farming systems .It ultimately poses a great threat and harm to agricultural production. This paper reviewed the effects of temperature stress on the growth of crops and invasive weeds,and described the possible dilemmas faced by crop systems,and finally looked into future directions for research related to crop-weed systems under the context of climate temperature change.
Key words:crop;invasive weed;extreme climate;temperature stress;crop-weed system
收稿日期:2022-09-09
基金項目:國家自然科學基金(編號:32271587、32071521);江蘇省自然科學基金(編號:BK20211321);江蘇省碳達峰碳中和科技創新專項資金(編號:BK20220030);江蘇大學青年英才計劃。
作者簡介:梅英皓(1997—),男,江蘇南京人,碩士研究生,主要從事農業環境生態學研究。E-mail:myh809772414@163.com。
通信作者:戴志聰,博士,副教授,主要從事環境科學與工程、環境生態學研究。E-mail:daizhicong@163.com。
外來入侵雜草是指由于人為或自然原因,從原始生境進入一個新生境后對該環境中的生物、農林牧漁業生產造成巨大經濟損失、對人類健康造成危害,破壞新生境中原來生態平衡的一類植物[1]。介于全球氣候變化的步伐不斷加快,外來雜草入侵的形勢已經迫在眉睫,如今外來雜草入侵是全球性的環境問題,尤其對農業生產造成了極大的危害。為了進一步了解有關農作物-雜草系統相關研究的現狀,筆者通過WOS數據庫檢索了近35年以來相關方向的出版物(圖1),發現近35年以來脅迫環境下農作物-雜草系統受到的關注度在不斷上升,特別是近5年的相關研究成果顯著。
溫度脅迫被認為是影響生物過程、限制植物生長和生產力的主要環境因素之一[2]。研究表明,全球平均氣溫將會在2050年前上升 1.5 ℃[3]。因此,短期內平均溫度的升高和極端溫度脅迫的出現都可能對農作物生長和發育產生不可挽回的負面影響[2]。例如,溫度脅迫能夠誘導植物在分子、細胞、生理和生化水平上發生顯著改變[4],并會嚴重破壞蛋白質、破壞細胞膜、影響水養關系、干擾光合作用、影響植物的生長發育,最終導致產量下降[2]。對農田系統而言,雜草入侵(生物脅迫)和溫度脅迫(非生物脅迫)的共同作用可能對田間農作物產生的負面影響更加嚴重,例如雜草競爭和高溫脅迫的共同作用對大豆的生產力造成了明顯的抑制[5];高溫加快了雜草的入侵速度并導致了大豆和玉米產量的損失[6]。
當今,入侵雜草在世界各地內都造成了嚴重的危害,例如嚴重破壞了當地的物種多樣性,對生態環境造成危害;過度入侵傷害了原生境內的本地農作物生長,給農業生產和人們的生活帶來巨大麻煩;需要消耗大量人力物力對入侵雜草進行治理,造成了巨大的經濟損失[7]。因此,揭示溫度脅迫下農作物及雜草的響應機制,合理預測并預防入侵雜草對農作物產生的負面影響,提出對農作物管理的可行性計劃,有助于實現未來農業的可持續發展并保障日益增長的世界人口的糧食安全[8]。本文分別闡述了溫度脅迫對農作物及入侵雜草生長的影響,并進一步探討現階段環境溫度脅迫對農作物-入侵雜草系統的影響,最后對未來有關溫度脅迫下農作物培養的研究方向進行展望。
1 溫度脅迫對農作物生長的影響
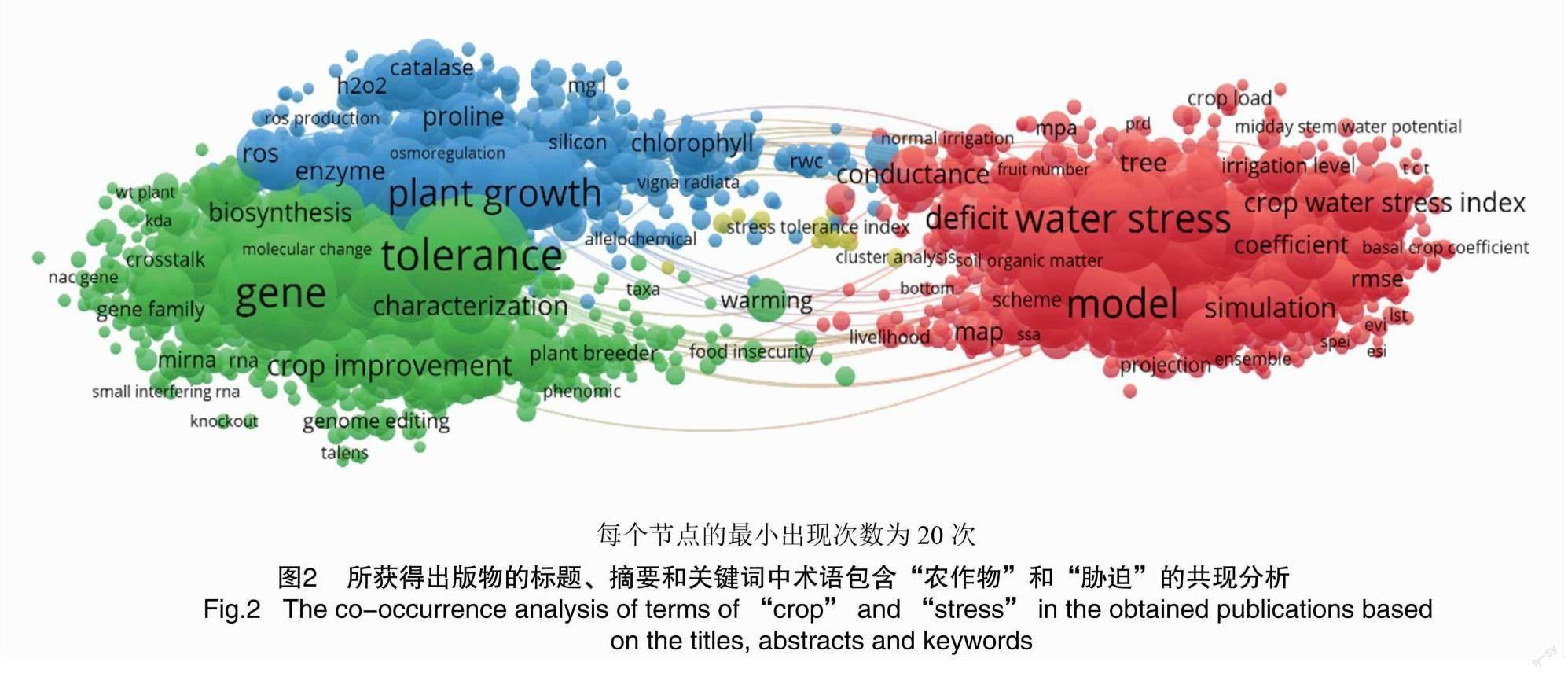
非生物脅迫通常是相互關聯的,無論是單獨的還是聯合的,都會引起形態、生理、生化和分子變化,從而對植物的生長和生產力產生不利影響,并最終影響農作物的產量。高溫、干旱、低溫和鹽分是主要的非生物脅迫,它們會在包括農作物在內的植物物種中引起嚴重的細胞損傷。植物生長和繁殖過程中會自然發生溫度波動,然而極端高溫或低溫變化會破壞正常生長所需的分子間的相互作用,從而損害植物發育和坐果。為了進一步了解溫度脅迫對農作物影響的相關研究程度,筆者首先在WOS數據庫中“農作物”和“脅迫”的基本搜索選項卡下搜索了所有領域的結果,利用VOSviewer 1.6.18軟件對WOS數據庫的前10 000個結果進行分析并生成了網絡圖,添加條件中每個術語出現的最小次數為20次。圖2中節點的距離反映了各個熱詞間的關聯性。網絡圖的結果表明,與脅迫(tolerance)、植物生長(plant growth)以及模型(model)有關的研究占據較大的比重,同時脅迫條件也與植物生長關聯性極強。極端氣候溫度的變化已經對全世界的農業生產產生了重大影響,因為溫度脅迫會造成嚴重的產量損失,給未來的全球糧食安全帶來巨大風險[9]。據預測,極端氣候溫度將對植物的生長和發育產生普遍的負面影響,導致作物生產力的災難性損失,并導致廣泛的饑荒。總的來說,溫度脅迫給未來的農業生產和全球糧食安全帶來了額外的挑戰,因此對溫度脅迫下農作物生長機制的研究是極具意義的。
1.1 高溫脅迫對農作物生長的影響
植物在自然界所面臨的主要非生物脅迫形式中,高溫脅迫對植物細胞的生理和代謝有不同程度的負面影響。植物對高溫的敏感性隨植物的發育階段而變化,高溫脅迫在一定程度上影響所有的無性和生殖階段。在高溫下可以觀察到植物的各種生理損傷,例如葉片和莖干被灼燒、葉片的脫落和衰老、芽和根的生長被抑制或是果實被損傷,這些都能夠導致植物的生產力下降[10]。但在許多情況下,植物會通過改變自身結構形態來避免高溫脅迫帶來的影響,例如下胚軸和葉柄伸長[11]。
通常來說,高溫脅迫會引起植物器官呼吸和光合作用的變化,從而導致植物生命周期的縮短和生產力的下降[12]。早期的高溫脅迫會促使植物的葉綠體蛋白復合物發生結構改變和酶活性降低[13]。此外,高溫脅迫會通過對細胞膜、微管組織并最終對細胞骨架造成傷害,從而改變細胞膜的通透性并改變細胞的分化、伸長和擴張過程[14]。例如,葉綠體基質和類囊體膜系統被認為是受高溫損傷的主要部位[15]。同時,高溫對葉綠素和光合器官產生的負面影響與有害活性氧 (ROS) 的產生有關[16],高溫脅迫通過增加葉綠素酶活性和減少光合色素的數量來降低植物的光合和呼吸活性[17]。
一般而言,在高溫脅迫下的植物組織中,代謝產物的生物合成往往會受到干擾[18]。高溫通過下調碳水化合物代謝中的特定基因來改變淀粉積累、碳代謝酶和蔗糖合成酶的活性[19],響應高溫脅迫而積累的主要代謝物包括脯氨酸、甘氨酸甜菜堿和可溶性糖[20]。許多植物物種也會積累其他滲透物,例如糖醇(多元醇)、三級和四級銨化合物[21]。酚類等次級代謝物包括黃酮類化合物、花青素和植物類固醇,也積極參與植物在高溫脅迫下的響應,并且通常在與耐熱性相關的非生物脅迫反應中發揮作用[20]。例如,高溫脅迫會導致番茄植株的可溶性酚類物質積累,提高苯丙氨酸解氨酶的活性并降低過氧化物酶和多酚氧化酶的活性,這可能是對高溫脅迫的一種響應[22]。對植物激素而言,包括脫落酸(ABA)、水楊酸(SA)和乙烯(ET)在內的幾種關鍵植物激素在高溫脅迫下含量會增加;而其他一些激素,例如細胞分裂素(CK)、生長素(AUX) 和赤霉素(GA)的含量則會降低,這些變化最終會導致植物過早衰老[23]。
對農作物而言,農作物的發育和生理過程廣泛受到高溫脅迫的負面影響。當高溫發生在繁殖等關鍵發育階段時,這成為農作物適應環境變化的主要限制之一[24]。例如,小麥生殖發育過程中的高溫加速了光合作用和葉面積的下降,降低了其芽和籽粒的質量及糖含量,同時也降低了水分利用效率[25]。長期以來,人們一直認為有性生殖和開花對高溫脅迫極為敏感,這通常會導致農作物產量下降[26]。在溫室下進行的研究表明,高溫在花芽萌生階段對其影響最大,植物對高溫的敏感性可以維持數十天[27]。許多豆類和谷類在開花期間對高溫脅迫表現得很敏感,并且一些溫帶甚至熱帶果樹物種的坐果率也嚴重下降[28]。此外,在許多對溫度敏感的農作物中可以廣泛觀察到高溫脅迫導致的雄性不育,高溫脅迫對花粉發育造成的損害是農作物減產的主要因素之一[29-30]。例如,高溫脅迫導致了番茄在發育階段可用碳水化合物的消耗,從而導致其坐果和其他相關參數減少[31]。在高粱中,高溫脅迫也減少了其花粉粒中碳水化合物及柱頭組織中三磷酸腺苷(ATP)的積累,從而影響了其最終的產率[32]。總的來說,高溫脅迫會對農作物的各種生理過程產生負面影響,最終影響農作物的營養和生長發育,從而對坐果和產量產生負面影響。
1.2 低溫脅迫對農作物生長的影響
低溫脅迫是嚴重影響植物生長發育、影響作物生產力的主要環境因素之一。植物已經進化出一系列機制,使它們能夠在生理和分子水平上適應冷脅迫。過去的20年中,在確定與低溫脅迫耐受性有關的關鍵成分并剖析其調節機制方面取得了很大進展。然而,許多農作物,包括水稻(Oryza sativa)、玉米(Zea mays)、番茄(Solanum lycopersicum)、大豆(Glycine max)和棉花(Gossypium hirsutum)缺乏適應寒冷溫度的能力[33]。低溫脅迫往往對農作物的生長和發育產生不利影響,限制了農作物物種的地理分布,并降低了世界范圍內的作物產量[34]。研究表明,植物已經進化出復雜的機制來抵御寒冷的壓力,冷馴化是其中之一,這是植物在先前暴露于非致命低溫時獲得增加的抗凍性的過程[35-36]。
冷脅迫,包括低溫(0~15 ℃)和冷凍(低于 0 ℃),是一種非生物脅迫,對植物的生長和農業生產力產生不利影響[37-38]。低溫脅迫通常會限制植物的生長發育,并對植物細胞有幾個主要影響。首先,低溫脅迫會影響植物細胞中的膜硬化,這被認為是觸發植物下游冷脅迫反應的主要事件[39]。其次,低溫脅迫會擾亂蛋白質或蛋白質復合物的穩定性,并降低酶如ROS清除酶的活性,由于二氧化碳的光合作用固定在低溫脅迫下十分有限,這些過程都會導致光抑制和光合作用受損,以及相當大的膜損傷[40-41]。因此,對低溫脅迫引起的光抑制的耐受性似乎也是一種冷適應的機制,并且這種機制與農作物的耐冷性密切相關。例如,菠菜、冬小麥、黑麥和蠶豆等耐寒性農作物能在低溫環境中保持較高的二氧化碳同化率,而對低溫敏感的黃瓜、煙草和大米的二氧化碳同化率則會顯著降低[42]。再次,低溫脅迫會影響基因表達和蛋白質合成,因為它有利于RNA二級結構的形成[43]。冷凍脅迫比寒冷脅迫對植物的危害更大,甚至可能導致植物死亡。在自然條件下,凍害始于細胞外冰核,一旦冰核形成,就會生長并形成冰晶,冰晶會擴散到質外體中,在那里它們會誘導水流出,導致細胞脫水,當冰晶擴散到細胞中時,會發生不可逆轉的損害[34]。植物已經進化出復雜的機制來限制寒冷引起的損害,例如,冷馴化是一個過程,在該過程中,植物在非致命低溫下暴露幾天會增強抵抗隨后的冰凍脅迫的能力[35-36]。植物會通過合成大量的保護性物質(例如可溶性糖、脯氨酸)和蛋白質(例如LEA、AFP、CSP)來提高它們對冷脅迫的耐受性[44]。可溶性糖、脯氨酸和其他低分子量溶質可作為滲透物保護植物免受冷脅迫造成的損害[40]。最后,低溫脅迫會通過影響植物激素的變化從而影響其耐寒性,植物激素中的脫落酸(ABA)、生長素、赤霉素(GA)、水楊酸(SA)和乙烯與寒冷反應呈正相關或負相關[45]。通常ABA水平會隨著溫度的降低而增加[46],但ABA對冷響應轉錄調節的作用較為有限。低溫對一些植物的結構有很大的影響,例如生長素和GA與低溫條件下的細胞伸長率有直接相關關系[47]。在低溫條件下,內源性GA和生長素水平降低,會導致植物的矮化結構,例如小麥品種Rht3的表型矮化[48];內源性游離SA和葡糖基SA在農作物的降溫過程中積累[22];SA處理增強了各種農作物的耐寒性,例如水稻、玉米、小麥、馬鈴薯等[49-50]。綜上所述,低溫脅迫會嚴重影響農作物的正常生長發育和生產力,因此有關低溫脅迫環境下如何幫助農作物完成生長發育的更深層次的研究是不可或缺的。
2 溫度脅迫對入侵雜草生長的影響
與農作物相比,入侵雜草具有更強的可塑性,因為它們沒有被針對特定的優勢性狀(如抗病性、生長均勻、高結實率等)進行選擇培育。因此,與農作物相比,入侵雜草往往在響應環境壓力時能夠表現出更強的適應能力。此外,入侵雜草之間的高度遺傳多樣性使它們能夠在氣候變化的環境下獲得更強的競爭適應性[51],氣候變化對入侵雜草的影響主要包括物種豐度、豐富度、地理范圍和物候。同樣地,為了進一步了解溫度脅迫對雜草影響的相關研究程度,筆者首先在WOS數據庫中“雜草”和“脅迫”的基本搜索選項卡下搜索了所有領域的結果,再利用VOSviewer (1.6.18)軟件對WOS數據庫中共2 306個結果進行分析并生成了網絡圖,添加條件中每個術語出現的最小次數為20次,圖中節點的距離反映了各個熱詞間的關聯性。結果如圖3表示,與管理(management)、競爭(competition)以及機制(mechanism)有關的研究占據較大的比重,同時競爭也與管理關聯性極強,這說明脅迫環境下有關雜草機制的研究是當前的熱點和重心。
環境溫度是植物生長最重要的環境因子之一,溫度的變化會在短期或長期時間尺度內對入侵雜草產生影響。許多研究報道了入侵雜草對環境溫度變化的響應,例如,入侵雜草石茅(Sorghum halepense)在寒冷氣候下的空間分布由于其根莖不耐受低溫而被限制[52]。同樣地,牽牛花不耐低溫,但它們能夠在較高的溫度環境里發芽和生長[53]。此外,Ziska等的研究表明,在溫度升高 3 ℃ 的環境下,筒軸茅(Rottboellia cochinchinensis)的生物量和葉面積都在很大程度上得到增長[54]。許多入侵雜草的地理范圍主要由溫度決定,人們早就認識到溫度決定了入侵雜草在新環境中的成功定植[55],氣候變暖將影響入侵雜草的生長、繁殖和分布。例如,溫度升高改變了美國境內中西部和中南部地區的緯度區分,從而改變了對入侵雜草的地理限制;熱帶或亞熱帶環境中許多入侵性非常強的入侵雜草導致了大量大豆和玉米的損失[56]。此外,溫度上升可能對C3和C4植物的生長特別重要,可能更有利于C4入侵雜草[51],如鼠尾粟(Sporobolus indicus)。同時,這又可以為一些受限于低溫的入侵雜草提供合適的生長條件,而一些熱帶和亞熱帶物種的分布則會因此受到限制。一些熱帶和亞熱帶C4物種的分布可能向北轉移[57],從而使溫帶地區的農業受到入侵雜草的影響。此外,Ziska等認為,溫度上升將促進入侵雜草的擴張,在溫度升高3 ℃的情況下,筒軸茅的生物量和葉面積分別增加88%和68%[54],這將可能幫助其加快入侵進程。相反,極端低溫氣候的出現可能會對部分入侵雜草的入侵范圍造成限制,例如野生黍稷和加拿大薊皆受到低溫影響[57]。
總體而言,大量的研究表明了入侵雜草在氣候變暖的環境下有進一步擴張其入侵范圍的可能性,但有關低溫脅迫對入侵雜草的限制和影響仍需要大量的研究投入,這將為預測全球氣溫變化對入侵雜草的影響(特別是雜草往兩極入侵)提供可靠的依據和對策。
3 溫度脅迫對農作物-入侵雜草系統的影響
氣候變化對農作物與入侵雜草競爭結果的影響很難僅通過單一模型來預測和判斷,因為在農田系統中的各種氣候條件會同時發生交互作用,最終將影響農作物和雜草競爭的結果[6]。氣候溫度的上升往往伴隨著二氧化碳濃度的上升和干旱
頻率的增多[58],因此,僅單一地探討溫度脅迫對農作物和入侵雜草體系的影響是不夠充足的,需要將多種氣候變化因素的相互作用綜合考慮在內,這樣才能合理地預估溫度脅迫對農作物和入侵雜草競爭體系的影響。相似地,為了進一步了解溫度脅迫對農作物-雜草體系影響的相關研究程度,筆者首先在WOS數據庫中以“農作物”“雜草”和“脅迫”的基本搜索選項卡下搜索了所有領域的結果,再利用VOSviewer (1.6.18)軟件對WOS數據庫中共941個結果進行分析并生成了網絡圖,添加條件中每個術語出現的最小次數為20次。如圖4所示,與溫度(temperature)、管理(management)以及除草劑(herbicide)有關的研究占據較大的比重,同時溫度條件也與諸多術語有緊密的相關性。
有研究表明,相對較小的氣溫變化也會強烈刺激熱帶雜草的生長[59],但對于熱帶農作物而言,二氧化碳上升對這些雜草的潛在協同效應尚未明確。據報道,增高的溫度和二氧化碳皆對草原生態系統中的植物生長速度產生負面影響[60]。溫度和二氧化碳水平升高將改變雜草與農作物的競爭作用,因而對入侵雜草的控制也會受到這些環境變化的影響[61]。對入侵雜草而言,溫度脅迫會降低其蒸騰作用并導致其葉片表型特征的變化,或因二氧化碳升高而導致其根莖比的增大,這會影響雜草對除草劑的吸收,最終導致除草劑效率的降低[62]。對農作物而言,溫度脅迫會導致其葉片發育、開花、收獲和結果的物候發生改變[63]。此外,溫度脅迫會導致更高的呼吸速率、更短的種子形成期和更少的生物量產量,從而降低了農作物產量[64]。尤其是玉米、大豆、小麥和棉花等溫度敏感作物的產量[65]及杏仁等核果、葡萄等漿果、柑橘類等特殊作物的產量將隨著區域和局部尺度的溫度升高而降低[66]。
有關高溫脅迫和長期干旱對雜草入侵的影響預期表明,較長的干旱期與偶爾潮濕的氣候相交織將加劇雜草的入侵擴張。因為高溫脅迫和干旱都會對本地定植的農作物產生負面影響,削弱本地農作物的生存能力,從而留出更多的生態空間給予雜草入侵的可能[67]。總體來說,有關農作物和入侵雜草在溫度脅迫下的交互是復雜而重要的,這能夠為人們在未來農作物生產及入侵雜草防治上提供一定的思路和方法。
4 展望
在過去的幾年里,保障糧食安全和準備應對極端氣候變化的必要性變得越來越重要。許多研究表明,溫度的上升會加快入侵雜草進一步向北部低溫地區的擴張進程[68-70],并且入侵雜草較強的可塑性往往能夠幫助自身更快地適應溫度的改變[71]。因此,適應性更強的入侵雜草嚴重影響了本地農作物的生長和生存,給人類經濟和生態環境都造成了傷害。目前,在全球氣候變暖的背景下,大量的研究著重于環境脅迫對農作物類植物所產生的影響和危害。實際上,隨著全球氣候變化的加劇,全球范圍內入侵雜草的擴張也很大程度上影響了農作物的生長和生存空間。預計未來的氣候變化會以多種方式影響農業生產,氣候變化可能會影響農作物和入侵植物的生長,有時使農作物受益,有時使入侵雜草受益。由于極端高溫或低溫的出現頻率增高,許多地區的農作物產量將下降,而盡管技術進步,但入侵雜草的惡性競爭將進一步加劇農作物產量的下降。因此,未來需要一種方法,不僅能夠幫助農作物來抵消溫度脅迫的負面影響,而且還需要提高農作物對抗入侵雜草的競爭力。根據氣候條件對農作物品種進行定向培育和篩選則是一種合理的方案[72],諸如培育能夠耐受高溫/低溫、干旱或養分短缺等氣候變化的品種可以大大減少肥料和灌溉的投入;選擇成熟期較長的品種有利于應對極端溫度和干旱期的延長;此外,將對入侵雜草有抑制能力的品種加入農田系統也可以減少除草劑的投入[73-74]。合理調整農作物密度和均勻度對雜草生物量有顯著而持久的抑制作用,這對農作物的生物量和產量也產生了積極的作用[75]。
選擇合適的品種和種植方式將有利于環境和農業的可持續發展,因此必須擴大研究力度,研究各種非生物脅迫和種植系統下的入侵雜草如何影響農作物的品種性能和隨后的產量結果。這些信息可以納入育種計劃,以提高農作物品種在非生物(氣候變化)和生物(入侵雜草競爭)脅迫下的性能,而不會影響最終產量。
參考文獻:
[1]廖慧璇,周 婷,陳寶明,等. 外來入侵植物的生態控制[J]. 中山大學學報(自然科學版),2021,60(4):1-11.
[2]Fahad S,Bajwa A A,Nazir U,et al. Crop production under drought and heat stress:plant responses and management options[J]. Frontiers in Plant Science,2017,8:1147.
[3]Key?er L T,Lenzen M.1.5 ℃ degrowth scenarios suggest the need for new mitigation pathways[J]. Nature Communications,2021,12:2676.
[4]Li N N,Yue C,Cao H L,et al. Transcriptome sequencing dissection of the mechanisms underlying differential cold sensitivity in young and mature leaves of the tea plant (Camellia sinensis)[J]. Journal of Plant Physiology,2018,224/225:144-155.
[5]Pacanoski Z,Mehmeti A. Review:managing weed populations through alteration of the cropping pattern[J]. Agraarteadus,2020,31(1):74-83
[6]Korres N E,Norsworthy J K,Tehranchian P,et al. Cultivars to face climate change effects on crops and weeds:a review[J]. Agronomy for Sustainable Development,2016,36(1):1-12.
[7]Kumar Rai P,Singh J S. Invasive alien plant species:their impact on environment,ecosystem services and human health[J]. Ecological Indicators,2020,111:106020.
[8]全世文. 中國糧食安全戰略及其轉型[J]. 華南師范大學學報(社會科學版),2022(3):112-121,207.
[9]Muhammad I,Shalmani A,Ali M,et al. Mechanisms regulating the dynamics of photosynthesis under abiotic stresses[J]. Frontiers in Plant Science,2021,11:615942.
[10]Jagadish S V K,Way D A,Sharkey T D. Plant heat stress:concepts directing future research[J]. Plant,Cell & Environment,2021,44(7):1992-2005.
[11]Tian J,Belanger F C,Huang B R. Identification of heat stress-responsive genes in heat-adapted thermal Agrostis scabra by suppression subtractive hybridization[J]. Journal of Plant Physiology,2009,166(6):588-601.
[12]Jahan M S,Guo S R,Sun J et al. Melatonin-mediated photosynthetic performance of tomato seedlings under high-temperature stress[J]. Plant Physiology and Biochemistry,2021,167:309-320.
[13]Hu S S,Ding Y F,Zhu C. Sensitivity and responses of chloroplasts to heat stress in plants[J]. Frontiers in Plant Science,2020,11:375.
[14]Faseela P,Sinisha A K,Bresticˇ M,et al. Chlorophyll a fluorescence parameters as indicators of a particular abiotic stress in rice[J]. Photosynthetica,2020,58(SI):293-300.
[15]Sachdev S,Ansari S A,Ansari M I,et al. Abiotic stress and reactive oxygen species:generation,signaling,and defense mechanisms[J]. Antioxidants,2021,10(2):277.
[16]Jahan M S,Shu S,Wang Y,et al. Melatonin pretreatment confers heat tolerance and repression of heat-induced senescence in tomato through the modulation of ABA-and GA-mediated pathways[J]. Frontiers in Plant Science,2021,12:650955.
[17]Yang P Q,van der Tol C. Linking canopy scattering of far-red sun-induced chlorophyll fluorescence with reflectance[J]. Remote Sensing of Environment,2018,209:456-467.
[18]Cohen I,Zandalinas S I,Huck C,et al. Meta-analysis of drought and heat stress combination impact on crop yield and yield components[J]. Physiologia Plantarum,2021,171(1):66-76.
[19]Gonzalez-Rivas P A,Chauhan S S,Ha M,et al. Effects of heat stress on animal physiology,metabolism,and meat quality:a review[J]. Meat Science,2020,162:108025.
[20]Li Y Q,Kong D X,Fu Y,et al. The effect of developmental and environmental factors on secondary metabolites in medicinal plants[J]. Plant Physiology and Biochemistry,2020,148:80-89.
[21]de Vries J,de Vries S,Curtis B A,et al. Heat stress response in the closest algal relatives of land plants reveals conserved stress signaling circuits[J]. The Plant Journal,2020,103(3):1025-1048.
[22]Meena Y K,Khurana D S,Kaur N,et al. Towards enhanced low temperature stress tolerance in tomato:an approach[J]. Journal of Environmental Biology,2018,39(4):529-535.
[23]Li N,Euring D,Cha J Y,et al. Plant hormone-mediated regulation of heat tolerance in response to global climate change[J]. Frontiers in Plant Science,2021,11:627969.
[24]Hall A E. Crop developmental responses to temperature,photoperiod,and light quality[M]//Crop responses to environment.Boca Raton:CRC Press,2018:95-107.
[25]Impa S M,Raju B,Hein N T,et al. High night temperature effects on wheat and rice:current status and way forward[J]. Plant,Cell & Environment,2021,44(7):2049-2065.
[26]Kumari V V,Roy A,Vijayan R,et al. Drought and heat stress in cool-season food legumes in sub-tropical regions:consequences,adaptation,and mitigation strategies[J]. Plants,2021,10(6):1038.
[27]El Yaacoubi? A,Oukabli A,Legave J. Response of almond flowering and dormancy to Mediterranean temperature conditions in the context of adaptation to climate variations[J]. Scientia Horticulturae,2019,257:108687.
[28]Frank G,Pressman E,Ophir R,et al. Transcriptional profiling of maturing tomato (Solanum lycopersicum L.) microspores reveals the involvement of heat shock proteins,ROS scavengers,hormones,and sugars in the heat stress response[J]. Journal of Experimental Botany,2009,60(13):3891-3908.
[29]Krishna Jagadish S V.Heat stress during flowering in cereals-effects and adaptation strategies[J]. New Phytologist,2020,226(6):1567-1572.
[30]Khan A,Ahmad M,Ahmed M,et al. Rising atmospheric temperature impact on wheat and thermotolerance strategies[J]. Plants,2020,10(1):43.
[31]Haque M S,Husna M T,Uddin M N,et al. Heat stress at early reproductive stage differentially alters several physiological and biochemical traits of three tomato cultivars[J]. Horticulturae,2021,7(10):330.
[32]Jain M,Vara Prasad P V,Boote K J,et al. Effects of season-long high temperature growth conditions on sugar-to-starch metabolism in developing microspores of grain Sorghum (Sorghum bicolor L. Moench)[J]. Planta,2007,227(1):67-79.
[33]Chinnusamy V,Zhu J H,Zhu J K. Cold stress regulation of gene expression in plants[J]. Trends in Plant Science,2007,12(10):444-451.
[34]Zhang Z B,Hu M H,Xu W W,et al. Understanding the molecular mechanism of anther development under abiotic stresses[J]. Plant Molecular Biology,2021,105(1):1-10.
[35]Raza A,Charagh S,García-Caparrós P,et al. Melatonin-mediated temperature stress tolerance in plants[J]. GM Crops & Food,2022,13(1):196-217.
[36]Sun Y J,Zhou J,Guo J S.Advances in the knowledge of adaptive mechanisms mediating abiotic stress responses in Camellia sinensis[J]. Frontiers in Bioscience-Landmark,2021,26(12):1714-1722.
[37]Guo X Y,Liu D F,Chong K.Cold signaling in plants:insights into mechanisms and regulation[J]. Journal of Integrative Plant Biology,2018,60(9):745-756.
[38]Liu J Y,Shi Y T,Yang S H.Insights into the regulation of C-repeat binding factors in plant cold signaling[J]. Journal of Integrative Plant Biology,2018,60(9):780-795.
[39]Amparo P,MaryRus M,?ngeles F M. Cold stress in Citrus:a molecular,physiological and biochemical perspective[J]. Horticulturae,2021,7(10):340.
[40]Ermilova E.Cold stress response:an overview in Chlamydomonas[J]. Frontiers in Plant Science,2020,11:569437.
[41]Wei X S,Liu S,Sun C,et al. Convergence and divergence:signal perception and transduction mechanisms of cold stress in Arabidopsis and rice[J]. Plants,2021,10(9):1864.
[42]Yamori W,Noguchi K,Hikosaka K,et al. Phenotypic plasticity in photosynthetic temperature acclimation among crop species with different cold tolerances[J]. Plant Physiology,2009,152(1):388-399.
[43]Zhong P,Zhou M,Zhang J J,et al. The role of cold-inducible RNA-binding protein in respiratory diseases[J]. Journal of Cellular and Molecular Medicine,2022,26(4):957-965.
[44]Bhat K A,Mahajan R,Pakhtoon M M,et al. Low temperature stress tolerance:an insight into the omics approaches for legume crops[J]. Frontiers in Plant Science,2022,13:888710.
[45]Rachappanavar V,Padiyal A,Sharma J K,et al. Plant hormone:mediated stress regulation responses in fruit crops:a review[J]. Scientia Horticulturae,2022,304:111302.
[46]Ritonga F N,Chen S.Physiological and molecular mechanism involved in cold stress tolerance in plants[J]. Plants,2020,9(5):684-689.
[47]Ku Y S,Sintaha M,Cheung M Y,et al. Plant hormone signaling crosstalks between biotic and abiotic stress responses[J]. International Journal of Molecular Sciences,2018,19(10):3206.
[48]Tonkinson C L,Lyndon R F,Arnold G M,et al. The effects of temperature and the Rht3 dwarfing gene on growth,cell extension,and gibberellin content and responsiveness in the wheat leaf[J]. Journal of Experimental Botany,1997,48(4):963-970.
[49]Kang H M,Saltveit M E.Chilling tolerance of maize,cucumber and rice seedling leaves and roots are differentially affected by salicylic acid[J]. Physiologia Plantarum,2002,115(4):571-576.
[50]Mora-Herrera M E,Lopez-Delgado H,Castillo-Morales A,et al. Salicylic acid and H2O2function by independent pathways in the induction of freezing tolerance in potato[J]. Physiologia Plantarum,2010,125(4):430-440.
[51]Balah M A,Balah A M.Growth and ecological characteristics of Physalis angulata invasive weed species in several invaded communities[J]. Biologia,2022,77(2):325-338.
[52]Dweikat I.A diploid,interspecific,fertile hybrid from cultivated sorghum,Sorghum bicolor,and the common johnsongrass weed Sorghum halepense[J]. Molecular Breeding,2005,16(2):93-101.
[53]Yadav S,Hemke A,Umekar M. Convolvulaceae:a morning glory plant[J]. International Journal of Pharmaceutical Sciences Review and Research,2018,51(1):103-117.
[54]Ziska L H,Bunce J A.Predicting the impact of changing CO2on crop yields:some thoughts on food[J]. New Phytologist,2007,175(4):607-618.
[55]Malhi G S,Kaur M,Kaushik P. Impact of climate change on agriculture and its mitigation strategies:a review[J]. Sustainability,2021,13(3):1318.
[56]Riar D S,Norsworthy J K,Steckel L E,et al. Assessment of weed management practices and problem weeds in the midsouth United States—soybean:a consultant's perspective[J]. Weed Technology,2013,27(3):612-622.
[57]Ziska L,Runion G B.Future weed,pest,and disease problems for plants[M]//Newton P C D,Carran R A,Edwards G R,et al.Agroecosystems in a changing climate.Boca Raton:CRC Press,2006:261-287.
[58]Jabran K,Dogˇan M N.Elevated CO2,temperature and nitrogen levels impact growth and development of invasive weeds in the Mediterranean region[J]. Journal of the Science of Food and Agriculture,2020,100(13):4893-4900.
[59]Bhusal D R,Ghimire K C,Patel P,et al. Temperature and altitude modulate feeding attributes of Mexican beetle,Zygogramma bicolorata Pallister on Parthenium hysterophorus[J]. Journal of Thermal Biology,2020,89:102540.
[60]Ruchel Q,Zandoná R R,Fraga D S,et al. Effect of high temperature and recovery from stress on crop-weed interaction[J]. Bragantia,2020,79(4):582-591.
[61]Vilà M,Beaury E M,Blumenthal D M,et al. Understanding the combined impacts of weeds and climate change on crops[J]. Environmental Research Letters,2021,16(3):034043.
[62]Ziska L H.Climate change and the herbicide paradigm:visiting the future[J]. Agronomy,2020,10(12):1953.
[63]Ding Y L,Shi Y T,Yang S H.Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants[J]. New Phytologist,2019,222(4):1690-1704.
[64]Morales F,Ancín M,Fakhet D,et al. Photosynthetic metabolism under stressful growth conditions as a bases for crop breeding and yield improvement[J]. Plants,2020,9(1):88.
[65]Schlenker W,Roberts M J.Nonlinear temperature effects indicate severe damages to U.S.crop yields under climate change[J]. Proceedings of the National Academy of Sciences of the United States of America,2009,106(37):15594-15598.
[66]Lobell D B,Field C B.California perennial crops in a changing climate[J]. Climatic Change,2011,109(1):317-333.
[67]Raza A,Razzaq A,Mehmood S S,et al. Impact of climate change on crops adaptation and strategies to tackle its outcome:a review[J]. Plants,2019,8(2):34.
[68]Wang H Z,Zhao K P,Li X J,et al. Factors affecting seed germination and emergence of Aegilops tauschii[J]. Weed Research,2020,60(3):171-181.
[69]Chen J Y,Burns E,Fleming M,et al. Impact of climate change on population dynamics and herbicide resistance in kochia [Bassia scoparia (L.) A.J.Scott][J]. Agronomy,2020,10(11):1700.
[70]Gandía M L,Casanova C,Sánchez F J,et al. Arable weed patterns according to temperature and latitude gradient in central and southern Spain[J]. Atmosphere,2020,11(8):853.
[71]Karkanis A,Ntatsi G,Alemardan A,et al. Interference of weeds in vegetable crop cultivation,in the changing climate of Southern Europe with emphasis on drought and elevated temperatures:a review[J]. Journal of Agricultural Science,2018,156(10):1175-1185.
[72]Xie H J,Han Y H,Li X Y,et al. Climate-dependent variation in cold tolerance of weedy rice and rice mediated by OsICE1 promoter methylation[J]. Molecular Ecology,2020,29(1):121-137.
[73]Kniss A R.Genetically engineered herbicide-resistant crops and herbicide-resistant weed evolution in the United States[J]. Weed Science,2018,66(2):260-273.
[74]Holmes K H,Lindquist J L,Rebarber R,et al. Modeling the evolution of herbicide resistance in weed species with a complex life cycle[J]. Ecological Applications,2022,32(1):e02473.
[75]Kumar J S,Nedunchezhiyan M,Sunitha S. Weed control approaches for tropical tuber crops:a review[J]. International Journal of Vegetable Science,2021,27(5):439-455.
