一種新發生的玉米細菌性葉腐病病原菌分離鑒定
2023-08-05 21:43:15王春明郭成周天旺等
植物保護 2023年4期
王春明 郭成 周天旺等


關鍵詞 玉米;細菌性病害;短小芽胞桿菌;分離鑒定;16SrDNA;gyrB
中圖分類號:S 435.131 文獻標識碼:A DOI:10.16688/j.zwbh. 2022445
玉米Zea mays由于其產量高,品質好,適應性強,不僅被作為重要的糧飼兼用型作物,而且被作為重要的工業原料等在生產中加以利用,在國民經濟發展中起著至關重要的作用口]。隨著玉米種植面積增加、新品種引進、氣候條件變化及多年連作等,玉米病害的發生日漸嚴重,已成為制約玉米產業健康發展的主要因素之一[2]。據報道,每年因病害造成的玉米產量損失在一些省份可達到10%~15%,大發生時甚至超過30%[3]。玉米細菌性病害在局部地區發生嚴重,給當地玉米生產造成了不同程度的損失[4-9]。種子和病殘體帶菌是玉米細菌性病害傳播的主要途徑,加之細菌具有繁殖速度快的特點,一旦在田間發生,如未及時采取相應的防治措施,可能對大田和制種田造成毀滅性的損失[10]。
甘肅省作為我國玉米主產區之一,玉米已成為第一大糧食作物。據有關部門統計顯示,至2020年甘肅省玉米種植面積已達到100萬hm,其中制種面積常年穩定在10萬hm左右[11]。玉米病害已成為影響甘肅省玉米產量的重要因素。據調查,近年來一些玉米細菌性葉斑病在甘肅河西和隴中部分地區有逐年上升的趨勢,對玉米生產產生了一定的影響。2021年7月,在白銀市景泰縣條山農場玉米種植區,出現一種新的玉米細菌性葉部病害,為了明確引發該病害的致病菌,本研究從發病玉米葉片中分離菌株,從形態學觀察、致病性測定、生理生化測定、16SrDNA和gyrB基因序列分析等方面進行鑒定,為玉米葉部細菌性病害的流行規律及綜合防治提供理論基礎。
1材料與方法
1.1病樣采集及病原菌分離純化
于2021年7月在甘肅省白銀市景泰縣條山農場普通玉米種植區不同玉米田塊,隨機采集玉米葉腐病病樣共計15份。參考方中達[12]的方法對病原菌進行分離和純化。
1.2煙草過敏性試驗
將分離菌株在NA培養基(蛋白胨5.0 g,牛肉浸膏3.0 g,酵母粉1.0 g,葡萄糖10.0 g,瓊脂粉15.0 g,蒸餾水1000 mL,pH 7.0)上劃線培養36 h后,分別刮下菌苔配成濃度為1×10 cfu/mL的菌懸液,用無菌注射器將菌懸液注射到6~8葉齡煙草葉片背面,以注射滅菌水為對照,室內套袋保濕,觀察有無枯斑產生。
1.3病原菌致病性測定及再分離
玉米種子(品種為‘先玉335’)用75%乙醇表面消毒30 min后,用無菌水沖洗5遍,于35℃培養箱催芽。待胚芽長1cm左右時種植于裝有滅菌蛭石的花盆中,每盆種植7株,3次重復,置于室溫下正常管理。待玉米長到4~6葉期,將培養36 h的分離菌分別刮下菌苔配制成1×10 cfu/mL的菌懸液(加少量吐溫- 20)分別噴霧接種于玉米葉片上,對照接NA培養液,套袋保濕培養24 h后,揭開塑料袋繼續正常管理,逐天觀察發病情況。對發病葉片上的病原進行再分離鑒定,完成柯赫氏法則驗證。
1.4病原菌鑒定
1.4.1形態學鑒定及生理生化測定
將病原細菌在NA平板上劃線培養,參照《植病研究方法》[12],觀察菌落形態、生長速度和產生色素情況。采用革蘭氏染色法,在100倍油鏡下觀察和測量病原細菌菌體大小、形狀及產生芽胞情況。生理生化指標測定參照文獻[12-14]的方法進行。
1.4.2病原菌16S rDNA和gyrB基因序列測定與分析
用天根生化科技(北京)有限公司細菌基因組DNA提取試劑盒提取代表性菌株B1-0和B2-1的全基因組DNA,選擇細菌16S rDNA通用引物27F/1492R[15]和gyrB引物UPl/UP2r[16],參考荊卓瓊等[17]和程勛輝[18]的方法對B2-0和B2-1的16SrDNA和gyrB序列分別進行PCR擴增。反應完成后,取3μL PCR產物進行1%瓊脂糖凝膠電泳檢測。將特異性條帶亮度好、純度高且無非特異性條帶的PCR產物冷凍條件下送生工生物工程(上海)股份有限公司進行測序,測序結果在NCBI上用BLAST進行同源性比較,下載短小芽胞桿菌Bacil-lus pumilus標準菌株及近緣種和外群菌株的目的序列,采用ClustalX 2.0.10軟件進行多序列比對后,利用Bioedit 5.0.6軟件進行校正,用MEGA7.0軟件以NJ法構建基于16S rDNA和gyrB序列的系統發育進化樹。
2結果與分析
2.1田間發生情況調查及病樣分離純化結果
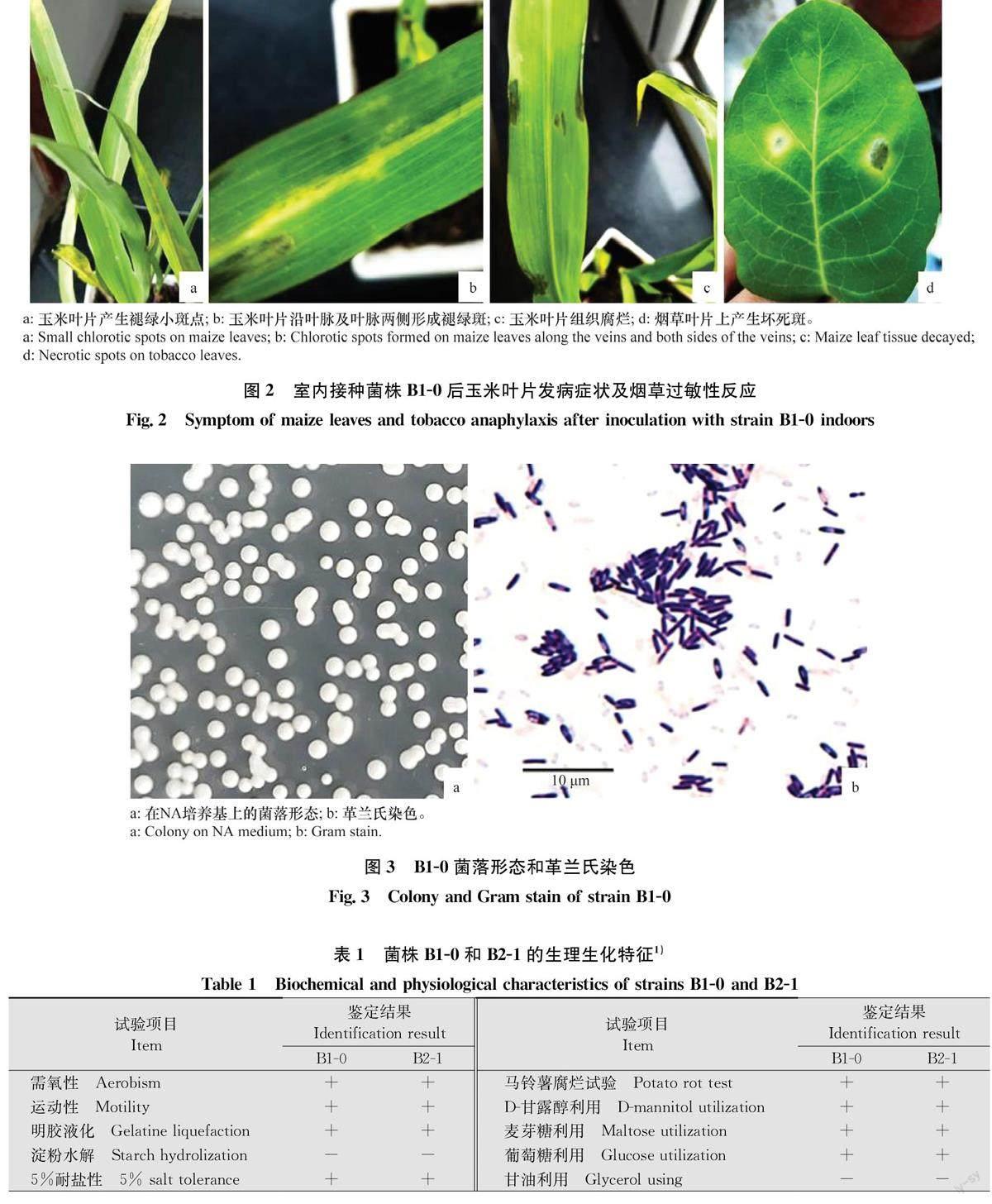
2021年7月,在甘肅省白銀市景泰縣玉米田中新發病害發生較為嚴重,田間病株率22.5%,表現為初期葉片產生不規則褪綠斑塊(圖1a,1b),病斑不受葉脈限制(圖1c),后期病部連片,葉肉組織腐爛,裂開(圖1d,1e)。發病嚴重的引起心葉卷曲,變褐腐爛(圖1f,1g)。觀察發現植株莖稈及根部正常。經對不同發病形狀的病斑進行分離均得到比較單一的1種菌落。挑取代表性菌株B1-0和B2-1進行致病性測定。
2.2致病性測定結果
將菌株B1-0和B2-1回接到玉米葉片24 h后開始顯癥,初期葉片產生褪綠小斑塊(圖2a),隨著病情的進一步發展,3d后病斑逐漸連片形成褪綠長斑塊,形狀不固定,且不受葉脈限制(圖2b)。后期如濕度增加,發病葉片變褐腐爛(圖2c)。接種Bl-0和B2-1的發病率均為100%,發病癥狀與田間自然發病癥狀相似,而對照葉片均未發病。對發病葉片進行再次分離得到與接種病原菌相同的分離物,符合柯赫氏法則。
2.3對煙草致病性測定結果
菌株Bl-0和B2-1接種煙草葉片24 h后開始顯癥,隨后進一步發展形成黃色枯斑(圖2d)。
2.4病原菌培養性狀及形態特征
菌株在NA培養基上菌落為白色、圓形、隆起,表面光滑,不透明,邊緣整齊(圖3a)。培養24~48 h后在100倍油鏡下觀察顯示,菌體短桿狀,大小(0.53~0.71》μm×(1.62~2.93) μm;革蘭氏染色陽性,芽胞中生(圖3b),故從形態學初步判斷其屬于芽胞桿菌Bacillus sp.。
2.5生理生化特性測定結果
試驗結果表明(表1),該病原菌需氧,能運動,能使明膠液化,在5% NaCl中可生長,不能水解淀粉,能利用檸檬酸鹽,接觸酶陽性,甲基紅試驗陰性,能使馬鈴薯腐爛,能利用D-甘露醇、麥芽糖和葡萄糖、不能利用甘油和肌醇,在4℃和40℃條件下均能生長。結合形態學及生理生化特性初步將其鑒定為短小芽胞桿菌B.pumilus。
2.6病原菌16S rDNA和gyrB基因序列分析
將測序結果經Blast相似性分析,菌株B1-0和B2-1的16S rDNA序列與短小芽胞桿菌B.pumilus同源性最高,相似性均在99%以上,且與登錄號為MT065817.1、MT065814.1、MI102721.1、MT102722.1、MT102723.1、MT197380.1和MT065805.1的短小芽胞桿菌B.pumilus相似性達100.00%;gyrB基因序列與登錄號為GU568184.1、KF194241.1、HM585095.1和MF784845.1的短小芽胞桿菌B.pumilus相似性均在99.00%以上。在GenBank中下載短小芽胞桿菌B.pumilus標準菌株及其近緣種和外群菌株的目的基因序列,比對后拼接,用MEGA 7.0軟件構建基于16S rDNA和gyrB的系統發育樹(圖4),B1-0和B2-1與短小芽胞桿菌B.pumilus (ATCC7061、BKS1-108、AUEC29、BP-hd-1、NMTD17和17)聚在一個分支。說明菌株Bl-0和B2-1與短小芽胞桿菌B.pumilus遺傳距離更近,結合形態學及分子生物學鑒定將其確定為短小芽胞桿菌B.pumilus。
3結論與討論
目前國內外報道的引起玉米葉部病害的病原細菌主要有須芒草伯克霍爾德氏菌Burkholderia an-dropogonis(異名高梁假單胞菌Pseudomonas an-dropogonis)[14,19]、燕麥嗜酸菌燕麥亞種Acidovorax avenae subsp. avenae[4,20]、斯氏泛菌Pantoea stew-artii[19-20]、菠蘿泛菌P.ananatis[5,21-22]、野油菜黃單胞菌絨毛草致病變種Xanthomonas campestrzs pv.holcicola E23]、丁香假單胞菌丁香致病變種P.syrin-gae pv. syringaeE23-24]、巨大芽胞桿菌Bacillus megaterium [25]等,分別可引起玉米葉片條紋病、葉疫病、枯萎病、葉斑病、褐斑病等,但未見短小芽胞桿菌能引起玉米葉部病害的報道。
短小芽胞桿菌作為芽胞桿菌屬Bacillus的一種細菌,被作為生防菌在國內外研究居多。于婷等[26]篩選出一株短小芽胞桿菌對鐮刀菌有較強的抑制作用;Agarwal等[27]研究結果顯示短小芽胞桿菌MSUA3對供試的尖鐮孢Fusarium oxysporum和立枯絲核菌Rhizoctonia solani抑制效果強;柳自清等[28]研究表明短小芽胞桿菌B102對棉花枯萎病具有較好的防治效果;馮永新等[29]發現短小芽胞桿菌AR03與噻菌銅聯合復配后能增強對煙草青枯病的防效。同時查閱文獻可知,其作為植物病原菌的相關報道也在不斷增多,如有研究表明該病原菌能引起幼齡桃Prunus persica細菌性斑疹病[30]、芒果Mangifera indica枯萎病[31]、菜豆Phaseolus vul-garis葉片褐斑病[32]、馬鈴薯Solanum tuberosum貯藏期塊莖軟腐病[33]、生姜Zingiber of icinale根腐病[34],且侵染歐洲松樹Pinus的莖和枝干造成發病部位變黃、枝干枯萎,針葉脫落等[35],還能引起甜瓜Cucumis melo果腐病[36]、雅榕Ficus concinna果腐病[37]和橡皮樹Ficus elastica枝干凸起病等[38];曹慧英等[39]2010年研究發現該病原菌能引起玉米細菌性莖腐病,其典型癥狀為莖基處開裂,變黑、變褐并出現干腐。本研究在田間發現的玉米細菌性病害,發病玉米葉片可產生不同大小的不受葉脈限制的不規則褪綠斑塊,隨著病情的進一步發展出現葉片開裂、腐爛等癥狀,發病嚴重的植株心葉卷曲、腐爛,無法抽出,嚴重影響后期玉米的生長發育。挖出發病植株觀察發現,植株莖基部及根部均正常,未出現變褐、開裂等癥狀,這與曹慧英等[39]的研究存在差異。通過形態學觀察、生理生化測定、16S rDNA及gyrB基因序列測定結果,菌株B1-0和B2-1與短小芽胞桿菌相似度最高,且在基于16S rDNA和gyrB的系統發育樹上與短小芽胞桿菌(ATCC 7061、BKS1-108、AUEC29、BP-hd-1、NMTD17和17)聚在一個分支,故將其鑒定為短小芽胞桿菌。但本研究獲得的菌株在生理生化特性上與已報道的短小芽胞桿菌菌株存在一定的差異,如Song等[36]研究報道稱短小芽胞桿菌不能使明膠液化,而本研究結果顯示供試菌株均能使明膠液化;曹慧英等[39]研究表明短小芽胞桿菌能水解淀粉,能利用甘露醇,不能利用葡萄糖,而本研究結果表明供試菌株不能水解淀粉,不能利用甘露醇、能利用葡萄糖。分析產生這種差異的原因主要可能與其來源及生長環境等存在一定的差異相關,使得不同來源的同種病原菌即使在相同的培養基和相同的培養條件下,在生理生化特性方面也呈現差異。
傅俊范等[2]對玉米葉部病害流行成災的原因分析發現,隨著栽培制度和品種抗病性的改變、田間病原菌積累以及一些極端天氣導致田間小氣候更適宜病害發生,是玉米葉部病害流行成災的根本原因。玉米細菌性病害的發生除與品種的抗病性有關外,還與環境溫度和濕度條件有較大的關系,一般溫暖多雨有利于細菌性病害的發生。對比2021年5月至7月上旬白銀市景泰縣的天氣情況,降雨量較往年有所增加,氣溫略低,更適宜細菌性病害的發生,因此導致細菌性葉腐病在該地區發生嚴重。查閱國內外報道的玉米細菌性葉部病害文獻,發現對玉米細菌性葉部病害的報道多停留在病害發生癥狀和病原菌的鑒定上,而對其發病規律、傳播途徑、品種抗性及綜合防治等發面的系統研究較少,因此有待于后期進一步的研究。
本研究通過形態學觀察、生理生化指標測定、16S rDNA和gyrB基因序列分析和致病性測定等研究,明確了引起甘肅省白銀市景泰縣玉米田一種細菌性葉腐病的病原菌為短小芽胞桿菌B.pumi-lus,該研究結果可為該病害的發生流行和綜合防治提供參考。
