鹽脅迫與供磷水平對柳枝稷苗期光合特性與總生物量的影響
2023-08-01 11:27:50麻仲花陳娟吳娜滿本菊王曉港者永清劉吉利
中國農業(yè)科技導報 2023年6期
關鍵詞:水平
麻仲花, 陳娟, 吳娜, 滿本菊, 王曉港, 者永清, 劉吉利
(1.寧夏大學農學院,銀川 750021; 2.寧夏大學生態(tài)環(huán)境學院,銀川 750021)
柳枝稷(Panicum virgatumL.)作為禾本科黍屬多年生草本C4植物,適宜在鹽堿地種植[1]。土地鹽漬化是目前農業(yè)生產中普遍存在的問題,嚴重制約植物的生長發(fā)育,影響農作物的產量和品質。磷作為植物必需營養(yǎng)元素之一,對植物光合作用和生長發(fā)育起至關重要的作用。但鹽脅迫抑制植物生長發(fā)育的同時,也顯著降低了土壤中有效磷的利用率,甚至限制根系對磷的吸收利用,影響作物產量。在農業(yè)生產中通常用磷肥來補充土壤中有效磷的含量[2-3]。黃高鑒等[4]研究表明,鹽脅迫顯著抑制柳枝稷的生長發(fā)育,使其生物量顯著降低。常雯雯等[5]研究表明,鹽脅迫條件下柳枝稷的光合特性顯著降低,但不同品種間存在差異。劉金彪等[6]認為,低磷脅迫環(huán)境下柳枝稷為吸收磷素而蒸騰損失水分,減弱光合作用,從而降低生物量。張蕊等[7]認為,在逆境脅迫下增施磷肥可增強植物光合生理特性及植物生長發(fā)育能力,從而緩解逆境對植物造成的傷害,進而提高產量。目前,在鹽脅迫條件下,磷素的流失對柳枝稷光合特性及生物量影響的研究多集中在鹽脅迫或供磷水平單因素上。柳枝稷苗期對鹽和低磷脅迫均比較敏感[6,8],因此,本研究采用水培試驗研究鹽脅迫與供磷水平2種因素對不同品種柳枝稷苗期光合特性及生物量的影響,并進一步揭示苗期柳枝稷光合特性與總生物量的關系,為柳枝稷在鹽堿地的推廣種植提供理論依據(jù)。
1 材料與方法
1.1 試驗材料
試驗在北京市農林科學院溫室大棚內開展,以3個不同品種柳枝稷(八倍體Alamo、Alamo和Pathfinder)為試驗材料,品種特性詳見表1。
1.2 試驗設計
采用水培試驗,設置鹽脅迫和供磷水平2個因素,其中鹽脅迫分別設置0(A1)和100 mmol·L-1NaCl(A2)共2個水平;磷肥施用量分別設置0(B1)、5(B2)、200(B3)和2 000 mmol·L-1KH2PO4(B4)4個水平,共計8個處理,分別為A1B1、A1B2、A1B3、A1B4、A2B1、A2B2、A2B3、A2B4,每個處理3次重復。
挑選大小一致飽滿的柳枝稷種子,通過沙培在光照培養(yǎng)箱中育苗(培養(yǎng)條件為:晝/夜室溫35 ℃/20 ℃,光周期14 h/10 h,光照強度400 μM·m-2·s-1),待幼苗長到3片展開葉,挑選生長一致的幼苗移栽至水培箱(箱體長41 cm,寬30.5 cm,高13.5 cm)中培養(yǎng)。每個水培箱內,幼苗基部用海綿包裹,放入聚乙烯泡沫板預先打好的孔中,然后固定于水培箱中。水培箱體用錫箔紙包裹,以防止光照,并避免藻類的繁殖。每個水培箱灌注營養(yǎng)液13.9 L,用氣泵進行24 h充氣,營養(yǎng)液每3 d更換1次。試驗營養(yǎng)液配比采用不同磷水平的Hoagland營養(yǎng)液(配方詳見表2)。KH2PO4為營養(yǎng)液中的磷素來源,為保持鉀素的平衡,改變KH2PO4的含量時需調節(jié)相應KCl含量。柳枝稷材料先在正常條件下培養(yǎng)至3葉1心,然后進行不同鹽脅迫和供磷水平處理,處理第7天進行各項指標測定。每個處理種植6盆,每盆種植16株。

表2 不同磷水平的Hoagland營養(yǎng)液配置Table 2 Hoagland nutrient solution configuration table with different phosphorus levels
1.3 測定指標及方法
(1)葉綠素含量采用乙醇浸提法進行測定:稱取葉片 0.05~0.08 g,記下具體質量,剪碎后置于離心管中;加入8 mL 95%的乙醇,避光靜置24 h;分別于665、649 nm 下測定其吸光度值,并計算葉綠素含量。
(2)凈光合速率(net photosynthetic rate,Pn)、氣孔導度(stomatal conductance,Gs)、蒸騰速率(transpiration rate,T)r、胞間CO2濃度(intercellular CO2concentration,C)i采用LI-6400光合儀進行測定。具體方法為:于晴朗天氣9∶00—10∶00,選取其各株從上往下數(shù)第1片葉進行測定,每個處理隨機測量5株。
(3)生物量測定:將整株取出,測定單株地上部分生物量鮮重;將待測植株沖洗干凈,置于105 ℃鼓風干燥箱中殺青30 min,然后在65 ℃下烘干至恒重,測定單株地上部分生物量干重。
1.4 數(shù)據(jù)處理
采用Excel 2016、SPSS 25.0和0rgin 2020統(tǒng)計軟件進行方差分析、相關性分析、模糊數(shù)學隸屬函數(shù)分析。隸屬函數(shù)的計算方法[9]如下。
式中,Xi為i指標測定值,Ximin和Ximax分別為i指標的最小值和最大值。
2 結果分析
2.1 鹽脅迫與供磷水平對苗期柳枝稷光合特性及總生物量的影響
2.1.1 鹽脅迫與供磷水平對葉綠素含量的影響由圖1可知,3個柳枝稷品種的葉片葉綠素含量在同一鹽處理下均隨著供磷水平的增加呈上升趨勢。在非鹽脅迫下,八倍體Alamo、Alamo和Pathfinder的葉綠素含量隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的葉綠素含量較無磷處理分別提高17.45%、95.16%、86.87%。在鹽脅迫條件下,八倍體Alamo、Alamo和Pathfinder葉綠素含量隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的葉綠素含量較無磷處理分別提高115.45%、188.33%、108.20%。
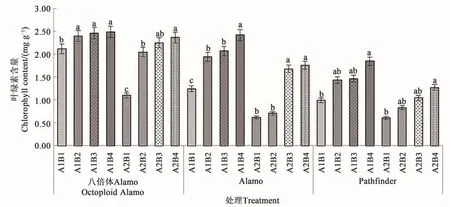
圖1 不同鹽脅迫與供磷水平下3個柳枝稷品種的葉綠素含量Fig.1 Chlorophyll content of 3 switchgrass varieties under different salt stress and phosphorus supply
由表3可知,鹽脅迫顯著降低了Alamo和Pathfinder葉綠素含量,但對八倍體Alamo影響不顯著;供磷顯著增加了Pathfinder葉綠素含量,但對八倍體Alamo和Alamo影響不顯著;兩因素間的交互作用對八倍體Alamo和Alamo的葉綠素含量影響顯著,但對Pathfinder影響不顯著。
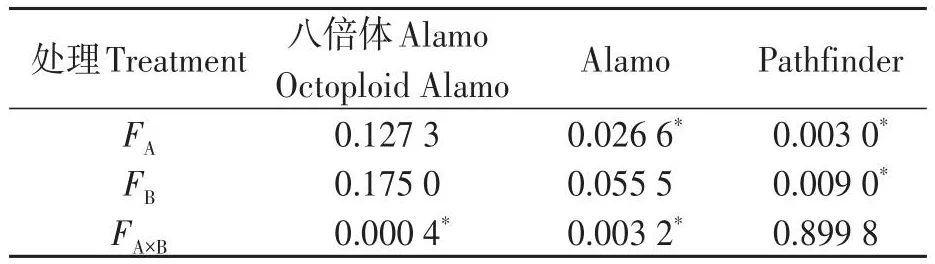
表3 葉綠素含量方差分析Table 3 Analysis of variance of chlorophyll content
2.1.2 鹽脅迫與供磷水平對凈光合速率的影響由圖2可知,3個柳枝稷品種的葉片凈光合速率在同一鹽處理下均隨著供磷水平的增加呈上升趨勢。在非鹽脅迫下,八倍體Alamo、Alamo和Pathfinder凈光合速率隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的凈光合速率較無磷處理分別提高21.84%、15.89%、18.33%。在鹽脅迫條件下,八倍體Alamo、Alamo和Pathfinder凈光合速率隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的凈光合速率較無磷處理分別提高14.63%、15.28%、23.35%。
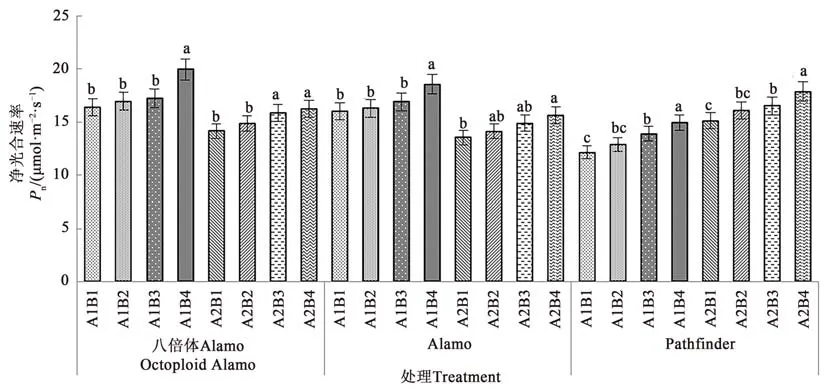
圖2 不同鹽脅迫與供磷水平下3個柳枝稷品種的凈光合速率Fig.2 Pn of 3 switchgrass varieties under different salt stress and phosphorus supply levels
由表4可知,鹽脅迫顯著降低了八倍體Alamo、Alamo和Pathfinder的凈光合速率;供磷顯著增加了Alamo和Pathfinder的凈光合速率,但對八倍體Alamo凈光合速率影響不顯著;兩因素的交互作用對八倍體Alamo、Alamo和Pathfinder的凈光合速率影響均不顯著。
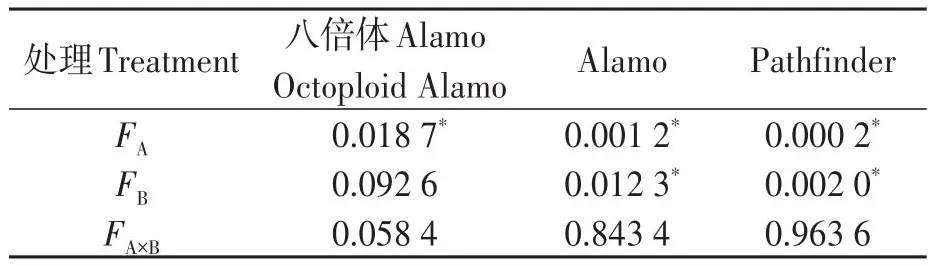
表4 凈光合速率方差分析Table 4 Analysis of variance of Pn
2.1.3 鹽脅迫與供磷水平對氣孔導度的影響 由圖3可知,3個柳枝稷品種苗期葉片氣孔導度在同一鹽處理下均隨著供磷水平的增加呈上升趨勢。在非鹽脅迫下,八倍體Alamo、Alamo和Pathfinder氣孔導度隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的氣孔導度較無磷處理分別提高42.86%、28.57%、75.00%。在鹽脅迫條件下,八倍體Alamo、Alamo和Pathfinder氣孔導度隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的氣孔導度較無磷處理分別提高16.67%、100.00%、100.00%。
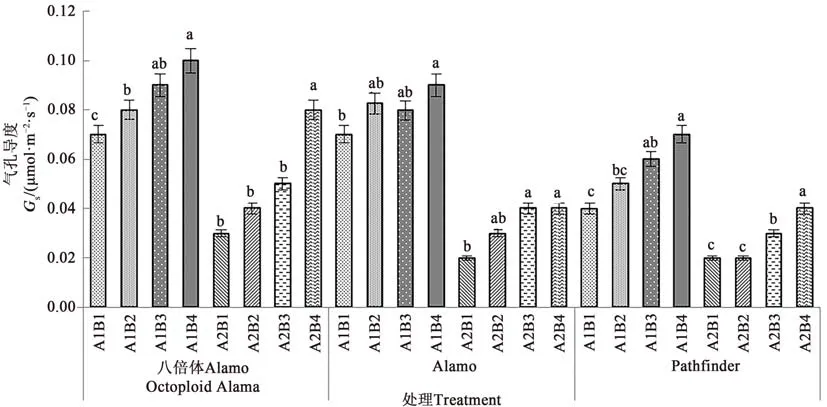
圖3 不同鹽脅迫與供磷水平下3個柳枝稷品種的氣孔導度Fig.3 Gs of 3 switchgrass varieties under different salt stress and phosphorus supply levels
由表5可知,鹽脅迫顯著降低了八倍體Alamo、Alamo和Pathfinder苗期葉片氣孔導度;供磷顯著增加了Pathfinder氣孔導度,但對八倍體Alamo和Alamo的影響不顯著;兩因素的交互作用對八倍體Alamo和Pathfinder影響均不顯著,但對Alamo影響顯著。
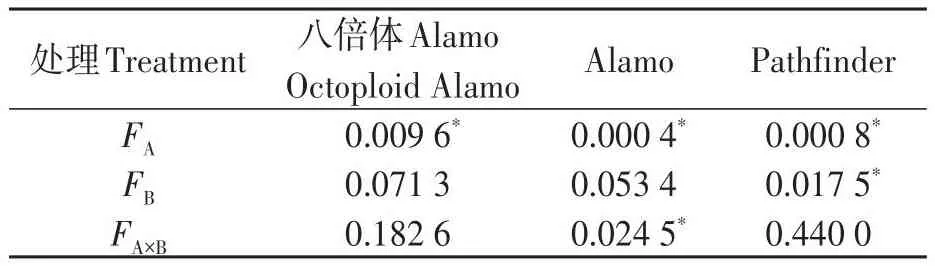
表5 氣孔導度方差分析Table 5 Analysis of variance of Gs
2.1.4 鹽脅迫與供磷水平對胞間CO2濃度的影響 由圖4可知,3個柳枝稷品種苗期葉片胞間CO2濃度在同一鹽處理下均隨著供磷水平的增加呈下降趨勢。在非鹽脅迫下,八倍體Alamo、Alamo和Pathfinder胞間CO2濃度隨著磷水平增加均呈現(xiàn)下降趨勢,其中全磷處理的胞間CO2濃度較無磷處理分別降低50.12%、68.07%、63.11%。在鹽脅迫條件下,八倍體Alamo、Alamo和Pathfinder胞間CO2濃度隨著磷水平增加均呈現(xiàn)下降趨勢,其中全磷處理的胞間CO2濃度較無磷處理分別降低49.68%、40.41%、40.21%。
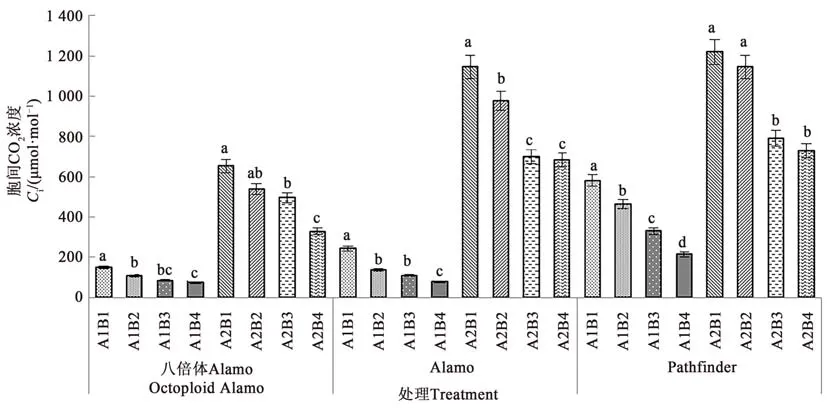
圖4 不同鹽脅迫與供磷水平下3個柳枝稷品種的胞間CO2濃度Fig.4 Ci of 3 switchgrass varieties under different salt stress and phosphorus supply level
由表6可知,鹽脅迫增加了八倍體Alamo、Alamo和 Pathfinder胞間CO2濃度;供磷顯著降低了Pathfinder胞間CO2濃度,但對八倍體Alamo和Alamo影響不顯著;兩因素的交互作用對八倍體Alamo、Alamo和 Pathfinder影響顯著。
2.1.5 鹽脅迫與供磷水平對蒸騰速率的影響 由圖5可知,3個柳枝稷品種蒸騰速率在同一鹽處理下均隨著供磷水平的增加呈上升趨勢。在非鹽脅迫下,八倍體Alamo、Alamo和Pathfinder蒸騰速率隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的蒸騰速率較無磷處理分別提高21.98%、37.62%、42.60%。在鹽脅迫條件下,八倍體Alamo、Alamo和Pathfinder蒸騰速率隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的蒸騰速率較無磷處理分別提高71.10%、37.20%、50.35%。
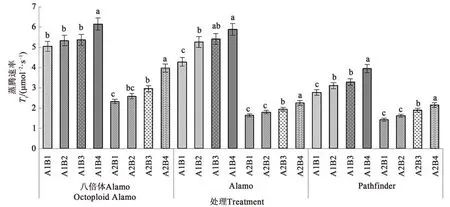
圖5 不同鹽脅迫與供磷水平下3個柳枝稷品種的蒸騰速率Fig.5 Tr of 3 switchgrass varieties under different salt stress and phosphorus supply level
由表7可知,鹽脅迫顯著降低了八倍體Alamo、Alamo和Pathfinder蒸騰速率;供磷對八倍體Alamo和Pathfinder蒸騰速率影響顯著,但對Alamo影響不顯著;兩因素的交互作用對Alamo和Pathfinder蒸騰速率影響顯著,但對八倍體Alamo影響不顯著。
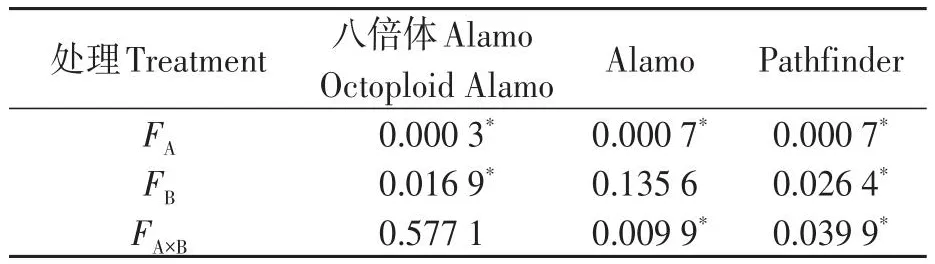
表7 蒸騰速率方差分析Table 7 Analysis of variance of Tr
2.1.6 鹽脅迫與供磷水平對總生物量的影響 由圖6可知,3個柳枝稷品種總生物量在同一鹽處理下均隨著供磷水平的增加呈上升趨勢。在非鹽脅迫下,八倍體Alamo、Alamo和Pathfinder總生物量隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的總生物量較無磷處理分別提高246.52%、115.32%、145.85%。在鹽脅迫條件下,八倍體Alamo、Alamo和Pathfinder總生物量隨著磷水平增加均呈現(xiàn)上升的趨勢,其中全磷處理的總生物量較無磷處理分別提高91.00%、113.97%、405.88%。
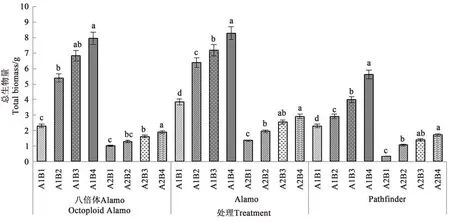
圖6 鹽脅迫與供磷水平下3個柳枝稷品種的總生物量Fig.6 Biomass of 3 switchgrass varieties under different salt stress and phosphorus supply levels
由表8可知,鹽脅迫顯著降低了八倍體Alamo、Alamo和Pathfinder總生物量;供磷對八倍體Alamo、Alamo和Pathfinder總生物量均無顯著影響;兩因素的交互作用對八倍體Alamo、Alamo和Pathfinder總生物量有顯現(xiàn)影響。
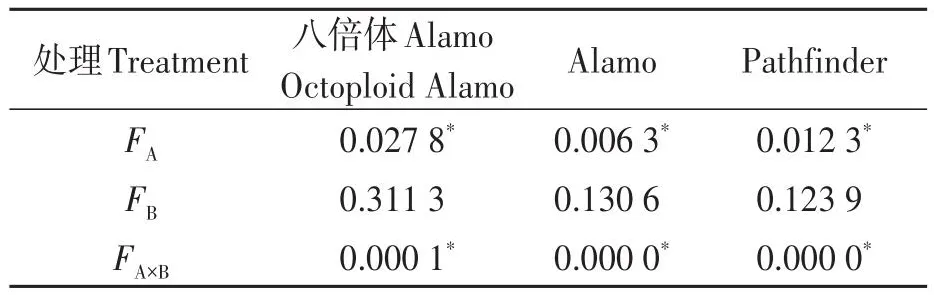
表8 總生物量方差分析Table 8 Analysis of variance of total biomass
2.2 不同柳枝稷品種苗期總生物量與光合特性的相關性分析
分析不同柳枝稷品種苗期總生物與光合特性之間的關系,結果(表9)表明,八倍體Alamo、Alamo和Pathfinder的總生物量與葉綠素含量、蒸騰速率、氣孔導度、凈光合速率均呈正相關關系,與胞間CO2濃度呈負相關關系。其中,八倍體Alamo總生物量與蒸騰速率、氣孔導度、凈光合速率呈顯著正相關,相關系數(shù)分別為0.88、0.85、0.89,與胞間CO2濃度呈顯著負相關,相關系數(shù)為0.83;Alamo總生物量與葉綠素含量呈顯著正相關,相關系數(shù)為0.86,與胞間CO2濃度呈顯著負相關,相關系數(shù)為0.92。

表9 3個柳枝稷品種苗期總生物量與光合特性的相關性分析Table 9 Correlation analysis of total biomass and photosynthetic characteristics of 3 switchgrass varieties at different seedling stage
2.3 不同柳枝稷品種苗期光合特性及總生物量的隸屬函數(shù)分析
不同鹽脅迫與供磷水平下各光合指標及總生物量隸屬函數(shù)值的平均值如表10所示。3個柳枝稷品種各光合指標及總生物量的隸屬函數(shù)值平均值表現(xiàn)為八倍體Alamo>Alamo>Pathfinder。其中,八倍體Alamo的光合指標及總生物量隸屬函數(shù)值的綜合平均值最高,為0.50,最適宜在鹽脅迫與低磷脅迫條件下種植。

表10 3個柳枝稷品種苗期光合特性及總生物量的隸屬函數(shù)值Table 10 Membership function values of photosynthetic characteristics and total biomass of 3 switchgrass varieties at seedling stage
3 討論
植物光合作用對光能的利用是從對光合色素吸收開始的[10],葉綠素是葉片吸收、傳遞和轉化光能的基礎[11-12]。葉綠素含量可以直接反映植物光合能力的強弱,也是衡量植物抗逆能力的重要指標之一[13-14]。研究發(fā)現(xiàn),鹽脅迫下植物葉片的葉綠素含量顯著降低[15]。本研究結果表明,鹽脅迫下柳枝稷苗期葉片葉綠素含量低于非鹽脅迫處理,這可能是葉綠體色素合成酶活性降低、葉綠素分解加速、葉綠體功能紊亂或結構受損等原因造成[16]。在相同鹽處理下,3個柳枝稷品種苗期葉片的葉綠素含量均隨著供磷水平的提高而增加。這說明適量供磷可以提高植物葉綠素含量,增強光合吸收能力[17],進而提高苗期柳枝稷的光合作用。
植物通過光合作用積累能量與貯藏營養(yǎng)物質,光照強弱直接影響植物生長發(fā)育[18]。氣孔導度是反映光、水、氣交換能力的重要生理指標,其直接影響著植物的光合速率和蒸騰速率[19-22]。研究表明,葉片的胞間CO2濃度越小,氣孔內外CO2濃度差值越大,氣孔能吸收的CO2越多,光合速率就越高;光合速率與氣孔導度、蒸騰速率呈顯著正相關,與胞間CO2濃度呈顯著負相關[23]。本研究結果表明,八倍體Alamo和Alamo的凈光合速率、氣孔導度和蒸騰速率三者呈正相關關系,而與胞間CO2濃度呈負相關關系。研究表明,凈光合速率和氣孔導度升高時,胞間CO2濃度隨之降低,這是由非氣孔因素引起的光合作用變化[24];鹽脅迫對植物幼苗的凈光合速率、氣孔導度、胞間CO2濃度和蒸騰速率有抑制作用[25];補施磷肥可以提高柳枝稷光合特性[6]。本研究表明,八倍體Alamo、Alamo和 Pathfinder在鹽脅迫下的凈光合速率、氣孔導度、蒸騰速率均低于非鹽脅迫處理,而胞間CO2濃度高于非鹽脅迫處理;但相同鹽處理下,隨著供磷水平的增加凈光合速率、氣孔導度、蒸騰速率隨之增大,胞間CO2濃度隨之減小。在同一鹽處理下,全磷處理下的凈光合速率、氣孔導度、蒸騰速率最高,胞間CO2濃度最低,這表明相同鹽處理下供磷越多柳枝稷品種苗期葉片的光合特性越強,與姜宗慶等[26]研究結果一致。
黃高鑒等[4]研究表明,鹽脅迫條件下柳枝稷的生長發(fā)育明顯被抑制,生物量顯著降低。本研究表明,3個柳枝稷品種在鹽脅迫下的生物量低于非鹽脅迫,與前人研究結果一致,這是由于鹽脅迫下葉綠素含量明顯降低,影響了光合作用,從而使生物量顯著降低[27]。陳永亮等[28]研究表明,缺磷時植物幼苗生長明顯受限,生物量累積速率降低。本研究表明,3個柳枝稷品種在相同鹽處理下其生物量均隨著供磷水平的增加而增加,均在全磷處理下達到最大值。鹽脅迫下適當施用磷肥可以補充土壤中的有效磷含量,有利于提高植物生物量[7]。本研究表明,光合作用也是決定柳枝稷品種生物量的關鍵因素之一,其中八倍體Alamo總生物量的增加還與蒸騰速率、氣孔導度、凈光合速率呈正相關,與胞間CO2濃度呈負相關;Alamo總生物量與葉綠素含量呈正相關,與胞間CO2濃度呈負相關,與常雯雯等[5]研究結果一致。3個柳枝稷品種中,八倍體Alamo的光合指標及總生物量隸屬函數(shù)值的平均值最高,表明其適應性最強,最宜在鹽脅迫與低磷地區(qū)推廣種植。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
