施肥方式對(duì)連作甘薯田土壤團(tuán)聚體穩(wěn)定性及酶活性的影響
2023-08-14 07:11:48李敏劉亞軍王文靜胡啟國王紅剛儲(chǔ)鳳麗
江蘇農(nóng)業(yè)科學(xué) 2023年13期
李敏 劉亞軍 王文靜 胡啟國 王紅剛 儲(chǔ)鳳麗


摘要:研究施肥措施對(duì)連作甘薯田土壤團(tuán)聚體穩(wěn)定性及酶活性變化的影響,為豫東地區(qū)連作甘薯田有機(jī)肥的合理施入提供理論依據(jù)。試驗(yàn)設(shè)不施肥(CK)、單施化肥(SF)、單施生物有機(jī)肥(SBF)、單施緩控釋肥(SSF)、50%緩控釋肥50%生物有機(jī)肥(SF50BF50)、50%化肥配施50%生物有機(jī)肥(F50BF50)6個(gè)處理,測(cè)定了不同施肥處理土壤團(tuán)聚體組成、穩(wěn)定性、容重、孔隙度、全土酶活性以及團(tuán)聚體酶活性等指標(biāo)。結(jié)果表明,與CK相比,化肥、緩控釋肥與生物有機(jī)肥配施能夠提高>2 mm、>1~2 mm粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)和團(tuán)聚體GMD、MWD和R>0.25值,降低D值;能夠提高土壤孔隙度,降低土壤容重。其中,F(xiàn)50BF50處理土壤容重顯著降低6.71%,SF50BF50、F50BF50處理土壤孔隙度分別顯著提高7.08%、10.27%;能夠提高全土以及團(tuán)聚體酶活性和>2 mm、>1~2 mm粒級(jí)團(tuán)聚體脲酶、堿性磷酸酶、蔗糖酶活性貢獻(xiàn)率。其中,F(xiàn)50BF50處理全土脲酶、過氧化氫酶、蔗糖酶活性分別顯著提高13.58%、16.90%、16.82%,SF50BF50 處理全土堿性磷酸酶活性顯著提高10.71%;相關(guān)性分析結(jié)果顯示,土壤酶活性變化與土壤結(jié)構(gòu)及團(tuán)聚體穩(wěn)定性變化緊密相連。綜上所述,化肥、緩控釋肥與生物有機(jī)肥配施能夠改善土壤結(jié)構(gòu)和團(tuán)聚體組成,提升團(tuán)聚體穩(wěn)定性,提高團(tuán)聚體酶活性及貢獻(xiàn)率。其中,50%化肥配施50%生物有機(jī)肥處理表現(xiàn)較好。
關(guān)鍵詞:施肥;團(tuán)聚體;容重;孔隙度;酶活性
中圖分類號(hào):S531.06;S152.3? 文獻(xiàn)標(biāo)志碼:A
文章編號(hào):1002-1302(2023)13-0252-09
甘薯是我國重要的糧食作物,因具有產(chǎn)量高、易栽活、營養(yǎng)豐富等特點(diǎn),在我國具有較大的種植面積[1-2]。近年來,隨著甘薯產(chǎn)業(yè)升級(jí)以及能源作物的開發(fā)利用,甘薯需求量逐年遞增[3-4]。而受耕地面積、輪作倒茬時(shí)間限制,廣大種植戶在保障甘薯產(chǎn)量的前提下,常年連作種植以及大量施用化肥,造成甘薯根際土壤結(jié)構(gòu)改變,養(yǎng)分及酶活性降低,菌群失衡等問題[5-8]。且隨著施肥時(shí)間的延長,連作甘薯田產(chǎn)量下降,品質(zhì)降低,病蟲害頻發(fā),嚴(yán)重影響了甘薯產(chǎn)值及產(chǎn)業(yè)健康發(fā)展[9-10]。因此,改善連作甘薯田土壤結(jié)構(gòu),提高土壤生物學(xué)活性等問題迫在眉睫。
土壤團(tuán)聚體是由較小土壤顆粒膠結(jié)而成的粒狀或塊狀結(jié)構(gòu)體,是土壤結(jié)構(gòu)的基本單元,也是土壤養(yǎng)分的儲(chǔ)存庫和微生物活動(dòng)的重要場(chǎng)所,其含量的變化能夠直接影響土壤物理結(jié)構(gòu)以及養(yǎng)分供應(yīng)能力[11]。土壤酶是農(nóng)田土壤的重要組成部分,能夠參與土壤中大多數(shù)復(fù)雜的生物化學(xué)過程,是指示土壤肥力和生物學(xué)特性的重要指標(biāo)[12]。有研究表明,無機(jī)肥與有機(jī)肥配施能夠改變土壤結(jié)構(gòu),提高土壤酶活性[13-15]。甘雅芬等研究表明,化肥減量與紫云英還田能夠改善土壤團(tuán)聚體粒徑分布,促進(jìn)較大團(tuán)聚體形成,提高土壤團(tuán)聚體穩(wěn)定性[16];聶鑫等研究表明,化肥減量20%配施紫云英30 000 kg/hm2還田能夠顯著促進(jìn)土壤中>1~2 mm、0.5~1 mm粒級(jí)團(tuán)聚體形成,提高土壤團(tuán)聚體穩(wěn)定性,但隨著紫云英還田量增加,>2 mm粒級(jí)團(tuán)聚體比例及團(tuán)聚體穩(wěn)定性降低[17];張帥等研究表明,與單施化肥或生物炭相比,化肥減量配施生物炭能夠顯著提高 >2 mm、>0.25~2 mm、0.053~0.25 mm粒徑團(tuán)聚體相關(guān)碳轉(zhuǎn)化酶活性,降低>0.25~2 mm、0.053~0.25 mm 粒徑團(tuán)聚體微生物熵[18];李委濤等研究表明,化肥與有機(jī)肥配施能夠顯著提高土壤團(tuán)聚體平均當(dāng)量直徑,增加土壤酶活性,改善土壤團(tuán)聚體粒徑結(jié)構(gòu),提高土壤生產(chǎn)力土壤[19]。
緩控釋肥能夠根據(jù)作物需肥特性調(diào)控施肥,具有肥效長、肥效高等優(yōu)點(diǎn),能夠?qū)崿F(xiàn)簡化施肥,省時(shí)省力[20]。生物有機(jī)肥含有豐富的營養(yǎng)物質(zhì)和功能菌群,施入土壤中能夠改善微生態(tài)環(huán)境,促進(jìn)微生物繁殖,提高土壤微生物活性[21]。當(dāng)前,緩控釋肥、生物有機(jī)肥在玉米、水稻方面的研究有很多,在甘薯方面的研究相對(duì)較少,而關(guān)于化肥、緩控釋肥與生物有機(jī)肥配施對(duì)甘薯田土壤團(tuán)聚體酶活性影響方面的研究更是未見報(bào)道[22-24]。因此,本研究通過化肥、緩控釋肥減量與生物有機(jī)肥配施,探究連作甘薯田土壤團(tuán)聚體粒徑分布、穩(wěn)定性、容重、孔隙度、酶活性及團(tuán)聚體酶活性貢獻(xiàn)率的變化規(guī)律,找到適宜的施肥模式,旨在為豫東地區(qū)連作甘薯田有機(jī)肥的合理施入提供理論依據(jù)。
1 材料與方法
1.1 研究區(qū)概況
試驗(yàn)于2020—2022年在河南省商丘市梁園區(qū)雙八鎮(zhèn)朱莊村(116°37′ E,39°93′ N)進(jìn)行。試驗(yàn)區(qū)域?qū)倥瘻貛О霛駶櫞箨懶约撅L(fēng)氣候,年均溫度 14.2? ℃,年均降水量675 mm,其中約80%降水量集中在6—9月,無霜期212 d,年日照時(shí)數(shù) 2 200 h。供試土壤為黃潮土黏土質(zhì),基礎(chǔ)土壤(0~20 cm)理化性質(zhì):含全氮0.92 g/kg、全磷0.65 g/kg、速效鉀123.37 mg/kg、堿解氮54.69 mg/kg、速效磷 37.68 mg/kg、有機(jī)質(zhì)11.26 g/kg,pH值為 8.02。試驗(yàn)地塊平整,便于排灌,常年小麥—甘薯輪作種植。
1.2 供試材料
供試品種:商薯18(商丘市農(nóng)林科學(xué)院生物研究所);商麥167(商丘市農(nóng)林科學(xué)院國家農(nóng)作物區(qū)試站)。
供試肥料,復(fù)合肥(N、P2O5、K2O含量分別為10%、10%、20%,河南億豐年生物科技有限公司);緩控釋肥(N、P2O5、K2O含量分別為18%、9%、18%,湖北金峰農(nóng)業(yè)科技有限公司);生物有機(jī)肥(芽孢桿菌≥0.5億CFU/g,黃腐酸含量≥12%,有機(jī)質(zhì)含量≥40%,山東泉林嘉有機(jī)肥料有限責(zé)任公司)。
1.3 試驗(yàn)設(shè)計(jì)
試驗(yàn)地種植制度常年冬季種植小麥,夏季種植甘薯。小麥季施肥不做特殊處理,按照當(dāng)?shù)胤N植習(xí)慣進(jìn)行,甘薯季采用不同施肥措施處理。試驗(yàn)設(shè)不施肥(CK)、單施化肥(SF)、單施生物有機(jī)肥(SBF)、單施緩控釋肥(SSF)、50%緩控釋肥50%生物有機(jī)肥(SF50BF50)、50%化肥配施50%生物有機(jī)肥(F50BF50)6個(gè)處理,各處理重復(fù)3次,共18個(gè)小區(qū),隨機(jī)區(qū)組排列。小區(qū)面積為56 m2,甘薯株行距為0.28 m×0.8 m,走道0.8 m,保護(hù)行3.0 m。單施化肥、緩控釋肥、生物有機(jī)肥用量分別為750、750、1 200 kg/hm2。不同施肥處理肥料均作為基肥在旋耕起壟前一次性施入,生育期內(nèi)不進(jìn)行追肥。小麥秸稈、甘薯地上部植株均通過粉粹機(jī)直接還田。其他田間種植、管理措施按照本地習(xí)慣進(jìn)行,不做特殊處理。小麥生育期10月20日至翌年6月8日,甘薯生育期6月15日至10月15日。
1.4 樣品采集與項(xiàng)目測(cè)定
樣品采集與處理。于2022年甘薯收獲前1周通過螺旋土鉆利用五點(diǎn)取樣法采集0~30 cm土壤樣品,混勻后帶回實(shí)驗(yàn)室。撿出根系、碎石等雜物后,將土壤樣品分成2個(gè)部分,一部分?jǐn)傞_自然風(fēng)干,用于全土酶活性指標(biāo)的測(cè)定;另一部分沿其土壤結(jié)構(gòu)輕輕剝開,盡量避免土塊因外力發(fā)生形變,自然風(fēng)干后用于土壤團(tuán)聚體組成、團(tuán)聚體酶活性等指標(biāo)的測(cè)定。在用土鉆采集樣品的同時(shí),利用環(huán)刀在每個(gè)小區(qū)內(nèi)采集0~20 cm環(huán)刀樣品1份,用于土壤容重、孔隙度指標(biāo)的測(cè)定。
測(cè)試項(xiàng)目與方法。土壤脲酶、過氧化氫酶、堿性磷酸酶、蔗糖酶活性分別采用苯酚鈉比色法、高錳酸鉀滴定法、磷酸苯二鈉比色法和3,5-二硝基水楊酸比色法[25]測(cè)定。土壤容重、孔隙度均采用環(huán)刀法[26]測(cè)定。土壤團(tuán)聚體組成采取干篩法測(cè)定[27]。然后分別測(cè)定粒徑>2 mm、>1~2 mm、>0.25~1 mm、0.053~0.25 mm 和<0.053 mm團(tuán)聚體質(zhì)量,用于土壤團(tuán)聚體穩(wěn)定性指標(biāo)計(jì)算。土壤團(tuán)聚體穩(wěn)定性及酶活性貢獻(xiàn)率計(jì)算公式[28-29]如下:
GMD=exp∑ni=1milnXi∑ni=1mi;
D=3-lgW(δ<Xi)WtlgXiXmax;
MWD=∑ni=1Xi·Wi;
R>0.25=mt>0.25/mt;
團(tuán)聚體貢獻(xiàn)率=(某粒級(jí)團(tuán)聚體酶活性×某粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù))/全土酶活性×100%。
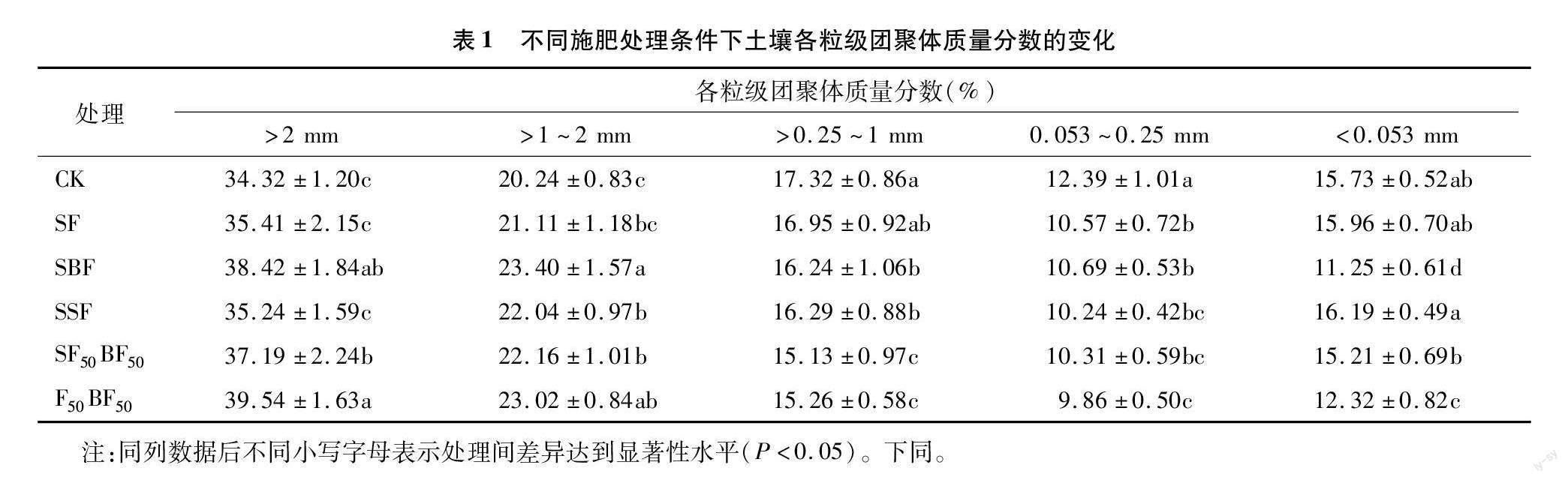
式中:MWD為團(tuán)聚體平均質(zhì)量直徑;D為分形維數(shù);GMD為幾何平均直徑;R>0.25為粒徑 >0.25 mm 團(tuán)聚體質(zhì)量分?jǐn)?shù);mi為i粒徑團(tuán)聚體質(zhì)量;Xi為i粒徑團(tuán)聚體平均直徑;Xmax為最大粒徑團(tuán)聚體平均直徑;Wi為i粒徑團(tuán)聚體質(zhì)量占比;W(δ 1.5 數(shù)據(jù)處理與分析 采用Excel 2018進(jìn)行原始數(shù)據(jù)整理與計(jì)算,采用SPSS 19.0進(jìn)行方差分析與相關(guān)性分析。其中,不同處理間差異顯著性采用新復(fù)極差法進(jìn)行檢驗(yàn)(α=0.05)。 2 結(jié)果與分析 2.1 不同施肥處理對(duì)土壤團(tuán)聚體粒徑分布特征的影響 由表1可知,連續(xù)3年不同施肥處理?xiàng)l件下土壤各粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)表現(xiàn)出明顯差異。與CK相比,SBF、SF50BF50、F50BF50處理>2 mm粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)分別顯著提高11.95%、8.36%、15.21%,SF、SSF處理無顯著性變化。其中,不同施肥處理間F50BF50處理最大,顯著高于SF、SSF、SF50BF50處理;SBF、SSF、SF50BF50、F50BF50處理>1~2 mm 粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)較CK分別顯著提高15.61%、8.89%、9.49%、13.74%,SF處理無顯著性變化。其中,不同施肥處理間SBF處理最大,顯著高于SF、SSF、SF50BF50處理;SBF、SSF、SF50BF50、F50BF50處理>1~2 mm粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)較CK分別顯著降低6.24%、5.95%、12.64%、11.89%,SF處理無顯著性變化。其中,SF50BF50、F50BF50處理顯著低于其他施肥處理。與CK相比,不同施肥處理0.053~0.25 mm粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)顯著降低13.72%~20.42%。其中,F(xiàn)50BF50處理最小,顯著低于SF、SBF處理。SBF、F50BF50處理0.053 mm粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)較CK分別顯著降低28.48%、21.68%,其他施肥處理均無顯著性變化。 2.2 不同施肥處理對(duì)土壤團(tuán)聚體穩(wěn)定性的影響 不同施肥處理土壤團(tuán)聚體穩(wěn)定性變化見表2。與CK相比,其他施肥處理土壤團(tuán)聚體幾何平均直徑(GMD)、平均質(zhì)量直徑(MWD)、>0.25 mm粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)(R>0.25)均不同程度提高,分形維數(shù)(D)均降低。其中,F(xiàn)50BF50處理GMD值最大,較CK、SF、SSF處理分別顯著提高10.48%、7.87%、7.03%,與其他處理相比無顯著性差異。SBF處理GMD值次之,但也均顯著高于CK、SF、SSF處理。SBF處理MWD值、R>0.25值均最大,較CK、SF、SSF處理分別顯著提高11.89%、7.81%、5.61%和8.60%、6.25%、6.08%,與F50BF50處理相比均無顯著性差異。與CK相比,不同施肥處理D值均降低,但均無顯著性差異,不同施肥處理間也均無顯著性差異。 2.3 不同施肥處理對(duì)土壤容重與孔隙度的影響 不同施肥處理土壤容重與孔隙度變化見圖1。與CK相比,不同施肥處理土壤容重降低,孔隙度增加。其中,SBF、F50BF50處理土壤容重較CK分別顯著降低5.37%、6.71%,其他施肥處理土壤容重均無顯著性變化。不同施肥處理間F50BF50處理土壤容重最低,較SSF處理顯著降低5.44%,與其他處理相比均無顯著性差異。SBF、SF50BF50、F50BF50處理土壤孔隙度較CK分別顯著提高9.37%、7.08%、10.27%,SF、SSF處理土壤孔隙度無顯著性變化。不同施肥處理間F50BF50處理土壤孔隙度最大,較其他施肥處理增加0.82%~7.71%,顯著高于SF、SSF處理。 2.4 不同施肥處理對(duì)全土酶活性的影響 不同施肥處理土壤全土酶活性變化見圖2。與CK相比,SBF、SF50BF50、F50BF50處理土壤脲酶活性分別顯著提高9.88%、7.41%、13.58%,SF、SFF處理無顯著性變化。其中,F(xiàn)50BF50處理脲酶活性最高,較其他施肥處理提高5.75%~10.84%,顯著高于除SBF處理外的其他施肥處理。與CK相比,SF、SF50BF50、F50BF50處理土壤堿性磷酸酶活性分別顯著提高5.36%、10.71%、9.82%,SF、SBF處理無顯著性變化。其中,SF50BF50處理堿性磷酸酶活性最高,但與F50BF50處理相比無顯著性差異。不同施肥處理土壤過氧化氫酶、蔗糖酶活性較CK分別顯著提高6.90%~16.90%、5.42%~16.82%。其中,F(xiàn)50BF50處理過氧化氫酶、蔗糖酶活性均最高,過氧化氫酶活性較SF、SBF、SSF處理分別顯著提高6.96%、9.03%、8.33%,蔗糖酶活性較SF、SSF、SF50BF50 處理分別顯著提高10.81%、6.40%、8.97%。SF50BF50處理過氧化氫酶與SBF處理蔗糖酶活性次之,與F50BF50處理相比均無顯著性差異。 2.5 不同施肥處理對(duì)土壤團(tuán)聚體酶活性的影響 不同施肥處理各粒級(jí)團(tuán)聚體酶活性變化見圖3。與CK相比,不同施肥處理各粒級(jí)團(tuán)聚體脲酶、堿性磷酸酶、過氧化氫酶、蔗糖酶活性均有所提高。其中,F(xiàn)50BF50處理>2 mm、>1~2 mm、>0.25~1 mm、<0.053 mm粒級(jí)團(tuán)聚體脲酶活性最高,較CK分別顯著提高16.05%、13.68%、16.09%、22.67%,顯著高于SF、SBF、SSF處理;SF50BF50處理活性次之,除<0.053 mm粒級(jí)活性顯著低于 F50BF50 處理外,其他粒級(jí)均無顯著性差異。SF50BF50 處理0.053~0.25 mm粒級(jí)團(tuán)聚體脲酶活性最高,較CK顯著提高14.10%,顯著高于除 F50BF50 處理外的其他處理,F(xiàn)50BF50處理活性次之。 圖3中F50BF50處理>2 mm、>1~2 mm、>0.25~1 mm、<0.053 mm粒級(jí)團(tuán)聚體堿性磷酸酶活性最高,較CK分別顯著提高14.15%、26.36%、14.05%、20.20%;>2 mm、>1~2 mm粒級(jí)活性顯著高于SF、SBF、SSF處理,>0.25~1 mm、<0.053 mm粒級(jí)活性顯著高于其他處理;SF50BF50處理>2 mm、>1~2 mm、<0.053 mm粒級(jí)和SBF處理>0.25~1 mm粒級(jí)活性次之。SF50BF50處理0.053~0.25 mm粒級(jí)團(tuán)聚體堿性磷酸酶活性最高,較CK顯著提高9.17%,顯著高于SF、SSF處理,F(xiàn)50BF50 處理活性次之。 圖3中F50BF50處理>1~2 mm、>0.25~1 mm、0.053~0.25 mm粒級(jí)團(tuán)聚體過氧化氫酶活性最高,較CK分別顯著提高16.85%、19.02%、21.99%,顯著高于SF、SBF、SSF處理;SF50BF50 處理活性次之。SF50BF50處理>2 mm、<0.053 mm 粒級(jí)過氧化氫酶活性最高,較CK分別顯著提高21.58%、13.53%,顯著高于SF、SBF、SSF處理,F(xiàn)50BF50 處理活性次之。 圖3中F50BF50處理>2 mm、>1~2 mm粒級(jí)團(tuán)聚體蔗糖酶活性均最高,較CK分別顯著提高21.41%、16.81%。>2 mm粒級(jí)活性顯著高于SF、SBF、SSF處理,>1~2 mm粒級(jí)活性顯著高于SF、SSF、SF50BF50處理;SF50BF50處理>2 mm粒級(jí)與 SBF>1~2 mm粒級(jí)活性次之。SBF處理>0.25~1 mm、0.053~0.25 mm、<0.053 mm粒級(jí)蔗糖酶活性最高,較CK分別顯著提高15.76%、10.51%、12.07%,顯著高于SF、SBF、SSF處理,F(xiàn)50BF50 處理蔗糖酶活性次之。 2.6 不同施肥措施條件下土壤各粒級(jí)團(tuán)聚體對(duì)酶活性的貢獻(xiàn)率變化 不同施肥處理土壤各粒級(jí)團(tuán)聚體對(duì)土壤酶活性貢獻(xiàn)率的變化見圖4。與CK相比,不同施肥處理>2 mm、>1~2 mm粒級(jí)團(tuán)聚體對(duì)各種酶活性的貢獻(xiàn)率均有所提高,0.053~0.25 mm粒級(jí)團(tuán)聚體對(duì)各種酶活性的貢獻(xiàn)率以及>0.25~1 mm、<0.053 mm 粒級(jí)團(tuán)聚體對(duì)過氧化氫酶、蔗糖酶活性的貢獻(xiàn)率均有所降低,>0.25~1 mm、<0.053 mm粒級(jí)團(tuán)聚體對(duì)脲酶、堿性磷酸酶活性的貢獻(xiàn)率表現(xiàn)出不同的變化。其中,SF50BF50處理>2 mm與>1~2 mm 粒級(jí)團(tuán)聚體對(duì)脲酶、蔗糖酶活性的貢獻(xiàn)率最高,較CK分別顯著提高18.39%、22.10%與18.96%、14.98%;F50BF50處理>2 mm粒級(jí)團(tuán)聚體對(duì)堿性磷酸酶的貢獻(xiàn)率最高,較CK分別顯著提高19.77%、19.48%,除SF50BF50處理>1~2 mm粒級(jí)團(tuán)聚體對(duì)堿性磷酸酶活性的貢獻(xiàn)率顯著低于F50BF50處理外,其他SF50BF50處理指標(biāo)與F50BF50處理均無顯著性差異。而SF50BF50、F50BF50處理 >0.25~1 mm、0.053~0.25 mm、<0.053 mm粒級(jí)團(tuán)聚體對(duì)各種酶活性的貢獻(xiàn)率較CK均有所降低。整體來看,>2 mm粒級(jí)團(tuán)聚體對(duì)脲酶、堿性磷酸酶、過氧化氫酶、蔗糖酶活性的貢獻(xiàn)率最高,可達(dá)30.15%~40.63%;其次為>1~2 mm粒級(jí)團(tuán)聚體,達(dá)到20.96%~30.51%;0.053~0.25 mm粒級(jí)團(tuán)聚體酶活性貢獻(xiàn)率最低,僅為9.14%~12.23%。 2.7 土壤酶活性與土壤結(jié)構(gòu)指標(biāo)及團(tuán)聚體穩(wěn)定性的相關(guān)性分析 不同施肥處理?xiàng)l件下土壤酶活性與土壤結(jié)構(gòu)指標(biāo)及團(tuán)聚體穩(wěn)定性的相關(guān)性分析見表3。脲酶、蔗糖酶與孔隙度、團(tuán)聚體穩(wěn)定性GMD值、MWD值、R>0.25值呈極顯著正相關(guān)(P<0.01),與容重、D值呈極顯著負(fù)相關(guān)(P<0.01);堿性磷酸酶與孔隙度、GMD值、MWD值、R>0.25值呈正相關(guān),與容重、D值呈負(fù)相關(guān);過氧化氫酶與孔隙度、GMD值、MWD值、R>0.25值呈正相關(guān),與容重、D值呈顯著負(fù)相關(guān)(P<0.05)。由此可見,土壤酶活性變化與土壤結(jié)構(gòu)及團(tuán)聚體穩(wěn)定性變化緊密相連,說明不同施肥措施可以通過改善土壤物理結(jié)構(gòu), 提升團(tuán)聚體穩(wěn)定性來促進(jìn)土壤相關(guān)酶活性的提高。 3 討論與結(jié)論 3.1 施肥措施對(duì)土壤團(tuán)聚體、容重及孔隙度的影響 土壤團(tuán)聚體粒徑分布變化不僅能夠調(diào)控土壤養(yǎng)分供應(yīng),還能夠改變土壤結(jié)構(gòu)與組成[30]。長期施肥對(duì)土壤團(tuán)聚體組成及穩(wěn)定性具有一定的影響,有研究表明,長期施用有機(jī)肥能夠促進(jìn)土壤大粒徑團(tuán)聚體形成[31]。但也有研究表明,外源有機(jī)物的攝入對(duì)土壤團(tuán)聚體組成的影響并不顯著,甚至能夠?qū)е峦寥缊F(tuán)聚體穩(wěn)定性降低[32]。本研究結(jié)果表明,化肥、緩控釋肥配施生物有機(jī)肥或單施生物有機(jī)肥能夠提高土壤>2 mm、>1~2 mm粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)以及團(tuán)聚體GMD、MWD和R>0.25值,降低D值;能提高土壤孔隙度,降低土壤容重。這與李銳等的研究[33]較為一致。分析其原因可能是外源有機(jī)物的攝入能夠?yàn)橥寥垒^小顆粒的團(tuán)聚提供較多的膠結(jié)物質(zhì),且外源有機(jī)物的攝入能夠?yàn)橥寥栏H微生物的活動(dòng)提供較多的碳源,進(jìn)而增強(qiáng)其代謝活性產(chǎn)生較多的代謝膠結(jié)產(chǎn)物,促使大粒徑團(tuán)聚體形成,提高土壤團(tuán)聚體穩(wěn)定性和土壤孔隙度,降低土壤容重,從而改善土壤結(jié)構(gòu)[34-35]。而不同生物有機(jī)肥處理間的差異可能是由外源有機(jī)物的攝入量引起,也可能是無機(jī)肥與有機(jī)肥配施前期更宜滿足作物根系吸收利用,從而促進(jìn)養(yǎng)分轉(zhuǎn)化與分解,產(chǎn)生更多的膠結(jié)物質(zhì),進(jìn)而有利于大顆粒團(tuán)聚體形成[36]。 3.2 施肥措施對(duì)土壤團(tuán)聚體酶活性及貢獻(xiàn)率的影響 土壤酶活性受田間耕作方式、施肥措施影響較大,有研究表明,施肥措施能夠顯著影響農(nóng)田土壤酶活性變化[37-39]。本研究結(jié)果表明,與對(duì)照不施肥處理相比,各施肥處理均能夠提高土壤全土酶和團(tuán)聚體脲酶、堿性磷酸酶、過氧化氫酶、蔗糖酶活性。其中,生物有機(jī)肥處理明顯高于化肥或緩控釋肥單施處理。分析其原因可能是外源有機(jī)物的攝入,為土壤微生物生命代謝活動(dòng)提供豐富的營養(yǎng)物質(zhì)和益生菌,且有機(jī)物的攝入有利于提高土壤孔隙度,降低土壤容重,能夠改善土壤微生態(tài)環(huán)境,提高微生物代謝水平,促進(jìn)根系對(duì)土壤養(yǎng)分的吸收與利用,從而提高土壤酶活性以及團(tuán)聚體酶活性[40]。本研究中,與對(duì)照不施肥或單施化肥/生物有機(jī)肥/緩控施肥處理相比,化肥、緩控釋肥與生物有機(jī)肥配施能夠提高>2 mm、>1~2 mm粒級(jí)團(tuán)聚體脲酶、堿性磷酸酶、蔗糖酶活性貢獻(xiàn)率。分析其原因可能是較大粒級(jí)團(tuán)聚體含有較多的有機(jī)碳,為酶促反應(yīng)提供更多的載體,使得大粒級(jí)團(tuán)聚體的酶促反應(yīng)效率高于小粒級(jí)團(tuán)聚體[19]。土壤不同粒級(jí)團(tuán)聚體酶活性的差異,表明團(tuán)聚體粒級(jí)對(duì)土壤酶活性的貢獻(xiàn)率與其組分有較大關(guān)系。而不同施肥處理?xiàng)l件下土壤酶活性與土壤容重、孔隙度及團(tuán)聚體穩(wěn)定性的相關(guān)分析表明,土壤酶活性的變化與土壤物理結(jié)構(gòu)及團(tuán)聚體穩(wěn)定性密切相關(guān)。說明合理施肥措施能夠通過改善土壤物理結(jié)構(gòu)與團(tuán)聚體組成,提升團(tuán)聚體穩(wěn)定性來促進(jìn)土壤酶活性的提高。 經(jīng)過3年定位試驗(yàn)發(fā)現(xiàn),與對(duì)照不施肥處理相比,化肥、緩控釋肥與生物有機(jī)肥配施能夠提高 >2 mm、>1~2 mm粒級(jí)團(tuán)聚體質(zhì)量分?jǐn)?shù)和團(tuán)聚體GMD、MWD和R>0.25值,降低D值;能夠提高土壤孔隙度,降低土壤容重;能夠提高全土酶以及團(tuán)聚體酶活性和>2 mm、>1~2 mm粒級(jí)團(tuán)聚體脲酶、堿性磷酸酶、蔗糖酶活性貢獻(xiàn)率。綜合土壤團(tuán)聚體、容重、孔隙度及酶活性方面結(jié)果,50%化肥配施50%生物有機(jī)肥處理整體表現(xiàn)優(yōu)于50%緩控釋肥配施50%生物有機(jī)肥處理。 參考文獻(xiàn): [1]馬仁罡,孫健英,李宗蕓. 基于生物信息學(xué)的甘薯基因組學(xué)等研究進(jìn)展[J]. 江蘇農(nóng)業(yè)學(xué)報(bào),2021,37(2):531-538. [2]劉亞軍,胡啟國,儲(chǔ)鳳麗,等. 不同栽培模式對(duì)甘薯干物質(zhì)分配及產(chǎn)量的影響[J]. 黑龍江八一農(nóng)墾大學(xué)學(xué)報(bào),2018,30(6):1-7,18. [3]王 欣,李 強(qiáng),曹清河,等. 中國甘薯產(chǎn)業(yè)和種業(yè)發(fā)展現(xiàn)狀與未來展望[J]. 中國農(nóng)業(yè)科學(xué),2021,54(3):483-492. [4]李 強(qiáng),趙 海,靳艷玲,等. 中國甘薯產(chǎn)業(yè)助力國家糧食安全的分析與展望[J]. 江蘇農(nóng)業(yè)學(xué)報(bào),2022,38(6):1484-1491. [5]蘭孟焦,張 輝,肖滿秋,等. 不同施肥處理對(duì)甘薯產(chǎn)量及土壤肥力與氮素利用的影響[J]. 西北農(nóng)業(yè)學(xué)報(bào),2023,32(1):44-52. [6]張偉彬. 有機(jī)肥和化肥配施對(duì)小麥甘薯輪作土壤腐殖質(zhì)結(jié)合形態(tài)及微生物群落結(jié)構(gòu)的影響[J]. 江蘇農(nóng)業(yè)科學(xué),2022,50(17):247-252. [7]段文學(xué),張海燕,解備濤,等. 化肥和生物有機(jī)肥配施對(duì)鮮食型甘薯塊根產(chǎn)量、品質(zhì)及土壤肥力的影響[J]. 植物營養(yǎng)與肥料學(xué)報(bào),2021,27(11):1971-1980. [8]劉亞軍,王文靜,李 敏,等. 無機(jī)肥與有機(jī)肥配施對(duì)甘薯土壤養(yǎng)分變化及微生物碳代謝功能的影響[J]. 河南農(nóng)業(yè)科學(xué),2022,51(7):75-84. [9]王 佳,嚴(yán) 偉,紀(jì) 要,等. 甘薯施肥施藥機(jī)械研究現(xiàn)狀及展望[J]. 中國農(nóng)機(jī)化學(xué)報(bào),2021,42(9):68-76. [10]張 輝,張永春. 肥料對(duì)甘薯營養(yǎng)品質(zhì)影響的研究進(jìn)展[J]. 江蘇農(nóng)業(yè)科學(xué),2017,45(17):1-5. [11]朱長偉. 輪耕模式對(duì)豫北潮土土壤肥力及作物產(chǎn)量的影響[D]. 鄭州:河南農(nóng)業(yè)大學(xué),2021. [12]秦俊梅,李嘉欣,宋建麗,等. 不同修復(fù)基質(zhì)對(duì)外源四環(huán)素污染土壤酶活性及微生物量碳氮的影響[J]. 應(yīng)用與環(huán)境生物學(xué)報(bào),2021,27(6):1500-1508. [13]吳強(qiáng)建,胡夢(mèng)蝶,侯松峰,等. 減氮配施生物炭基肥對(duì)蜜柚土壤理化性質(zhì)及酶活性的影響[J]. 河南農(nóng)業(yè)大學(xué)學(xué)報(bào),2022,56(5):732-741. [14]劉寒雙,崔紀(jì)菡,劉 猛,等. 有機(jī)肥替代部分化肥對(duì)谷子產(chǎn)量、土壤養(yǎng)分及酶活性的影響[J]. 中國土壤與肥料,2022(7):71-81. [15]楊文飛,杜小鳳,顧大路,等. 長期施肥對(duì)根系及土壤微生態(tài)環(huán)境、養(yǎng)分和結(jié)構(gòu)的影響綜述[J]. 江西農(nóng)業(yè)學(xué)報(bào),2020,32(12):37-44. [16]甘雅芬,徐永昊,周富忠,等. 紫云英還田與氮肥減施對(duì)水稻土團(tuán)聚體中各形態(tài)鐵錳含量的影響[J]. 植物營養(yǎng)與肥料學(xué)報(bào),2022,28(7):1238-1248. [17]聶 鑫,魯艷紅,廖育林,等. 化肥減施下紫云英不同翻壓量對(duì)水穩(wěn)性團(tuán)聚體及雙季稻產(chǎn)量的影響[J]. 華北農(nóng)學(xué)報(bào),2020,35(6):155-164. [18]張 帥,成宇陽,吳 行,等. 生物炭施用下潮土團(tuán)聚體微生物量碳氮和酶活性的分布特征[J]. 植物營養(yǎng)與肥料學(xué)報(bào),2021,27(3):369-379. [19]李委濤,李忠佩,劉 明,等. 秸稈還田對(duì)瘠薄紅壤水稻土團(tuán)聚體內(nèi)酶活性及養(yǎng)分分布的影響[J]. 中國農(nóng)業(yè)科學(xué),2016,49(20):3886-3895. [20]李 敏,劉亞軍,王文靜,等. 不同施肥模式對(duì)小麥—甘薯輪作田土壤性質(zhì)及甘薯生長發(fā)育的影響[J]. 江蘇農(nóng)業(yè)科學(xué),2022,50(13):250-256. [21]劉拴成. 有機(jī)肥與無機(jī)肥配施對(duì)馬鈴薯生長發(fā)育及產(chǎn)質(zhì)量的影響[J]. 河南農(nóng)業(yè)科學(xué),2020,49(3):32-39. [22]韓正砥,周明耀,楊雅琴,等. 生物有機(jī)肥配施對(duì)節(jié)水灌溉稻田養(yǎng)分累積及水稻生長的影響[J]. 水資源與水工程學(xué)報(bào),2022,33(4):210-216. [23]王 靜,王 磊,劉耀斌,等. 長期施用不同有機(jī)肥對(duì)甘薯產(chǎn)量和土壤生物性狀的影響[J]. 水土保持學(xué)報(bào),2021,35(2):184-192. [24]趙 營,劉曉彤,羅健航,等. 緩/控釋肥條施對(duì)春玉米產(chǎn)量、吸氮量與氮平衡的影響[J]. 中國土壤與肥料,2020(5):34-39. [25]關(guān)松蔭. 土壤酶及其研究法[M]. 北京:農(nóng)業(yè)出版社,1986:64-126. [26]劉小粉,劉春增,賈萌萌,等. 化肥減施條件下紫云英翻壓量對(duì)土壤物理特性和水稻產(chǎn)量的影響[J]. 江蘇農(nóng)業(yè)科學(xué),2021,49(16):67-73. [27]張玉銘,胡春勝,陳素英,等. 耕作與秸稈還田方式對(duì)碳氮在土壤團(tuán)聚體中分布的影響[J]. 中國生態(tài)農(nóng)業(yè)學(xué)報(bào),2021,29(9):1558-1570. [28]孫 雪,張玉銘,張麗娟,等. 長期添加外源有機(jī)物料對(duì)華北農(nóng)田土壤團(tuán)聚體有機(jī)碳組分的影響[J]. 中國生態(tài)農(nóng)業(yè)學(xué)報(bào),2021,29(8):1384-1396. [29]隋夕然,吳麗芳,王 妍,等. 滇中巖溶高原不同石漠化程度土壤團(tuán)聚體養(yǎng)分及酶活性特征[J]. 浙江農(nóng)林大學(xué)學(xué)報(bào),2022,39(1):115-126. [30]林洪羽,周明華,張博文,等. 生物炭及秸稈長期施用對(duì)紫色土坡耕地土壤團(tuán)聚體有機(jī)碳的影響[J]. 中國生態(tài)農(nóng)業(yè)學(xué)報(bào),2020,28(1):96-103. [31]劉希玉,王忠強(qiáng),張心昱,等. 施肥對(duì)紅壤水稻土團(tuán)聚體分布及其碳氮含量的影響[J]. 生態(tài)學(xué)報(bào),2013,33(16):4949-4955. [32]葉麗麗,王翠紅,周 虎,等. 添加生物質(zhì)黑炭對(duì)紅壤結(jié)構(gòu)穩(wěn)定性的影響[J]. 土壤,2012,44(1):62-66. [33]李 銳,陶 瑞,王 丹,等. 減氮配施有機(jī)肥對(duì)滴灌棉田土壤生物學(xué)性狀與團(tuán)聚體特性的影響[J]. 應(yīng)用生態(tài)學(xué)報(bào),2017,28(10):3297-3304. [34]席凱鵬,楊蘇龍,席吉龍,等. 長期棉花秸稈配施有機(jī)肥對(duì)土壤理化性質(zhì)及棉花產(chǎn)量的影響[J]. 中國土壤與肥料,2022(7):82-90. [35]王西和,楊金鈺,王彥平,等. 長期施肥措施下灰漠土有機(jī)碳及團(tuán)聚體穩(wěn)定性特征[J]. 中國土壤與肥料,2021(6):1-8. [36]王 威,吳大付,唐 蛟,等. 秸稈全量還田下沼液替代化肥對(duì)潮土團(tuán)聚體及結(jié)合有機(jī)碳的影響[J]. 土壤通報(bào),2022,53(4):847-857. [37]胡慶蘭,楊 凱,王金貴,等. 地膜覆蓋及不同施肥處理對(duì)玉米產(chǎn)量和根際土壤酶活性的影響[J]. 江蘇農(nóng)業(yè)科學(xué),2022,50(21):122-126. [38]周東興,李 磊,李 晶,等. 玉米/大豆輪作下不同施肥處理對(duì)土壤微生物生物量及酶活性的影響[J]. 生態(tài)學(xué)雜志,2018,37(6):1856-1864. [39]胡哲偉,金 淑,應(yīng)蓉蓉,等. 蚓糞和益生菌配施對(duì)土壤微生物生物量及酶活性的影響[J]. 江蘇農(nóng)業(yè)科學(xué),2021,49(11):201-207. [40]李 娜,韓曉增,尤孟陽,等. 土壤團(tuán)聚體與微生物相互作用研究[J]. 生態(tài)環(huán)境學(xué)報(bào),2013,22(9):1625-1632.
