寒地蘋果資源遺傳多樣性及群體結構的SSR 標記分析*
2023-07-17 11:16:24郭勁鵬胡穎慧宋宏偉卜海東于文全顧廣軍
中國果樹 2023年5期
關鍵詞:資源
劉 暢,郭勁鵬,高 源,胡穎慧,楊 悅,宋宏偉,卜海東,于文全,王 昆,顧廣軍
(1 黑龍江省農業科學院牡丹江分院,寒地果樹育種與栽培重點實驗室, 157000)(2 黑龍江省林業科學院牡丹江分院)(3 中國農業科學院果樹研究所,農業農村部園藝作物種質資源利用重點實驗室)(4 吉林省農業科學院果樹研究所)
東北屬于寒地小蘋果栽培區,以楸子、山定子、海棠及綿蘋果等小蘋果為主,通過引種并同當地野生種進行雜交育種,形成了北方寒地特有的蘋果資源群落[1]。大量的蘋果資源之間雜交使得群體間基因交流頻繁,導致表型變異幅度增大,增加了種質資源的遺傳背景和親緣關系的鑒定難度。目前對于寒地蘋果資源的鑒定大多從葉片、枝條、果實等形態指標進行[2-5],由于受環境影響較大而影響結果的準確性。
隨著分子技術的發展,SSR 標記具有多態性高、重復性好以及準確性高等特點,已成為蘋果資源遺傳多樣性研究的主要方法[6-7]。Gharghani 等[8]利用篩選出的9 對SSR 位點對159 份蘋果資源進行遺傳多樣性分析,發現伊朗地方蘋果品種與俄羅斯的東方海棠關系比其他海棠品種更近,來自世界不同地區的老蘋果與西方蘋果、東方蘋果和伊朗蘋果的遺傳關系比其他野生品種更為密切;Gasia 等[9]利用10對SSR 引物對24 份波黑蘋果品種和15 個國外品種進行遺傳多樣性分析,發現2 個傳統的本國品種與國外品種緊密地聚類在一起,而其他品種形成了獨立的聚類;Zhang 等[10]利用29 個SSR 引物對19 份克什米爾地區蘋果材料遺傳多樣性進行分析,共擴增出218 個多態性等位基因,同時發現所有蘋果品種主要分為2 個大類,存在更廣泛的遺傳多樣性。
目前,針對中國北方寒地蘋果資源群體多樣性研究較少。本研究用篩選出多態性好的20 對熒光SSR 引物,對49 份寒地蘋果種質資源進行SSR 多態性分析、聚類分析和群體結構分析,旨在揭示寒地蘋果資源遺傳多樣性特征和群體遺傳分化結構,為寒地蘋果資源收集利用、親緣關系分析和分子育種提供理論依據。
1 材料與方法
1.1 試驗材料
供試49 份寒地蘋果資源材料見表1,其中21份為在黑龍江野外對蘋果資源收集考察時獲得,6份為在俄羅斯遠東地區考察時獲得,13 份取自國家果樹種質公主嶺寒地果樹圃,其余9 份取自黑龍江省農業科學院牡丹江分院蘋果資源圃。
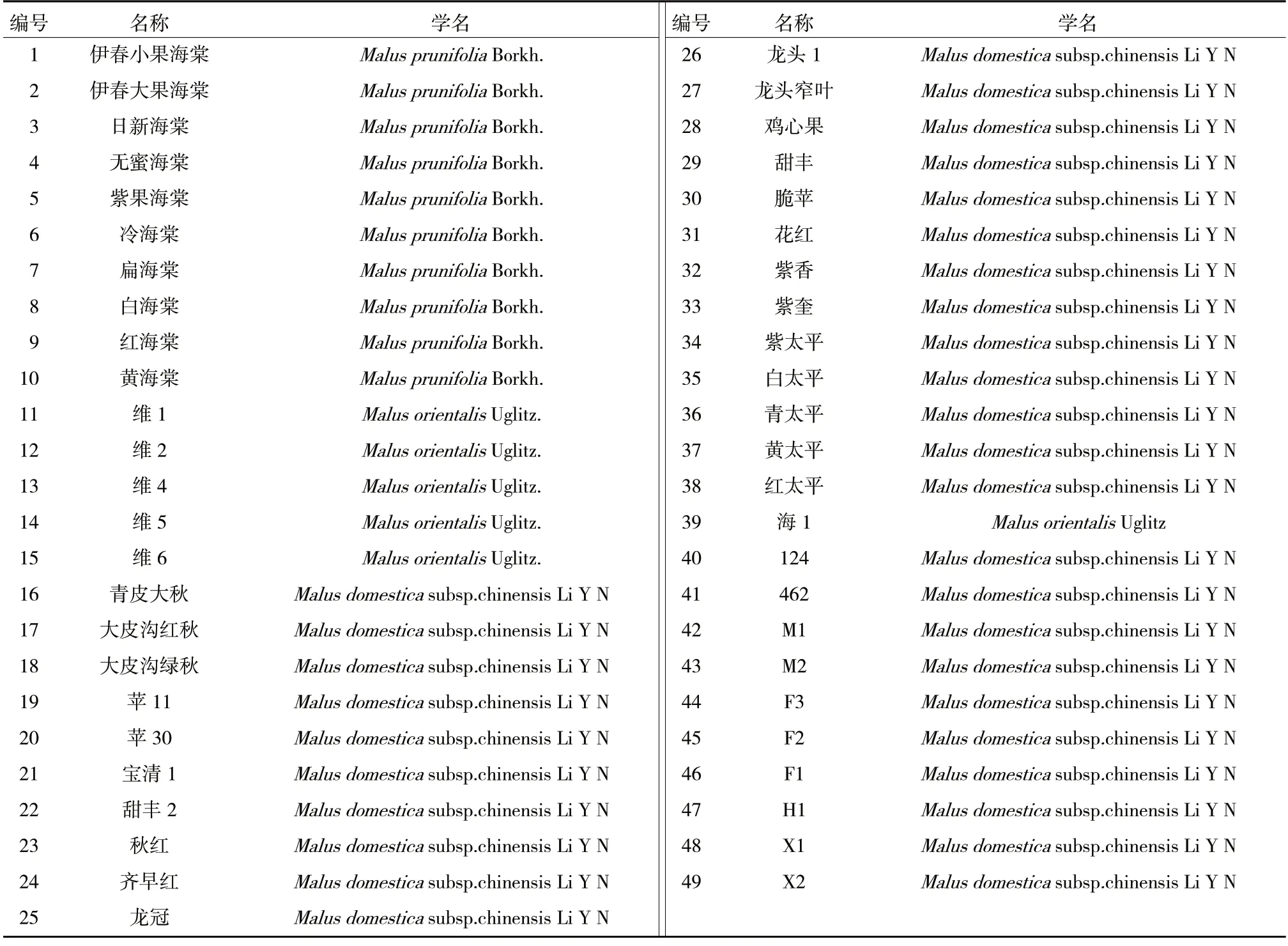
表1 49 份寒地蘋果資源
1.2 試驗方法
2020 年5 月采集幼嫩葉片,采用德國QIAGEN的DNeasy Plant Mini Kit 提取DNA,1%瓊脂糖凝膠電泳檢測DNA 完整性,用紫外分光光度計檢測其濃度。從Hokanson 等[11]、Liebhard 等[12]、Yamamoto等[13]和Guiford 等[14]報道的序列中選取擴增產物片段長度在100~300 bp 并經檢測具有高度多態性的SSR 引物20 對,引物相關信息見表2。SSR 反向引物和帶有6FAMTM 熒光標記的SSR 正向引物均由上海生工有限公司合成。
PCR 體系參照Cao 等[15]的方法,擴增產物的純化體系參照高源等[16]的方法。PCR 反應在Bio-Rad PTC-200 上進行。熒光SSR 的PCR 擴增產物經過純化后在美國ABI-3730 基因測序儀上進行熒光檢測,收集原始數據。
1.3 數據統計與分析
利用GeneMapper 3.0 軟件對ABI-3730 收集數據進行分析,獲得樣品在每個SSR 位點的擴增片段長度,即指紋數據。利用遺傳數據分析軟件GenAlEx 6.501 計算多態性等位基因數(Na)、SSR 位點的有效等位基因數(Ne)、觀察雜合度(Ho)、期望雜合度(He)、固定指數(F)及香農多樣性指數(I)等遺傳多樣性指標,并分析種群間的分子變異(AMOVA)。基于SSR 不同位點,通過Excel 2017構建(0,1)矩陣,利用NTSYS 1.2 構建49 份寒地蘋果材料的進化樹,使用STRUCTURE 2.3.4 分析群體的遺傳結構并確定最佳的群體分組。
2 結果與分析
2.1 SSR 擴增產物多態性分析
利用20 對多態性熒光SSR 引物對49 份寒地蘋果資源基因組DNA 進行擴增,結果見表3,共擴增出278 個多態性等位基因(Na)。多態性等位基因數最少為3.000(GD147),最多為21.000(CH01f07a和CH01f02),平均多態性等位基因數為13.850,多態性位點百分率為100%。有效等位基因數(Ne)為1.353(NZ28f4)~12.637(CH01f07a),平均值為6.649。香農多樣性指數(I)為0.646(NZ28f4)~2.745(CH01f07a),平均值為2.065。觀察雜合度(Ho)為0.000(GD147)~0.776(GD15),平均值為0.569。期望雜合度(He)為0.261(NZ28f4)~0.921(CH01f07a),平均值為0.799。固定指數(F)為-0.026(GD15)~1.000(GD147),平均值為0.296,只有在GD15 中為負值,其余均為正值,說明供試寒地蘋果資源群體內含有雜合子較少。
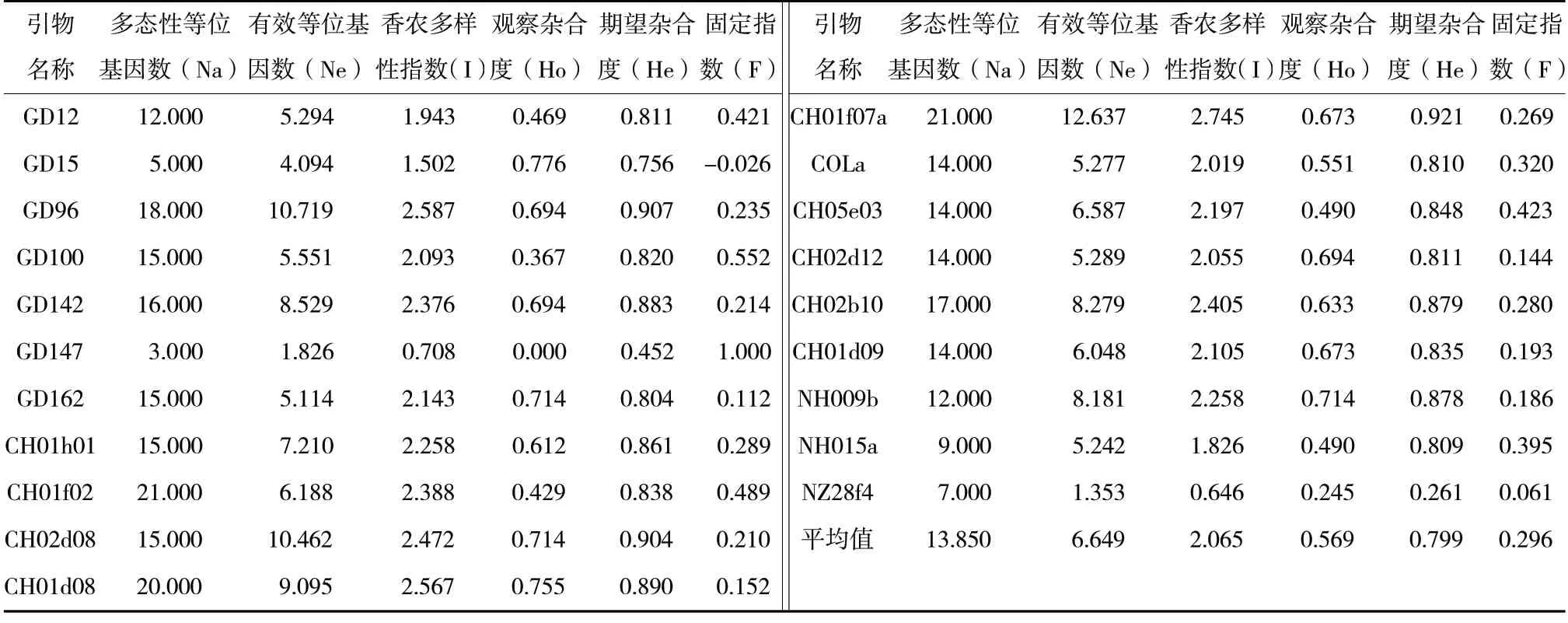
表3 不同SSR 位點的遺傳多樣性特征
2.2 不同蘋果群體的遺傳多樣性和方差分析
49 份寒地蘋果資源分屬于3 個不同的群體(表4),群體分析發現中國蘋果的多態性等位基因數、有效等位基因數和香農多樣性指數最多,分別為12.750、6.885、2.054;觀察雜合度楸子群體最高,為0.600,西洋蘋果群體最低,為0.480;期望雜合度全部群體均高于0.500,中國蘋果群體最高,為0.803,西洋蘋果群體最低,為0.635,整體上遺傳多樣性較高;期望雜合度與觀察雜合度差值最小的為楸子群體,差值最大的為中國蘋果群體;固定指數在各個群體中均為正值,其中在楸子群體中最小,為0.144,與其他兩個群體相比雜合子較多。對3 個群體進行群體分子遺傳變異方差分析,群體內方差分量的貢獻率占28%,個體內方差分量的貢獻率占69%,遺傳變異顯著;群體間方差分量的貢獻率占3%,遺傳變異不顯著。
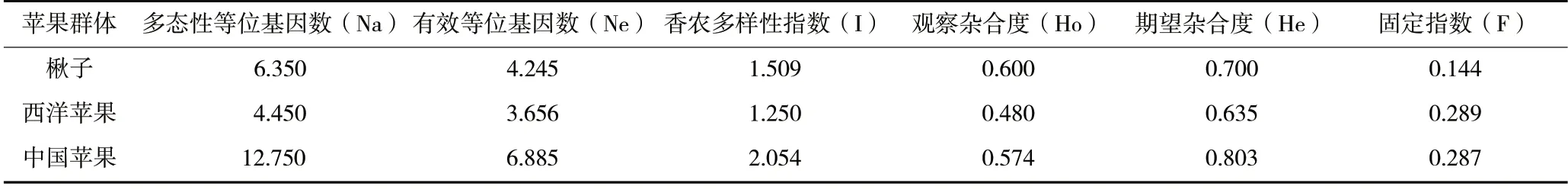
表4 3 個蘋果群體的遺傳多樣性
2.3 群體間的遺傳分化
按照所屬種劃分群體后計算Nei 遺傳距離和遺傳分化系數(表5),楸子群體與西洋蘋果群體間遺傳距離最大,為0.411;其次是西洋蘋果群體與中國蘋果群體,為0.305;楸子群體與中國蘋果群體間遺傳距離最小,為0.261。不同群體之間遺傳分化系數為0.047~0.081,西洋蘋果群體與楸子群體遺傳分化系數最高,為0.081;西洋蘋果群體與中國蘋果群體遺傳分化系數居中,為0.058;中國蘋果群體與楸子群體遺傳分化系數最低,為0.047;體現了不同群體地理位置和遺傳距離的相關性。
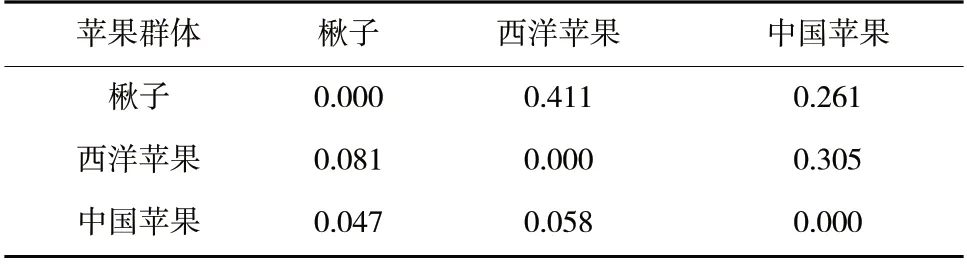
表5 寒地3 個蘋果群體間的遺傳距離和遺傳分化系數
2.4 寒地蘋果資源聚類分析
基于20 對引物對49 份供試材料的SSR 數據進行UPGMA 聚類分析(圖1),遺傳距離為0.80~1.00,表明各種質間親緣關系差異較大。在遺傳距離0.80 處供試的49 份蘋果資源分為2 個大類:第Ⅰ類包括18 份資源,全部是中國蘋果群體;第Ⅱ類包括31 份資源,其中楸子群體10 份,中國蘋果群體16 份,西洋蘋果群體5 份。在遺傳距離0.81處,第Ⅰ類進一步分為Ⅰa 和Ⅰb 2 個亞類。其中Ⅰa 亞類包括124、甜豐、脆蘋、花紅、紫奎5 份資源;Ⅰb 亞類含有13 份資源,進一步細分可以看出青皮大秋、大皮溝紅秋、大皮溝綠秋3 份資源聚為一類,雞心果、462、紫香3 份資源聚為一類,龍頭、龍頭窄葉等7 份資源聚為一類。
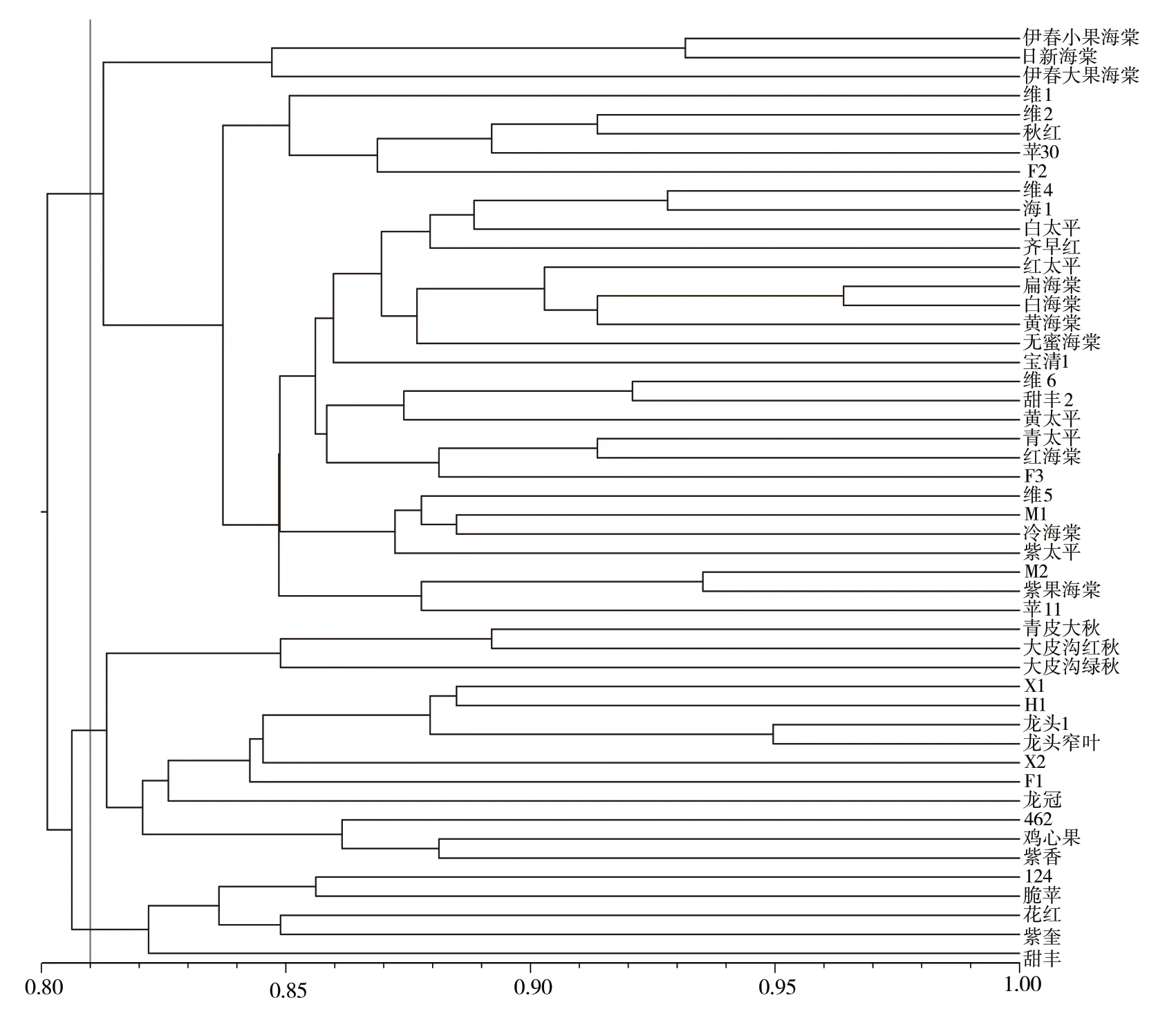
圖1 49 份寒地蘋果資源的UPGMA 聚類圖
2.5 群體遺傳結構分析
基于SSR 分子標記數據對供試的49 份寒地蘋果資源進行群體結構分析,參照Evanno 等[17]的方法來確定K 值。設置推測群體數K 為1~8,重復10次,當K=2 時,ΔK 取得最大值(圖2),隨后急劇下降,推測49 份蘋果資源由于基因型不同分為2個類群(圖3)。不同群體的材料在2 個類群中均有分布,類群1 含有21 份資源,包括楸子群體3份、中國蘋果群體13 份和西洋蘋果群體5 份。類群2 含有28 份資源,包括楸子群體7 份、中國蘋果群體21 份。群體結構分析結果與聚類結果相類似,UPGMA 聚類Ⅰ類中除去中國蘋果群體6 份全部歸于STRUCTURE 分組類群2 中,UPGMA 聚類Ⅱ類中楸子群體10 份中3 份、中國蘋果群體16 份中7 份和西洋蘋果群體5 份歸于STRUCTURE 分組類群1 中。
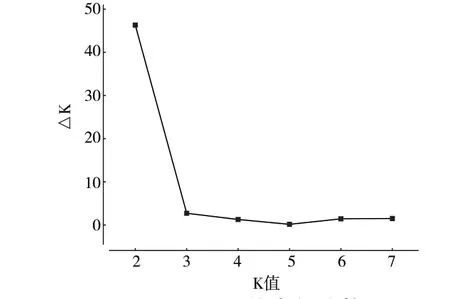
圖2 ΔK 值變化趨勢圖
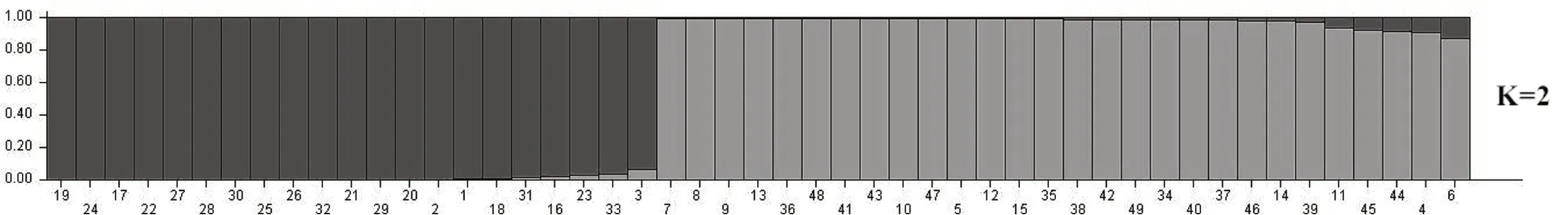
圖3 K=2 時,49 份寒地蘋果資源的遺傳結構分析圖
3 討論
3.1 寒地蘋果資源遺傳多樣性
SSR 分子標記技術在揭示蘋果屬不同群體間遺傳差異具有廣泛應用。Kumar 等[18]利用31 個SSR 位點對喜馬拉雅海棠進行遺傳多樣性研究,共檢測出96 個位點,每個SSR 位點的平均等位基因數為3.29個。Potts 等[7]利用10 個SSR 分子標記對164 份蘋果資源進行擴增,共擴增出283 個等位基因,其中稀有等位基因百分率為39%。Ana 等[19]利用20 個SSR分子標記技術對西班牙東北部地區的183 份蘋果種質進行遺傳多樣性研究,發現不同群體中多態位點的檢出率為100%。出現差異的原因可能與所用標記引物和供試群體類型相關。本研究利用20 對SSR引物對49 份寒地蘋果資源進行遺傳多樣性分析,共擴增出278 個多態性等位基因,平均多態性等位基因數為13.850,每對引物在群體中擴增出的多態位點的百分率為100%,表明供試寒地蘋果不同群體間基因多態性豐富。
有效等位基因數(Ne)、香農多樣性指數(I)、期望雜合度(He)等是評價群體變異程度高低的重要指標。本研究中,49 份寒地蘋果資源總體遺傳多樣性的期望雜合度(He=0.799)、香農多樣性指數(I=2.065)和有效等位基因數(Ne=6.649)均高于變葉海棠[20](He=0.438 9,I=0.628 2,Ne=1.81)、湖北海棠[21](He=0.262 8,I=0.401 5,Ne=1.437 5)和三葉海棠[22](He=0.699,I=1.458,Ne=3.954)等蘋果屬植物。Lassois 等[23]發現,雜合度大于0.5的群體具有較高的遺傳多樣性。本研究中49 份寒地蘋果資源的期望雜合度(He)均高于0.5,遺傳多樣性相對較高。雜合度觀測值和雜合度期望值的相似程度也反映了群體的遺傳多樣性,差值越小,遺傳多樣性越高。本研究中楸子差值小于西洋蘋果和中國蘋果,說明楸子群體遺傳多樣性高于其他兩個群體。固定指數大小也能體現群體的遺傳差異,固定指數為正值且數值越大,多樣性越小。本研究結果顯示楸子群體的固定指數最小,為0.144,與其他兩個群體相比雜合子較多。Urrestarazu 等[24]利用16 對SSR 引物對493 份本地品種和45 個參考品種共擴增出267 個等位基因,其中參考品種僅占56%。Wasim 等[25]基于SSR 對喜馬拉雅北部寒冷地區29 份蘋果資源的遺傳結構分析發現,種群內遺傳變異占93%,而種群間的遺傳變異僅占7%。本研究顯示,49 份寒地蘋果資源群體間變異占3%,顯著低于前人研究結果,但整體趨勢均顯示變異主要存在于群體內,而不是群體間。本研究中楸子群體多樣性指標(He=0.700,I=1.509,Ne=4.245)均低于高源等[26]對楸子種質資源的遺傳多樣性研究(He=0.868,I=2.446,Ne=9.399)。其原因一方面是材料全部采自中國東北,地域過于集中;另一方面群體數量太少,可進一步增加品種來進行驗證。鑒于楸子大多為地方栽培品種且分布廣泛,建議在寒地蘋果育種中加大對楸子群體的考察收集,豐富群體的遺傳多樣性。
3.2 寒地蘋果資源遺傳結構
群體間遺傳分化是評價群體變異的重要指標,Fst 值的高低反映群體間的遺傳差異程度。Wright[27]認為,群體Fst 值在0~0.05,各種群間不存在分化;Fst 值在0.05~0.15 為中度分化;Fst 值在0.15~0.25為高度分化。本研究中楸子群體與中國蘋果群體間遺傳距離最小,為0.261,Fst 值最低,為0.047,群體間不存在分化;楸子群體與西洋蘋果群體間遺傳距離最大,為0.411,Fst 值最高,為0.081,其群體間屬于中度分化。推測原因是寒地蘋果品種選育中優先選擇當地楸子作親本與蘋果雜交,存在基因滲透且交流頻繁,導致楸子群體與中國蘋果群體間無分化。發現兩個群體之間的遺傳距離與遺傳差異相關,這與高源等[28]在對栽培種楸子的遺傳多樣性上面的研究結果一致。
為進一步明確各群體遺傳結構,對49 份寒地蘋果資源進行UPGMA 聚類,發現在遺傳距離0.80處被分為2 個大類,與STRUCTURE 群體結構分析結果一致,但是在同類內部兩種遺傳關系分析結果并不完全一致,第Ⅰ類全部為中國蘋果,其中雞心果、甜豐、脆蘋、花紅、紫香、紫奎在群體結構中與楸子、西洋蘋果歸為一類,第Ⅱ類中楸子全部聚為一類,其中伊春小果海棠、伊春大果海棠、日新海棠在群體結構中與中國蘋果和西洋蘋果聚為一類,楸子群體與中國蘋果群體聚類比較緊密,又與西洋蘋果群體相互交錯,各種群遺傳距離和類群歸屬與地理位置不完全相關。Silva 等[29]研究發現,種群的雜交方式對種群分布有重要影響,所以我們推測俄羅斯遠東地區和黑龍江省相鄰,楸子群體大多分布在人類居住區附近,極易受到人為擴散產生屬內自然雜交,導致基因滲透。同時本研究中采集各群體數量不一致對聚類結果可能會有一定影響,因此應加大不同種群蘋果資源收集力度來完善遺傳結構分析結果。
4 結論
本研究通過20 對多態性SSR 引物開展蘋果群體的遺傳多樣性分析,49 份寒地蘋果資源總體遺傳多樣性較高,中國蘋果群體遺傳多樣性最高,西洋蘋果群體遺傳多樣性最低,遺傳變異主要存在于種群內的個體間(28%)和個體內(69%)。49 份寒地蘋果資源共分為2 個大類,不同資源相互交錯,結果表明遺傳距離和類群歸屬與地理位置不完全相關,各群體間存在基因滲透和遺傳分化。
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
資源節約與環保(2022年8期)2022-09-20 02:25:22
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
藝術品鑒(2020年7期)2020-09-11 08:04:44
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
當代貴州(2018年28期)2018-09-19 06:39:04
資源再生(2017年3期)2017-06-01 12:20:59
決策(2015年9期)2015-09-10 07:22:44
