山羊α-干擾素基因克隆、原核表達及其抗病毒活性研究
2023-07-20 09:17:00徐菁昱劉艷芬陳紹紅
西北農業學報 2023年8期
徐菁昱, 劉艷芬, 陳紹紅,劉 鈾
(1.廣東海洋大學 濱海農業學院,廣東湛江 524088;2.廣東海洋大學 食品科技學院,廣東湛江 524088)
中國是世界上養羊數量最多的國家,羊肉總產量居世界首位[1]。近年來,隨著規模化養羊業的快速發展,傳染病尤其是病毒性傳染病成為制約中國養羊業健康發展的重要因素[2]。流行病學調查顯示,嚴重危害山羊健康的疾病多達54種,傳染病占64.8%,其中病毒性傳染病占31.4%,且多種病原體混合感染的情況較為普遍[3]。因此,通過疫苗接種控制傳染病暴發流行的同時,積極研究開發廣譜、高效、安全的抗病毒制劑對于山羊病毒性傳染病的防治有著十分現實的意義。
審美趣味的改變制約了水墨動畫的進一步發展。上世紀80年代以后,西方審美標準的盛行改變了動畫受眾的審美趣味,繼而影響了國產動畫的創作風格,水墨動畫隨即成為過眼煙云。隨著社會的發展和進步,在高科技迅速發展的時代,中國的日新月異,各種先進的動畫技術以及西方審美思想的影響,從制作技術、審美趣味到藝術風格上,中國的動畫片逐漸被“西化”,喪失了本民族的特色和優勢,外來動畫風格成為動畫創作的主要風格。
干擾素(interferon,IFN)是在病毒或其他抗原刺激下由白細胞和淋巴細胞分泌的一類糖蛋白[4],具有廣譜抗病毒、抗腫瘤和免疫調節等多種生物學活性[5-6],其中α-干擾素的抗病毒活性最強[7]。采用體外培養外周血單核細胞、病毒或絲裂原誘導是制備天然干擾素的常規方法,但干擾素產量低,純化不易,且生產成本較高,極大地限制了干擾素的臨床應用。利用基因工程技術生產重組干擾素是解決這一難題的有效途徑。目前,重組α-干擾素制劑已在醫學臨床中得到廣泛應用,在人類病毒性疾病的預防和治療中發揮了重要作用[8]。近年來,重組干擾素已應用于犬、貓、豬、雞等病毒性傳染病的預防和治療[9]。目前,有關雷州山羊α-干擾素基因的遺傳變異、重組山羊α-干擾素的抗病毒活性的報道較少,市場亦無商品化山羊干擾素產品。
本研究通過克隆雷州山羊α-干擾素基因,分析其遺傳變異,將其成熟肽編碼序列導入大腸桿菌表達,制備重組α-干擾素成熟肽并檢測其抗PPRV和GPV的活性,旨在為山羊病毒性傳染病的防治提供新的途徑。
1 材料與方法
1.1 材料及主要試劑
雷州山羊來自廣東海洋大學雷州山羊保種場;質粒pET-32a(+)、大腸桿菌(Escherichiacoli)菌株DH5α和BL21(DE3)、Vero細胞、LDG-2細胞、小反芻獸疫病毒疫苗株Nigeria 75/1(Peste des petits ruminants virus,PPRV)、山羊痘病毒疫苗株AV41(Goat pox virus,GPV)均由廣東海洋大學生化中心保存。
雷州山羊α-干擾素基因與豬(NM_214393)、人(NM_024013)和鼠(NM_010502)的核苷酸序列同源性較低,分別為81.6%、77.8%和70.0%,氨基酸序列同源性分別為67.9%、61.6%和 55.3%(表1),說明與反芻動物親緣關系較遠的動物種屬之間α-干擾素基因序列存在較大差異。
1.2 引物設計
根據GenBank中山羊IFN-α基因序列(登錄號:FJ959074)設計引物,F1:5′-ATGGCCCCAGCCTGGTCCTTA-3′,R1:5′-TCAGTCCTTCCTCCTGAATCTCTCC-3′用于擴增α-干擾素全基因;F2:5′-CATGCCATGGCAGGCTGCCACCTGCCTCACATCCA-3′(NcoⅠ),R2:5′-CCCAAGCTTTCAGTCCTTCCTCCTGAAT CT-3′(HindⅢ)用于擴增α-干擾素成熟肽編碼序列。引物由上海生工生物工程股份有限公司 合成。
1.3 山羊α-干擾素全基因的克隆
利用DNA凝膠回收試劑盒純化目的基因片段后,與pMD19-T載體于16 ℃連接過夜,連接產物用于轉化DH5α感受態細胞。挑取陽性克隆作增菌培養,以菌液為模板,按上述條件進行PCR擴增。陽性轉化子用質粒DNA提取試劑盒提取質粒,分別用HindⅢ和HindⅢ+BamH Ⅰ 酶切消化,消化產物用10 g/L瓊脂糖電泳鑒定。經PCR和酶切鑒定的重組質粒由上海生工生物工程股份有限公司測定目的基因核苷酸序列。重組質粒命名為pMD19-T-IFN-α。
在25 μL反應體系中加入RNA模板5 μL、dNTP(10 mmol/L)1 μL、Oligo dT 1 μL、5×RT Buffer 5 μL、Rnase Inhibitor 0.5 μL、M-MLV RT 1 μL、DEPC水11.5 μL,采用反轉錄制備α-干擾素全基因cDNA。反應條件為: 65 ℃ 5 min,42 ℃ 50 min,70 ℃ 5 min。在20 μL反應體系中加入cDNA 5 μL,F1/R1各1 μL,dNTP 0.4 μL,Taq酶0.2 μL,10×Buffer 2 μL,ddH2O 10.4 μL,采用PCR方法擴增山羊IFN-α全基因。反應條件為:94 ℃ 3 min;94 ℃ 40 s, 56 ℃ 40 s,72 ℃ 40 s,32 個循環,72 ℃ 延伸 10 min。PCR產物用10 g/L瓊脂糖凝膠電泳 檢測。
無菌采集健康雷州山羊靜脈血,用外周血淋巴細胞分離液分離外周血單核細胞,按2×106mL-1接入細胞培養瓶,加入終質量濃度為5 μg/mL的植物血凝素(PHA),于37 ℃、5% CO2條件下培養24 h后,3 500 r/min離心收集細胞,采用Trizol 試劑提取細胞總RNA,瓊脂糖電泳檢測RNA完整性,紫外吸收法測定RNA含量。
1.4 山羊α-干擾素表達載體的構建
采用微量細胞病變抑制法檢測重組山羊α-干擾素的抗病毒活性[13]。將Vero細胞和LDG-2細胞分別接種96孔板,待細胞長至單層,試驗組細胞分別加入用維持液進行4倍倍比稀釋的重組山羊α-干擾素100 μL,每個稀釋度8個重復,細胞對照組和病毒對照組分別加入100 μL維持液,于37 ℃、5% CO2條件下培養。24 h后,試驗組和病毒對照組的Vero細胞按100 μL/孔分別加入100TCID50的PPRV懸液,試驗組和病毒對照組LDG-2細胞則按100 μL/孔分別加入100TCID50的GPV懸液。每12 h觀察細胞病變情況,待病毒對照組出現明顯細胞病變,將試驗組中抑制50%細胞病變的干擾素最高稀釋度確定為1個干擾素活性單位。
1.5 重組山羊α-干擾素基因的原核表達及表達條件優化
——近期摩根士丹利發布了一份報告,揭示了澳大利亞葡萄酒出口中國的強勁勢頭。但報告也發現,雖然越來越多的澳大利亞葡萄酒出口到中國,但按價值計算,出口到中國的高檔葡萄酒數量仍然占比較低。同時,一些小酒廠的葡萄酒質量上乘,但價格并不低。且中間環節層層累加,到中國消費者手中時價格畸高。
薛宇航:這篇作文寫完后,我自己讀了兩遍,感覺還不錯,嚴老師也表揚了我,說寫得不錯。看過兩位“小編輯”的修改之后,我覺得他們改得真好,我的這篇作文,果然還有很大的提升空間。看來,以后我寫完作文,也要注意多修改了。
1.6 重組山羊α-干擾素融合蛋白的分離與純化
根據“1.5”的檢測結果,用優化后的條件誘導表達目的蛋白。破碎細胞后收集沉淀,獲得包涵體。用2 mol/L尿素洗滌、8 mol/L尿素溶解包涵體后,稀釋變性液至尿素終濃度為4 mol/L后置于透析袋中,分別用3、2、1、0.5 mol/L尿素梯度透析復性[11-12]。復性蛋白用Ni-NAT親和層析柱純化,用10K超濾管濃縮目的蛋白,并以PBS緩沖液洗脫。所得蛋白質樣品經Western-blot鑒定、BCA蛋白測定試劑盒測定蛋白濃度后,用 0.22 μm濾膜過濾除菌分裝,-80 ℃保存,備用。
1.7 重組山羊 α-干擾素的抗病毒活性測定
以pMD19-T-IFN-α質粒為模板、F2/R2為引物擴增山羊α-干擾素成熟肽編碼序列,擴增產物和pET-32a(+)分別用NcoⅠ、NcoⅠ+HindⅢ限制性內切酶消化,消化產物用10 g/L瓊脂糖電泳分離,經DNA凝膠回收試劑盒純化后的目的基因和載體用T4 DNA連接酶于16 ℃連接過夜,連接產物用于轉化DH5α感受態細胞。重組質粒參照“1.3”中的方法進行PCR和酶切鑒定,重組質粒命名為pET-mIFN-α。
2 結果與分析
2.1 山羊α-干擾素全基因的克隆與序列分析
以pMD19-T-IFN-α為模板、F2/R2為引物擴增獲得大小約為500 bp的特異性擴增片段。重組質粒pET-mIFN-α經NcoⅠ酶切消化得到大小約6 400 bp的DNA片段、經NcoⅠ+HindⅢ酶切消化得到大小約為5 900 bp的載體片段和500 bp的目的片段,與預期結果一致(圖3)。
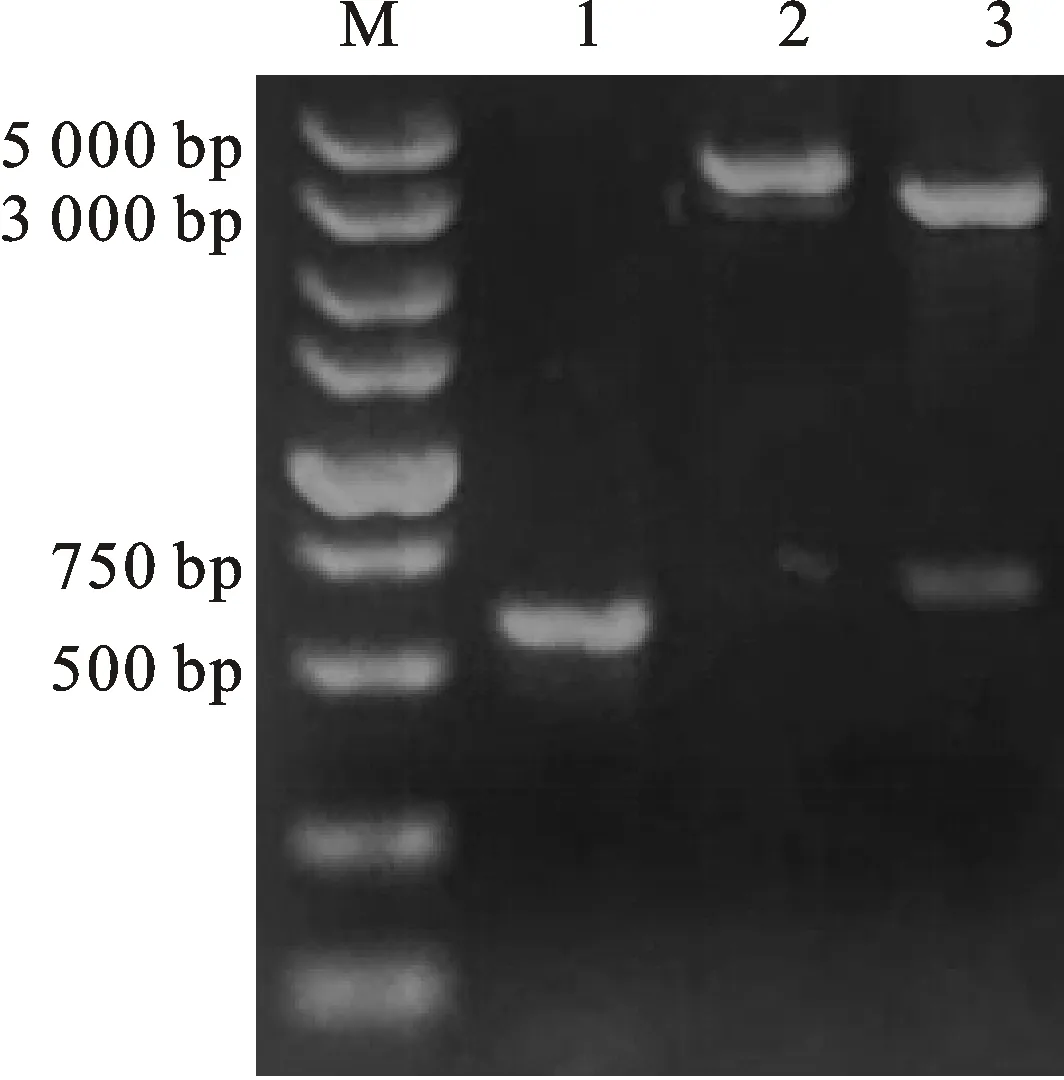
M.DL5000 DNA分子質量標準;1.山羊α-干擾素基因的擴增產物;2.HindⅢ 酶切產物;3.BamH I+HindⅢ酶切產物
測序結果表明,雷州山羊α-干擾素基因全長為570 bp,編碼189個氨基酸。Dnastar(Version 7.1)分析結果顯示,雷州山羊α-干擾素基因與GenBank 上發表的山羊(NM_001285704)、林麝(MW394237)、彎角大羚羊(XM_040238322)、黃牛(NM_001172041)、瘤牛(HM853483)和綿羊(XM_004004404)的核苷酸序列同源性較高,分別為99.8%、95.6%、96.8%、94.7%、94.7%和96.8%,其氨基酸同源性分別為100%、92.6%、94.7%、92.1%、93.2%和93.7%,提示反芻動物α-干擾素基因的核苷酸和氨基酸序列高度保守。
外周血淋巴細胞分離液購自天津灝洋生物制品科技有限責任公司;RPMI 1640培養基購自北京索萊寶科技有限公司;DMEM細胞培養基、MEM細胞培養基、胎牛血清(FBS)購自美國Gibco公司;植物血凝素(PHA)購自上海源葉生物科技有限公司;Trizol RNA提取試劑購自Invitrogen公司;M-MLV反轉錄酶購自 Promega公司;pMD19-T、T4 DNA 連接酶、限制性內切酶BamH Ⅰ、Hind Ⅲ、NcoⅠ,DNA片段凝膠回收試劑盒、質粒DNA抽提試劑盒、DNA分子質量標準均購自寶日醫生物技術(北京)有限公司;TaqDNA聚合酶購自北京鼎國生物公司;PMSF購自北京蘭杰柯科技有限公司;BCA蛋白測定試劑盒購自中國上海博彩生物科技有限公司;HRP標記的馬抗小鼠IgG購自美國Cell Signaling Technology;Ni-NAT親和層析樹脂購自GE Healthcaer公司;10K超濾管購自Millipor公司。
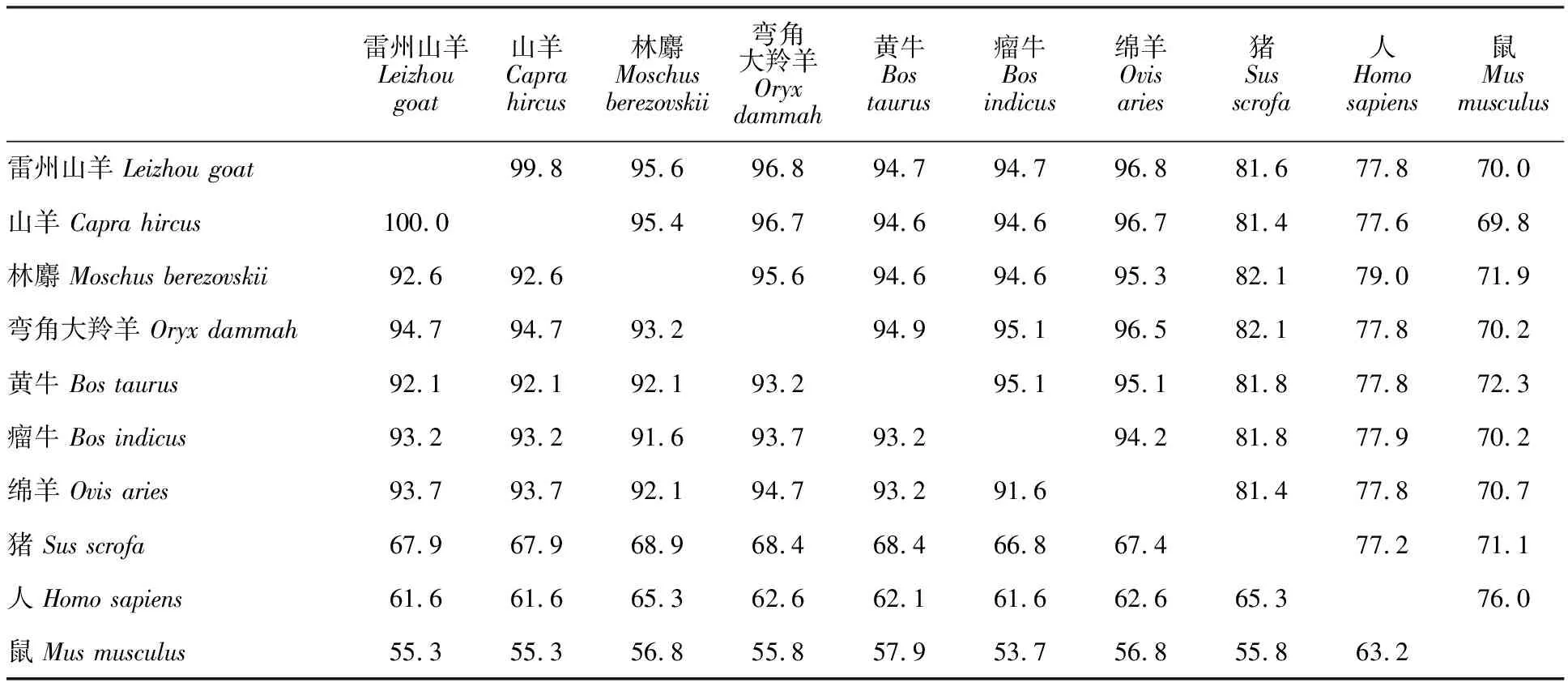
表1 α-干擾素基因核苷酸與氨基酸序列同源性比較
氨基酸序列分析結果表明,雷州山羊α-干擾素信號肽潛在裂解位點位于N末端22~23個氨基酸,其成熟肽由167個氨基酸組成,其中無N-糖基化位點,有4個潛在的O-糖基化位點,分別為第48位(S)、第92位(S)、第140位(S)和第151位(T),4個半胱氨酸殘基分別位于第24、52、122、162位,與其他反芻動物α-干擾素的一級結構相同(圖2)。雷州山羊和其他反芻動物的α-干擾素成熟肽在26~185位氨基酸均含有保守的結構域,雷州山羊、彎角大羚羊和綿羊α-干擾素成熟肽在第48、140、151位均有潛在O-糖基化位點,瘤牛α-干擾素成熟肽在第48和140位也有潛在O-糖基化位點,瘤牛和綿羊α-干擾素成熟肽在第84位均有潛在O-糖基化位點,但均無潛在N-糖基化位點,且與豬、人和鼠的潛在O-糖基化位點截然不同;包括雷州山羊在內的反芻動物與豬、人和鼠α-干擾素成熟肽均含有4個位點相同的半胱氨酸殘基。
2.2 山羊α-干擾素成熟肽擴增及表達載體的構建
通過RT-PCR方法擴增的雷州山羊α-干擾素全基因大小約570 bp,與預期結果一致。重組質粒pMD19-T-IFN-α經HindⅢ 酶切消化得到大小約3 200 bp的DNA片段,經BamH Ⅰ+HindⅢ酶切消化得到大小分別為2 700 bp的載體片段和570 bp的目的片段,與預期結果一致(圖1)。
2.3 重組山羊α-干擾素的表達、純化及鑒定
通過比較不同表達時間、誘導劑濃度、誘導表達溫度對重組α-干擾素成熟肽表達量的影響,確定最佳表達條件為37 ℃、IPTG終濃度0.5 mmol/L、誘導表達6 h,此時重組蛋白表達量最高,且主要以包涵體形式存在,其分子質量約32 ku,與預期結果一致(圖4)。Western-blot 檢測結果顯示,重組菌BL-pET-32a(+)和BL-pET-32a-mIFN-α的誘導表達產物均能與6×His抗體發生特異性結合反應,其分子質量分別為19 ku和32 ku,與預期結果相符(圖5)。
用重組質粒pET-mIFN-α轉化BL21(DE3)感受態細胞,獲得重組菌BL-pET-mIFN-α,于 37 ℃、210 r/min培養至OD600nm約0.6~1.0,加入IPTG使其終濃度為0.5 mmol/L,繼續誘導表達6 h,收集菌體用滅菌三蒸水重懸,反復凍融3 次,加入DNase Ⅰ 置于37 ℃至菌液不再粘稠, 4 ℃、12 000 r/min離心15 min,分別收集上清和沉淀,用12% SDS-PAGE 凝膠電泳檢測;分別于25、30、37 ℃以IPTG終濃度為0.1、0.5、1.0、2.0 mmol/L分別誘導0、1、2、4、6 h,用12% SDS-PAGE 凝膠電泳分析重組蛋白的可溶性,以優化表達條件[10]。
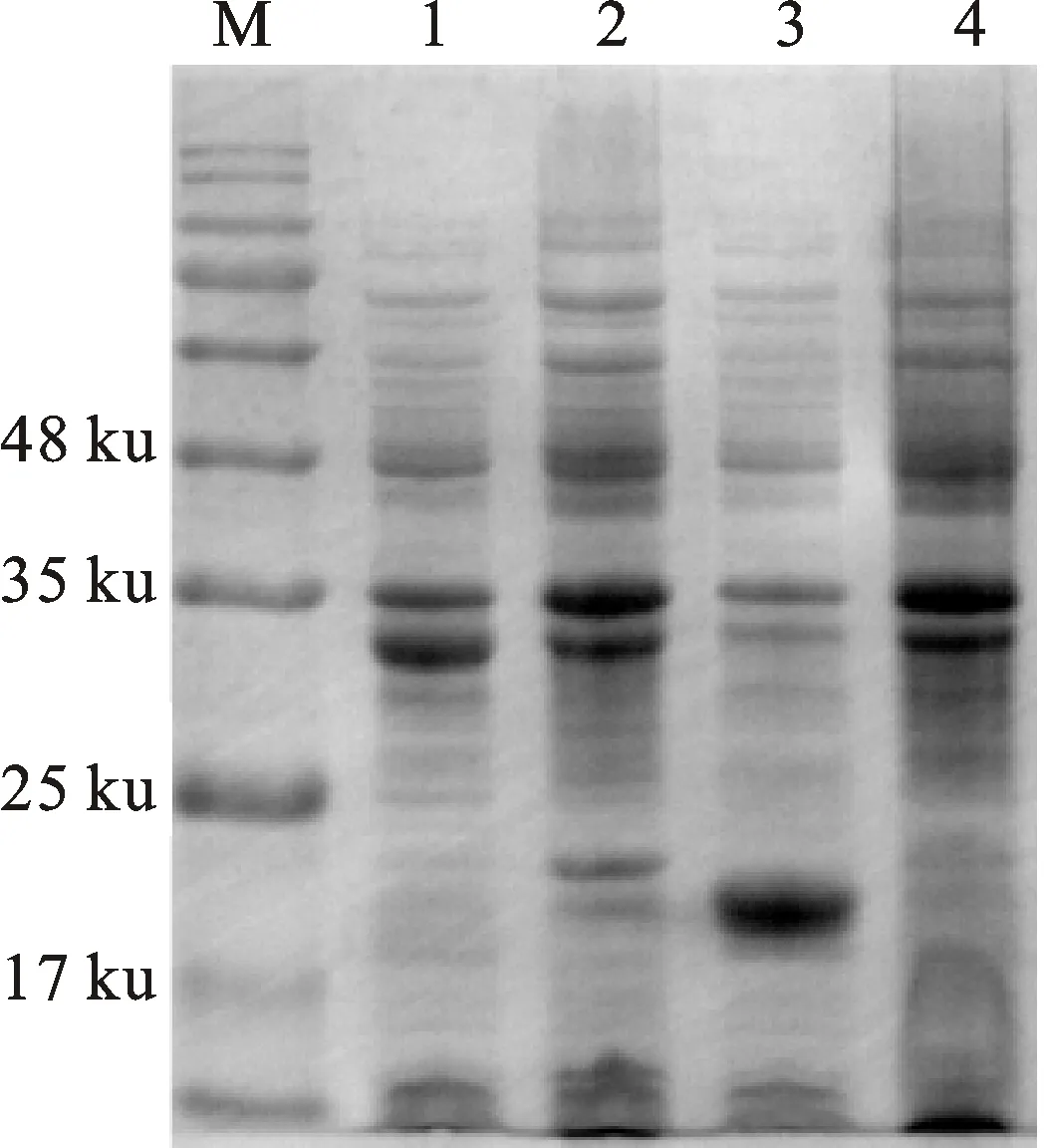
M.蛋白質分子質量標準;1.誘導表達6 h的BL-pET-mIFN-α重組菌裂解產物;2.未誘導的BL-pET-mIFN-α重組菌裂解產物;3.BL-pET-32a(+)重組菌裂解產物;4.BL21(DE3)細菌裂解產物
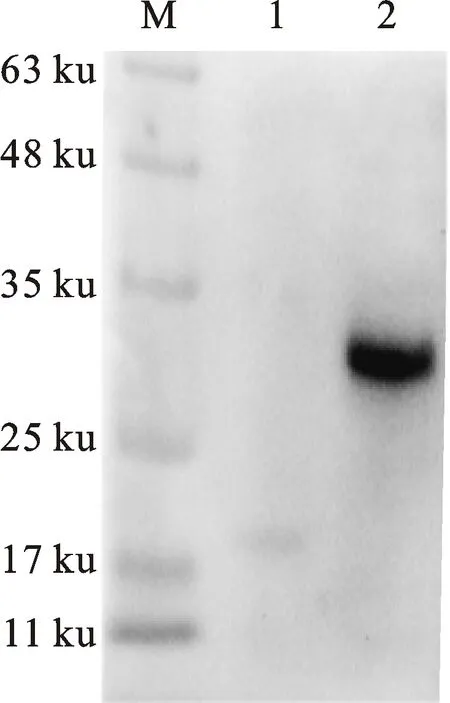
M.蛋白質分子質量標準;1.BL-pET-32a(+)細菌裂解產物;2.純化的rG-mIFN-α
2.4 重組山羊α-干擾素抗病毒活性測定
采用微量細胞病變抑制法檢測重組山羊 α-干擾素的抗PPRV和GPV活性。對照組細胞大小形態正常,貼壁良好;病毒對照組細胞在PPRV或GPV感染后,逐漸出現細胞變圓、融合和壞死脫落現象;經 rG-mIFN-α處理的細胞,僅見少量細胞融合和脫落,且隨著rG-mIFN-α終濃度增加,細胞病變程度越輕微。由此可見,重組山羊α-干擾素成熟肽能以劑量依賴方式顯著抑制PPRV和GPV誘導的細胞病變(圖6)。rG-mIFN-α抗PPRV和GPV活性分別為1.024×104U/mL和8.67×103U/mL,比活力分別為 1.6×104U/mg和1.35×104U/mg。說明重組山羊α-干擾素成熟肽具有較強的抗PPRV和抗GPV活性。
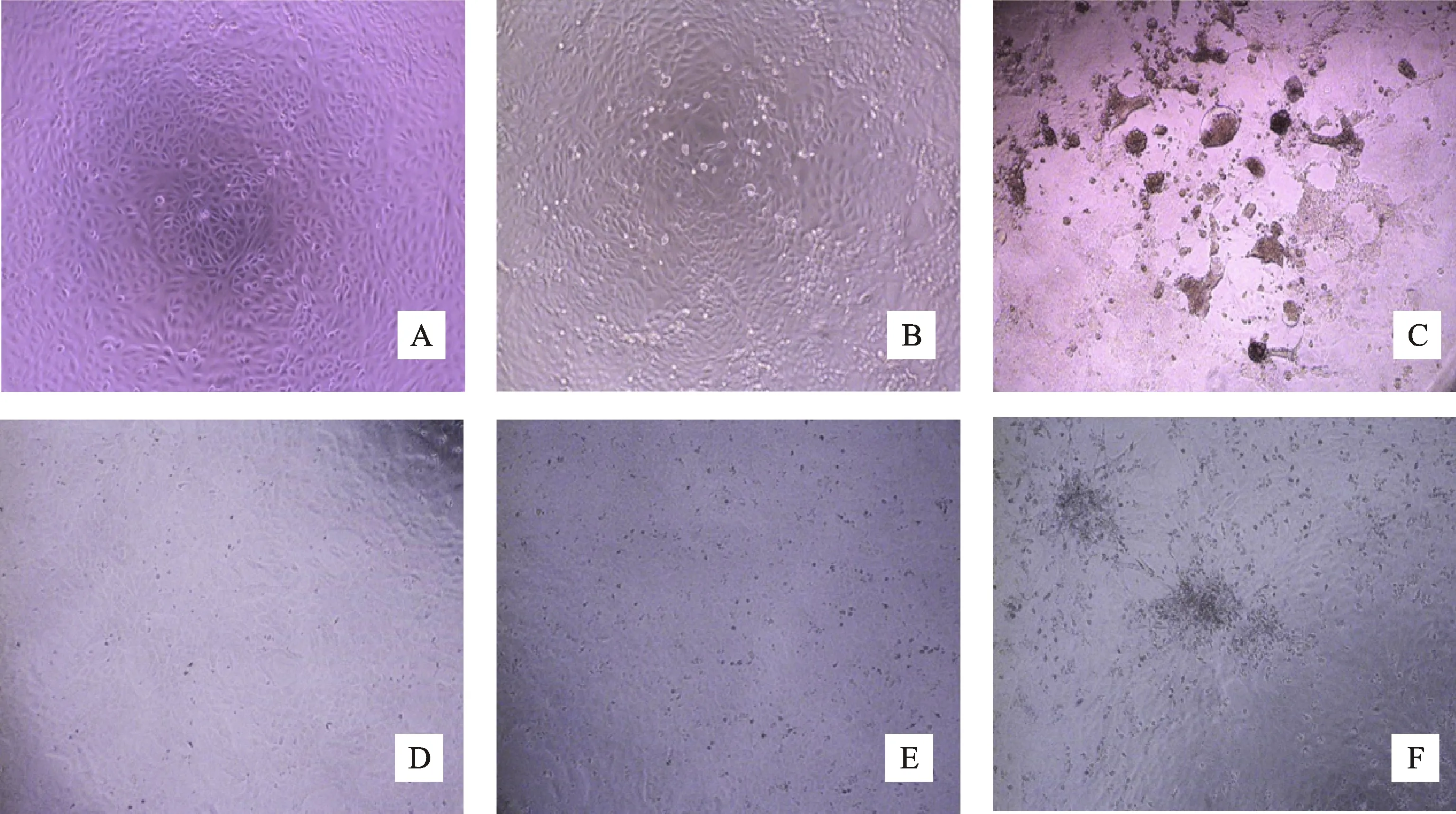
A.Vero細胞對照;B.rG-mIFN-α處理的PPRV感染Vero細胞;C.PPRV感染Vero細胞;D.LDG-2細胞對照;E.rG-mIFN-α處理的GPV感染LDG-2細胞;F.GPV感染LDG-2細胞
3 討 論
本研究證實,雷州山羊IFN-α基因由570個核苷酸組成,編碼189個氨基酸,與Genbank報道的其他山羊品種的α-干擾素核苷酸與氨基酸序列同源性高達99.8%和100%,僅存在1個核苷酸位點的同義突變;與綿羊、彎角大羚羊、林麝、瘤牛等反芻動物的 α-干擾素氨基酸序列都有較高同源性,與豬、人、小鼠的同源性較低,提示反芻動物的α-干擾素氨基酸序列高度保守,且親緣關系較近的動物干擾素同源性也較高。
我恨恨地把窗戶玻璃打破了,爬了出去。月亮很亮,我踩著自己的影子晃悠在扒鍋街,我去找劉佳,扒鍋街的人我最惦記的就是他了,我只親了他一次,遠遠不夠哩。
第三,游客對長江三峽地域文化認知過程具有選擇性。出游前,游客通過信息刺激產生初始認知;出游過程中,游客在個體已有知識和體驗的影響下,產生對信息的選擇性“接觸-注意-理解-保持”[33]86過程;出游后,游客形成的認知印象以概念網絡的形式進行儲存,反映了部分的目的地文化。
IFN-α的N-末端有22個氨基酸構成的信號肽,其切割位點位于第22位(L)和23位(G)之間,氨基酸序列分析結果表明,反芻動物α-干擾素信號肽的胞外區和胞內區均具有共同基序(motif),即MAPAWS和SCNAICS;除彎角大羚羊外,跨膜區則含有數量相同的疏水氨基酸—亮氨酸(L);除綿羊與瘤牛外,大多數反芻動物的α-干擾素信號肽裂解位點序列均為第22位(L)-第23位(G),提示反芻動物α-干擾素的易位、跨膜轉運、信號肽切除及分泌機制基本相同。雷州山羊和其他反芻動物α-干擾素成熟肽在26~185位氨基酸均含有保守的結構域,其中山羊、彎角大羚羊和綿羊α-干擾素成熟肽在第48、140、151位氨基酸均有潛在O-糖基化位點,瘤牛和綿羊α-干擾素成熟肽的糖基化位點則位于第48、84、140氨基酸,且與豬、人和鼠的潛在O-糖基化位點截然不同,目前尚不清楚這些糖基化位點的細微差別對反芻動物α-干擾素高級結構和抗病毒活性的影響。
在這一段簡短的語言描述里,作者綜合運用了多種修辭方法:運用比喻的修辭,在把“廣場”比作“露天公寓”的過程中,形象地再現了蘇比生活的貧窮;運用擬人修辭,通過“打招呼”這一動詞性的短語,把北風預示的寒冷以及這些寒冷給予像蘇比一樣窮人的警告,以一種幽默的方式體現在語言描寫之中;運用借代,作者把像蘇比一樣的廣場流浪漢們,形象地稱之為“房客們”。從而在幽默之中表達著一絲諷刺。
原核表達系統具有操作簡便、重組蛋白表達量高、生產成本低廉等優勢。郝飛等[14]成功克隆山羊IFN-τ基因并進行原核表達,發現重組山羊IFN-τ能有效抑制水泡性口炎病毒及山羊副流感病毒3型在宿主細胞中的增殖。Jacobe等[15]在大腸桿菌中表達了綿羊IFN-γ基因,并測得融合蛋效價為3.2×105U/mg。本研究結果表明,山羊α-干擾素能在大腸桿菌中高效表達,表達量可達1.84×103mg/L,但重組山羊α-干擾素多以包涵體形式存在。通過優化表達條件如調整重組菌密度、培養溫度、誘導劑終濃度等仍未能實現重組山羊α-干擾素的可溶性表達。采用常規方法對包涵體進行變性和復性處理,并通過超濾工藝大大縮短了重組蛋白的濃縮時間,獲得的高純度重組山羊α-干擾素具有較強的抗病毒活性,其抗PPRV和GPV效價分別達到1.6×104U/mg和1.35×104U/mg。值得指出的是,在原核細胞中表達的外源蛋白無法進行糖基化修飾,可能對重組蛋白的折疊、抗原表位形成、電荷性質、熱穩定性及其生物學功能產生顯著影響[16]。李丹等[17]利用家蠶桿狀病毒表達系統表達出羊α干擾素,抗PPRV活性為6.5×105U/mL;于力等[18]利用畢赤酵母系統表達山羊α干擾素,抗病毒活性為5.62×109U/mg。考慮到真核表達系統制備重組蛋白的周期長、成本高、表達量較低等因素,原核表達系統仍是生產重組蛋白的重要手段。未來將通過選擇適宜的分泌型表達載體和分子伴侶以增加重組蛋白的可溶性表達,簡化重組蛋 白的分離純化,進一步提高重組蛋白的生物學活性。
