我國松毛蟲發生率的空間分布格局及其驅動因子
2023-08-05 19:53:05李晨陽蒙鳳群
植物保護 2023年4期
李晨陽 蒙鳳群
關鍵詞 松毛蟲;空間分布;氣候因子;一般線性模型;最暖季降水量
中圖分類號:S 763 文獻標識碼:A DOI:10.16688/j.zwbh.2022353
松毛蟲是鱗翅目Lepidoptera枯葉蛾科Lasio-campidae松毛蟲屬Dendrolimus昆蟲的統稱,是針葉樹的主要食葉害蟲[1]。松毛蟲是世界性森林害蟲,在全球氣候變暖的背景下,松毛蟲的暴發頻率和發生范圍不斷增加[2-3],其中歐洲和亞洲東部等地區發生較重[4-7],松毛蟲亦為我國主要林業害蟲,全世界已知30余種,我國分布有27種,其中經常暴發成災的有6種,即馬尾松毛蟲Dendrolimus punctatus(Walker)、落葉松毛蟲D.superans(Butler)、油松毛蟲D.tabulae formis Tsai et Liu、赤松毛蟲D.spectabilis (Butler)、云南松毛蟲D.houi Lajonqui-ere和思茅松毛蟲D. kikuchii Matsumura[8]。在我國,松毛蟲平均每年為害的森林面積超200萬hm,造成立木生產量損失約180萬~360萬m,嚴重威脅森林生態安全及林業經濟的發展[9-10]。掌握松毛蟲發生的空間分布格局及其驅動因子,對于松毛蟲發生的監測和防控從而減少其發生至關重要。
目前,國外對不同尺度上松毛蟲發生的空間分布格局及其驅動因子已有大量研究,但均為單一物種的研究[6,11-13]。國內的相關研究亦主要集中于單一物種,不同尺度上的研究則多集中在松毛蟲暴發的局部地區,全國尺度上的研究極少[10, 14-17],僅王慶等[14]對2002年- 2012年期間我國各地區馬尾松毛蟲發生率與氣候因子的關系進行了研究。然而,在全國尺度上,關于松毛蟲屬整個類群發生的空間分布格局及其驅動因子的研究目前并沒有報道,其空間分布格局及其驅動因子是否與單一物種一致還不得而知。雖然影響松毛蟲發生的因素很多(例如地形、林分、氣候和天敵等),但是在大尺度范圍內,氣候因子是影響松毛蟲發生的主要驅動因子[2,6,12-13,18],尤以溫度和降水對松毛蟲發生的影響最大[3,19-20]。一方面,溫度和降水可直接影響松毛蟲的生長發育、繁殖和存活等,從而影響松毛蟲的發生[21-24];另一方面,溫度和降水可以通過影響松毛蟲寄主植物而間接影響松毛蟲的發生[9,25]。我國幅員遼闊,南北相距約5500km,跨緯度約50°,東西相距約5200km,跨經度超過60°,幾乎包括北半球所有的氣候類型[26],我國氣候因子的空間異質性可能是導致松毛蟲發生的空間異質性的因素之一。然而,在全國尺度上,關于氣候因子如何影響松毛蟲屬類群的發生進而影響其空間分布格局的研究并沒有報道。
本研究基于我國各省級行政區2004年—2012年間每年松毛蟲發生面積及其寄主植物面積,確定我國松毛蟲發生率的空間分布格局;同時,確定氣候因子對松毛蟲發生率的影響,旨在為我國各行政區精準監測和防控松毛蟲的發生提供科學依據,降低我國松毛蟲發生所造成的損失。
1材料與方法
1.1數據獲取
1.1.1松毛蟲發生面積及其寄主植物面積
首先,從中國林業統計年鑒獲取2004年—2012年(僅公布了這期間的數據)我國各省級行政區每年松毛蟲發生面積。其次,通過《松毛蟲監測預報技術規程》(LY/T 3030—2018)[27]確定松毛蟲的寄主植物,包括馬尾松、黑松、濕地松、火炬松、落葉松、紅松、油松、樟子松、云杉、冷杉、華山松、赤松、思茅松、云南松、高山松、柏木、柳杉和黃山松等。同時,從植物科學數據中心獲取各寄主植物在我國的分布范圍,精確到地級行政區水平。最后,從林業專業知識服務系統獲取2004年—2012年我國各省級行政區每年松毛蟲各寄主植物的面積,各寄主植物面積的總和即為松毛蟲寄主植物面積。基于以上數據,計算我國各省級行政區每年松毛蟲發生率,即各省級行政區每年松毛蟲發生面積除以各省級行政區每年松毛蟲寄主植物面積。將2004年—2012年期間各省級行政區每年松毛蟲發生率進行平均即為各省級行政區松毛蟲平均發生率,后續所有分析將基于松毛蟲平均發生率。由于缺少港澳臺地區松毛蟲的相關統計數據,因此本研究不包括港澳臺地區松毛蟲的發生情況。
1.1.2氣候因子
從美國國家海洋和大氣管理局(National Cen-ters for Environmental Information)獲取2004年一2012年期間松毛蟲寄主植物分布范圍內各地級行政區內所有氣象站(各行政區氣象站的數量與行政區的面積大小正相關)的日均溫和日降水量,計算各省級行政區的19個生物氣候因子。具體參考O’Donnell等[28]的計算公式,首先,對每個生物氣候因子,計算各氣象站每年相關因子的數值,然后將各氣象站每年數值進行平均得到各氣象站2004年—2012年期間的生物氣候因子;最后,各省級行政區數值以該行政區內所有寄主植物分布的地級行政區氣象站的平均值表示。
1.2數據分析
1.2.1多重共線性檢驗
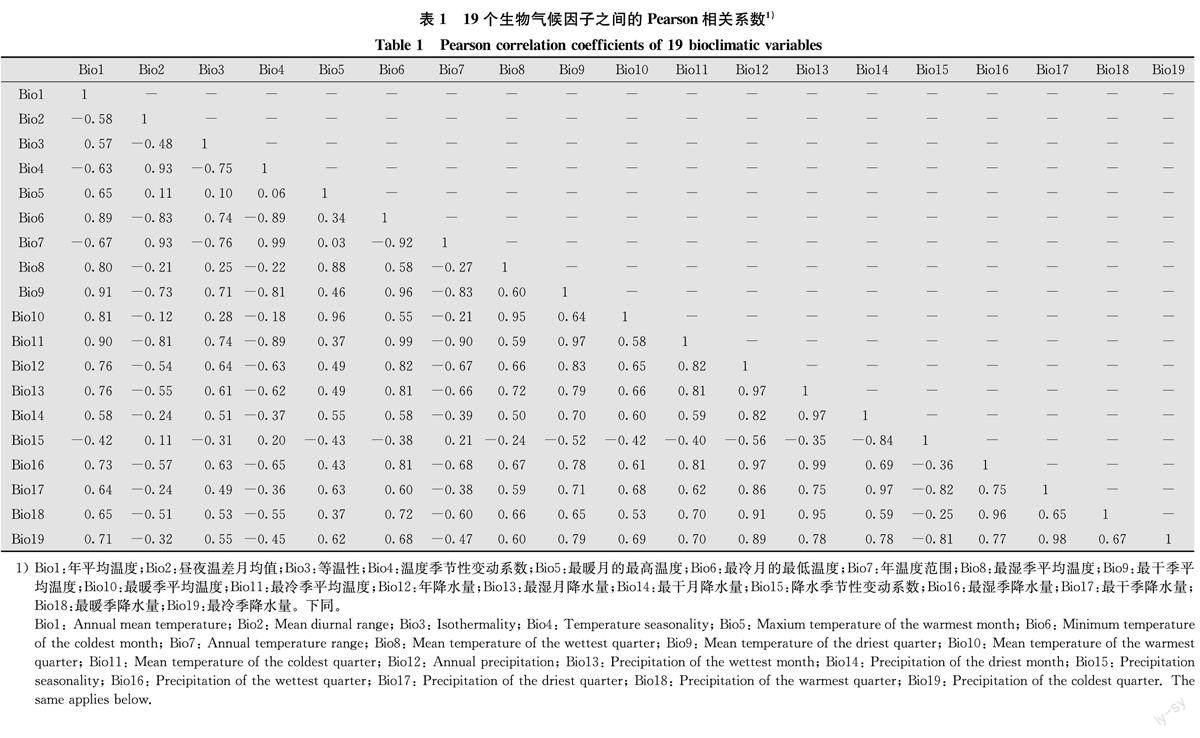
由于各生物氣候因子之間存在多重共線性,為了提高模型精度,本研究參考Xu等[29]的方法對19個生物氣候因子(表1)進行多重共線性檢驗,刪除多重共線性較強的因子。具體地,基于2004年—2012年期間各省級行政區平均生物氣候因子和相應的松毛蟲平均發生率,通過Pearson相關系數和方差膨脹因子(variance inflation factor,VIF)檢驗生物氣候因子之間的共線性。首先,計算任意兩個因子之間的Pearson相關系數,當Pearson相關系數的絕對值≥0.7時被視為高度相關[29]。同時,將19個生物氣候因子與松毛蟲平均發生率進行線性回歸分析,以確定它們之間的相關性。在高度相關的因子中,選擇線性回歸分析中相關性更高(即R更大)的因子。最后,利用VIF將通過Pearson相關系數篩選出的因子進一步進行共線性檢驗,當某一因子的VIF≥5時,表明該因子與篩選出的其他因子共線性強,將該因子從模型中刪除[29]。
1.2.2一般線性模型
基于2004年—2012年各省級行政區平均生物氣候因子和相應的松毛蟲平均發生率,采用一般線性模型檢驗松毛蟲發生率與篩選出的各生物氣候因子之間的關系,其基本形式如下:
2結果與分析
2.1我國松毛蟲發生率的空間分布格局
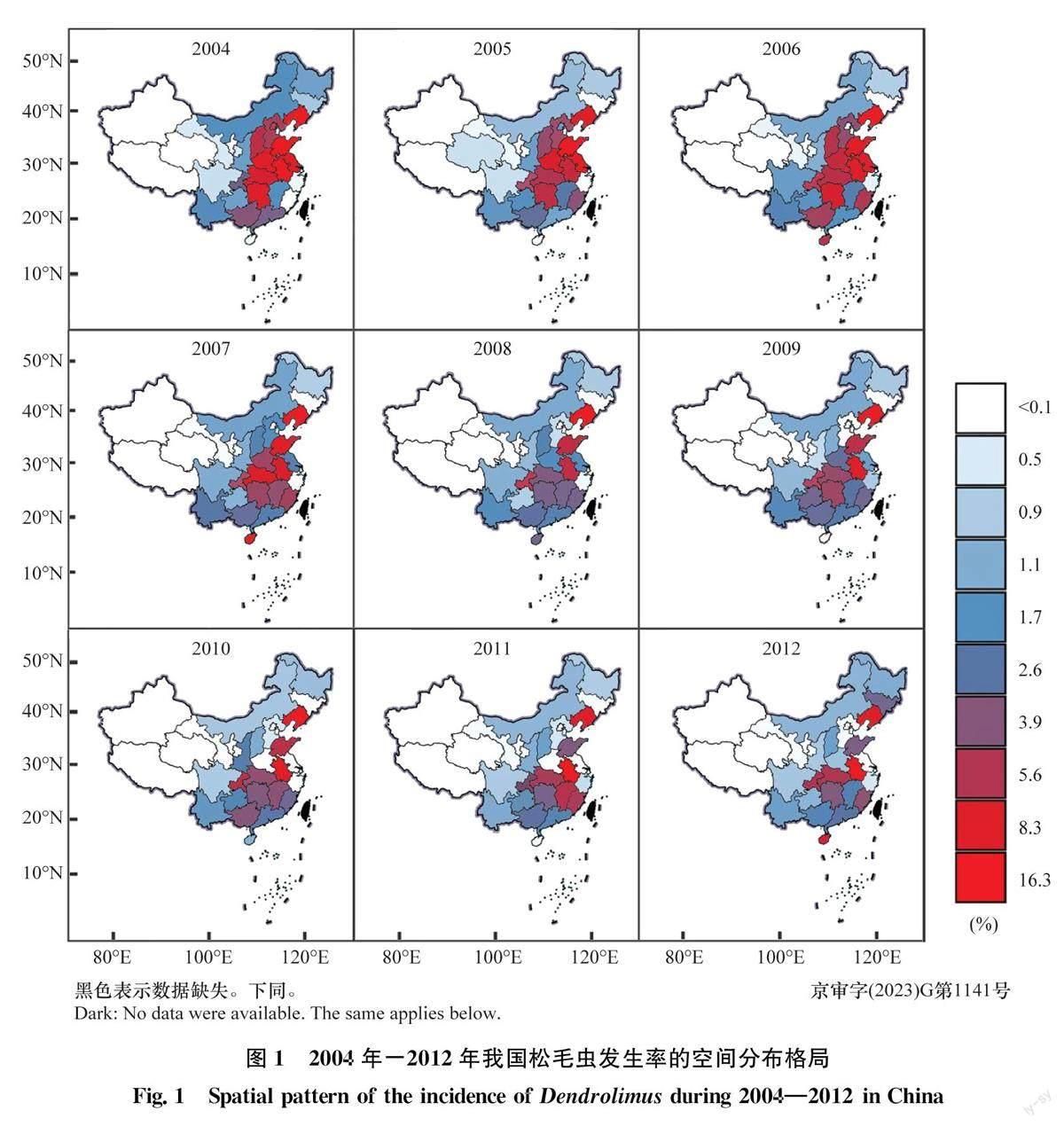
2004年- 2012年全國平均每年松毛蟲發生面積約100萬h1_112,以輕度為害為主,占松毛蟲發生面積的61%,其次為中度為害,占松毛蟲發生面積的26%,而重度為害最少,僅占松毛蟲發生面積的13%。
2004年—2012年,松毛蟲在我國的發生呈現明顯的空間異質性。根據地理區域[31],將我國劃分為北方(包括黑龍江、吉林和遼寧東北3省,以及河北、山西、山東、陜西、河南、北京和天津等黃河中下游5省2市)、南方(包括湖南、湖北、江西、安徽、江蘇、浙江和上海等長江中下游6省1市,以及廣西、廣東、福建、海南、云南、貴州、重慶和四川東南地區)、西北(包括內蒙古、新疆、寧夏和甘肅)和青藏地區(包括青海、西藏以及四川西北地區),不同區域松毛蟲發生率不同。其中,南方和北方部分地區(遼寧、山東和河南等)松毛蟲發生率較高,而西北和青藏地區松毛蟲發生率相對較低(圖1)。例如,雖然松毛蟲寄主植物在全國各省級行政區均有分布,但是2004年—2012年青海、西藏和寧夏3個地區均未見松毛蟲發生,新疆也僅在2011年有發生且發生率極低,為0.01%。2004年- 2012年,我國松毛蟲發生率整體呈現下降趨勢,但總體空間分布格局年際間沒有明顯的變化(圖1)。
2.2我國松毛蟲發生率空間分布格局的主要驅動因子
2.2.1多重共線性檢驗結果
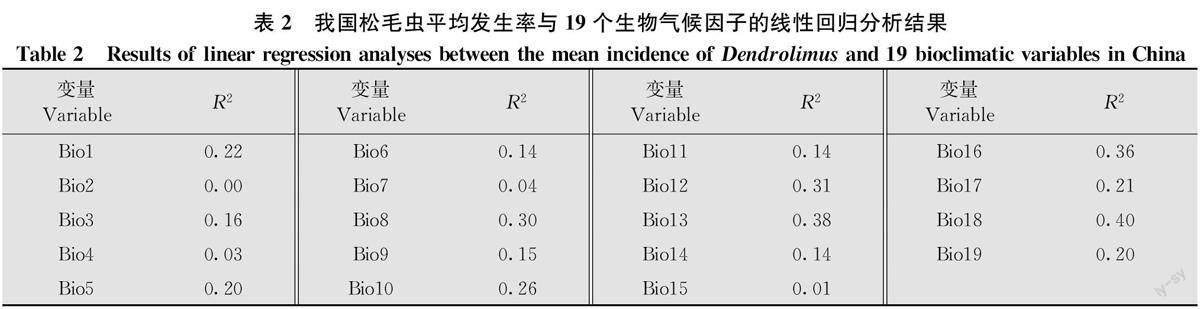
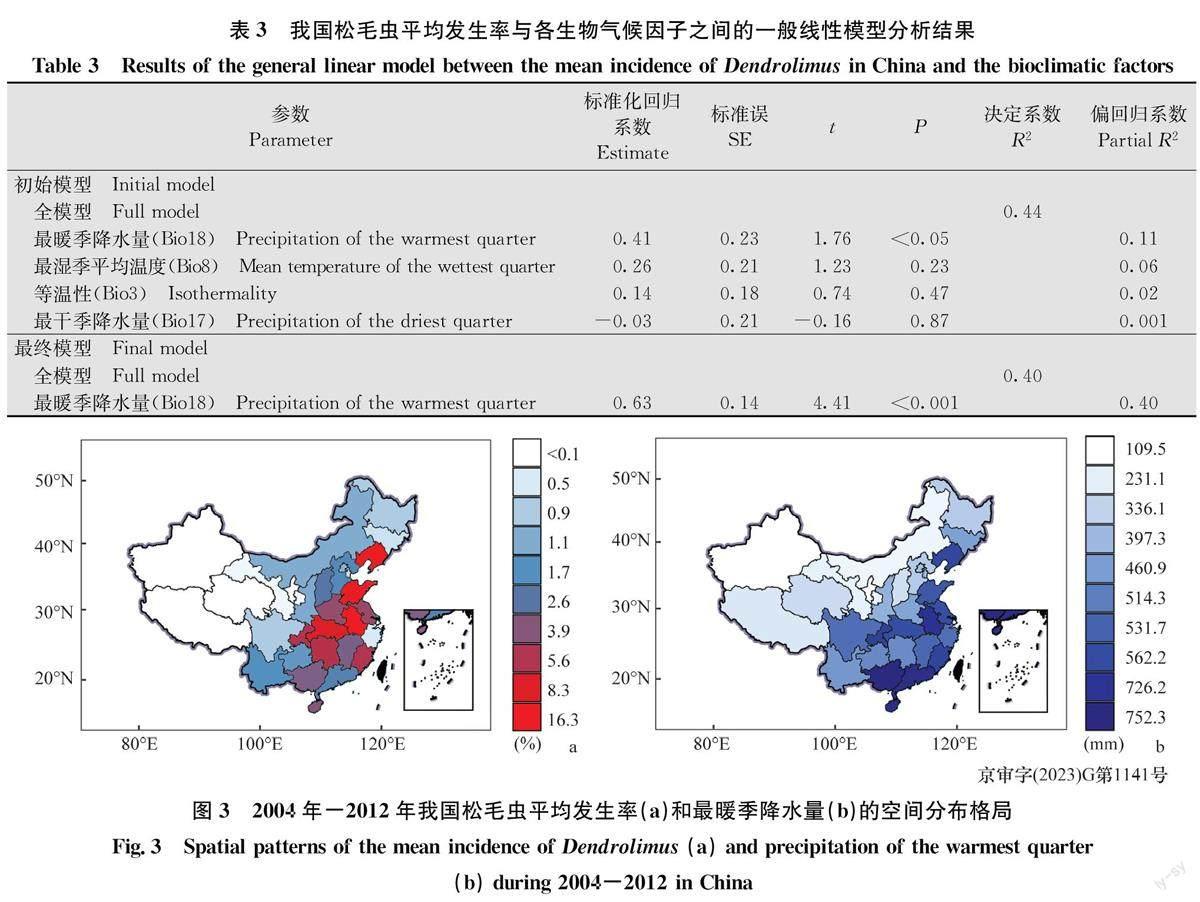
通過Pearson相關系數(表1)和線性回歸分析(表2)篩選出4個生物氣候因子,即Bi018(最暖季降水量,R=0. 40)、Bio8(最濕季平均溫度,R=0.30)、Bi017(最干季降水量,R=0.21)和Bio3(等溫性,R=0.16)。VIF進一步分析發現,Bi018(VIF=2.56)、Bio8(VIF=2.05)、Bio17(VIF=2.06)和Bio3(VIF=1.56)4個生物氣候因子的VIF均小于5,說明四者之間不存在高度共線性,從而保留這4個生物氣候因子。
2.2.2一般線性模型分析結果
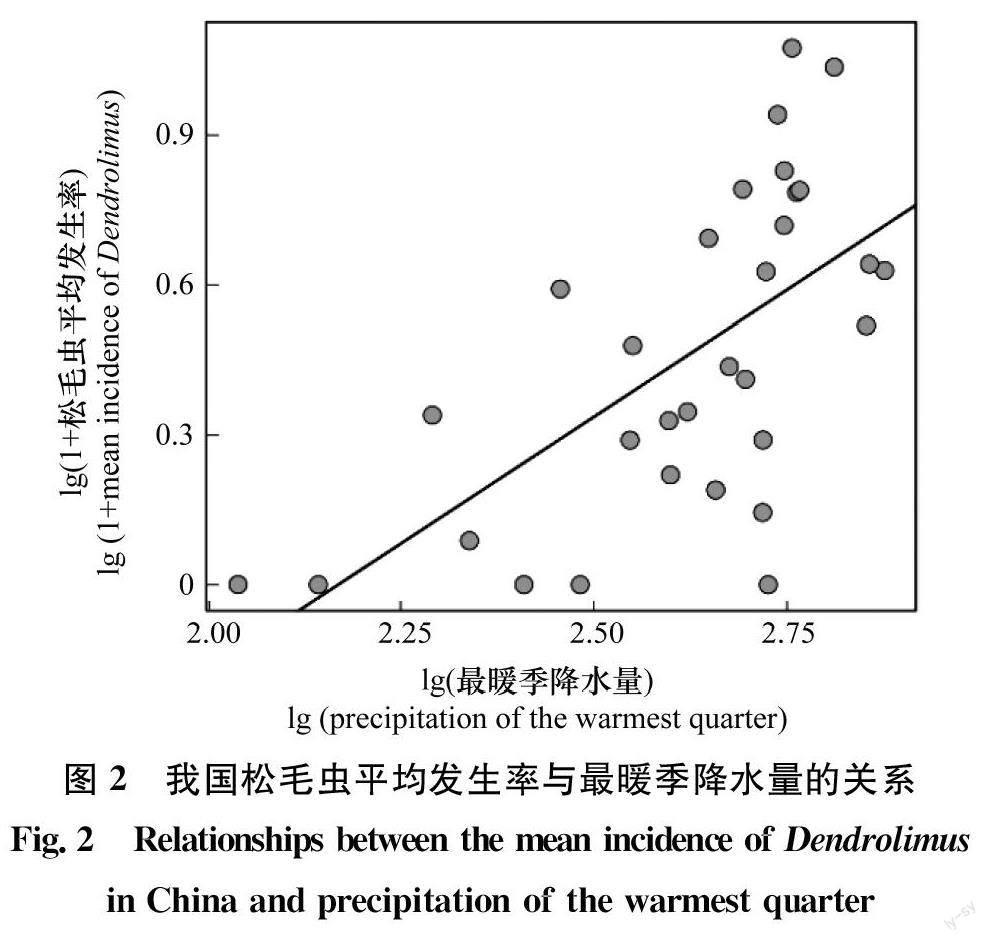
利用一般線性模型初步分析發現,松毛蟲平均發生率與最暖季降水量(P<0.05)顯著相關,而與最濕季平均溫度(P=0.23)、最干季降水量(P=0.87)和等溫性(P=0.47)無關(表3)。因此,最暖季降水量為我國松毛蟲發生率空間分布格局的主要氣候驅動因子。進一步分析發現,松毛蟲平均發生率與最暖季降水量(圖2,表3)呈顯著正相關,最暖季降水量解釋了我國松毛蟲平均發生率40%的空間異質性(表3)。因此,我國松毛蟲的高發區主要集中于最暖季降水量較高的南方和北方部分地區,而最暖季降水量較低的西北和青藏地區則松毛蟲發生率較低(圖3a,b)。
3結論與討論
通過研究2004年—2012年我國松毛蟲發生率空間分布格局及其與氣候因子的相關關系發現,全國尺度上松毛蟲的發生主要受最暖季降水量影響,受溫度影響不大。松毛蟲平均發生率與最暖季降水量(Bio18)呈顯著正相關,因此,我國松毛蟲發生率呈現明顯的空間分布格局。最暖季降水量較高的南方和北方部分地區松毛蟲發生率較高,而降水量較低的西北和青藏地區松毛蟲發生率較低。
最暖季(即夏季)降水量是影響松毛蟲發生最主要的氣候因子,且與松毛蟲發生率顯著正相關,主要原因可能是濕潤環境有利于松毛蟲生長發育并促進其寄主植物生長,這與已有的研究結果一致[1,14]。例如,王慶等研究了2002年—2012年全國馬尾松毛蟲發生率與氣候因子的關系,發現馬尾松毛蟲發生率與夏季降水量正相關。松毛蟲卵和幼蟲喜濕潤環境,其卵孵化率和低齡幼蟲存活率隨空氣濕度增加而上升,而長期處于濕度小于75%的環境中,卵和幼蟲則無法正常生長發育[1,32-33]。在我國發生的6種松毛蟲,盡管它們每年能完成的世代數不同,最長的為2年1代,最短的為1年5代,但是卵孵化和幼蟲生長發育均集中在夏季(例如馬尾松毛蟲在海南省即使1年5代,但也有2代集中在夏季)[27]。因此,夏季降水量越高空氣濕度越大,越有利于卵孵化和幼蟲生長,從而松毛蟲發生的風險越高。此外,較高的夏季降水量能促進寄主植物的生長,提高松針的生物量[34-35],為松毛蟲的生長發育和繁殖提供充足食料,從而增加其發生的風險。我國夏季降水量由東南沿海向西北內陸逐漸減少[36],相對應地,西北和青藏地區為松毛蟲的低發區,而南方和北方部分地區為松毛蟲高發區。然而,雖然廣西、廣東夏季降水量很高,但與相同降雨量(甚至更低)的其他地區相比,松毛蟲發生率較低,這可能與兩廣地區的林分類型和松毛蟲天敵有關。過去兩廣地區為改善由于大面積馬尾松純林引發的馬尾松毛蟲暴發問題,引入闊葉樹種對馬尾松林進行混交改造,因而可能降低了松毛蟲的發生[37]。此外,華南地區(包括廣西、廣東和海南)松毛蟲天敵的數量和種類最豐富,一定程度上也抑制了松毛蟲的暴發[8]。
在全國尺度上,夏季降水量是松毛蟲發生最主要的氣候驅動因子,但也只解釋了松毛蟲發生率40%的空間異質性,說明除氣候因子外,松毛蟲的發生也受其他生物(例如天敵、林分類型和結構等)和非生物因素(例如地形、海拔等)的影響[9,15]。然而,本研究結果仍然在一定程度上揭示了我國松毛蟲發生的空間分布規律及其主要驅動因子,對松毛蟲發生的分區域監測和防控從而減少其發生具有重要指導意義。將來我國各行政區可根據當地的氣候條件對松毛蟲的發生進行監測預報,若當年的夏季降水比往年高,松毛蟲高發區的有關部門應加強對松毛蟲的監測,及時采取預防和防治措施,松毛蟲低發區則進行常規的監測和防控,以減少我國松毛蟲的發生及造成的經濟損失。
