玉米小分子肽在賴氨酸發酵中應用探究及工藝優化
2023-08-17 21:11:59王金多李瀾瀟徐慶陽
中國調味品 2023年8期
王金多 李瀾瀟 徐慶陽

摘要:為了降低賴氨酸發酵生產過程中“三廢”的排放,減少成分復雜的玉米漿對發酵液的影響,提高分離提取良品率,該研究優化了L-賴氨酸生產菌谷氨酸棒桿菌LS260的發酵條件,通過氮源分析,采用近似等效替代法,使用玉米小分子肽替代玉米漿,通過5 L生物反應器發酵試驗探究清潔發酵工藝的可行性,通過單因素試驗、響應面優化試驗進一步優化了培養基營養成分,結合營養適量流加發酵控制工藝,確定了最適合LS260的發酵條件。結果顯示,通過氮源等效替代試驗,賴氨酸產量為124 g/L,轉化率提高至71.5%;通過營養適量流加工藝將產量提高至136 g/L,發酵上清液透光率提高至84.4%;由單因素試驗、響應面法優化試驗可知,生物素、KH2PO4的最適添加量分別為3.2 mg/L、4.11 g/L,最適接種量為25.49%。在試驗所得的最適條件下,賴氨酸產量達到160.3 g/L,轉化率達到71.5%。結果顯示,最終產量和轉化率與模型數值基本相符,發酵液上清液透明度大幅降低,為提取工藝進一步優化創造了條件,試驗結果具有較好的工業應用前景。
關鍵詞:L-賴氨酸;玉米小分子肽;清潔發酵;谷氨酸棒桿菌;響應面優化
中圖分類號:TS211.2????? 文獻標志碼:A???? 文章編號:1000-9973(2023)08-0018-06
Application and Process Optimization of Maize Small-Molecule Peptides in Lysine Fermentation
WANG Jin-duo1,2,3, LI Lan-xiao1,2,3, XU Qing-yang1,2,3*
(1.College of Bioengineering, Tianjin University of Science and Technology, Tianjin 300457, China;
2.National and Local United Engineering Laboratory of Metabolic Control Fermentation
Technology, Tianjin 300457, China; 3.Tianjin Engineering Laboratory of Efficient
and Green Amino Acid Manufacturing, Tianjin 300457, China)
Abstract: In order to reduce the emission of “three wastes” in the lysine fermentation production, reduce the effect of corn pulp with complex components on the fermentation broth, and improve the separation and extraction rate, in this study, the fermentation conditions of L-lysine producing bacterium Corynebacterium glutamicum LS260 is optimized. Through nitrogen source analysis, the approximate equivalent substitution method is adopted to replace corn pulp with maize small-molecule peptides, and the feasibility of clean fermentation process is explored through 5 L bioreactor fermentation test. The nutrients of the medium are further optimized by single factor test and response surface optimization test, and the optimal fermentation conditions for LS260 are determined by combining the nutrient-appropriate flow-adding fermentation control process. The results show that through the nitrogen source equivalent substitution test, the yield of lysine is 124 g/L and the conversion rate increases to 71.5%. The yield increases to 136 g/L and the light transmittance of fermentation supernatant increases to 84.4% by nutrient-appropriate flow-adding process. According to single factor test and response surface method optimization test, the optimal addition amount of biotin and KH2PO4 is 3.2 mg/L and 4.11 g/L respectively, and the optimal inoculation amount is 25.49%. Under the optimal conditions, the yield of lysine reaches 160.3 g/L and the conversion rate reaches 71.5%. The results show that the final yield and conversion rate are basically consistent with the model values, and the transparency of fermentation broth supernatant is greatly reduced, which has created conditions for further optimization of the extraction process. The test results have a good prospect for industrial application.
Key words: L-lysine; maize small-molecule peptide; clean fermentation; Corynebacterium glutamicum; response surface optimization
收稿日期:2023-02-24
基金項目:天津市合成生物技術創新能力提升行動計劃項目(TSBICIP-KJGG-005-08);山東省重點研發計劃(2021ZDSYS10);天津市科技計劃項目(21ZYQCSY00050)
作者簡介:王金多(1997-),男,碩士,研究方向:代謝工程與發酵過程控制。
通信作者:徐慶陽(1980-),男,研究員,博士生導師,博士,研究方向:代謝工程與發酵過程控制。
L-賴氨酸作為人和動物的八大必需氨基酸之一,在醫療、畜牧業及食品工業中得到了廣泛的應用[1]。在飼料中添加賴氨酸,能夠促進牲畜的生長速度,改善飼料中氨基酸平衡,提高飼料和蛋白質資源的利用效率,提高出欄率并減少飼養成本。飼料中添加適量的賴氨酸能夠提高出欄時牲畜的肉質及瘦肉率,亦可增強經濟動物的抵抗力。由于分解法和化學合成法制得的賴氨酸很難達到獸用標準,不能直接產出,現多通過發酵法生產賴氨酸,但在發酵過程中多采用營養豐富但成分復雜的農業副產物,如:玉米漿、大豆蛋白水解液、糖蜜等,但受季節、產地、作物品種等變化導致的發酵培養基成分波動在一定程度上影響菌體的生長和產酸[2],且在產物分離提取過程中,由于農業副產物中常含有大量色素、雜蛋白,造成提取過程中耗水、耗電;產生的固形廢物、含鹽廢水亦會對周圍土地及水環境產生惡劣影響。賴氨酸的發酵過程屬于典型的高密度發酵,菌體在積累賴氨酸的過程中受到多個關鍵酶的共同調控[3]。在分析玉米漿、糖蜜、大豆蛋白水解液主要組分后,探究使用成分穩定且環境友好的玉米小分子肽做近似等效替代,探究清液發酵在賴氨酸生產方面的可行性。
1 材料與方法
1.1 菌種
谷氨酸棒桿菌LS260:保藏于天津科技大學代謝工程研究室。
1.2 培養
1.2.1 種子培養基
葡萄糖80 g/L,玉米漿干粉20 g/L,(NH4)2SO4·7H2O 6 g/L,KH2PO4 1.7 g/L,MgSO4 0.6 g/L,VB1 1.5 mg/L,VB5 1.5 mg/L,VB12 1.5 mg/L,VH 2 mg/L。
1.2.2 對照發酵培養基
葡萄糖60 g/L,玉米漿干粉25 g/L,(NH4)2SO4·7H2O 5 g/L,KH2PO4 4 g/L,MgSO4 1 g/L,VB1 1 mg/L,VB5 1 mg/L,VB12 1 mg/L,VH 1.5 mg/L。
1.2.3 清液發酵培養基
葡萄糖60 g/L,玉米小分子肽25 g/L,(NH4)2SO4·7H2O 5 g/L,KH2PO4 4 g/L,MgSO4 1 g/L,VB1 1 mg/L,VB5 1 mg/L,VB12 1 mg/L,VH 1.5 mg/L,FeSO4·7H2O 50 mg/L,MnSO4·H2O 110 mg/L,CuSO4·5H2O 2 mg/L,ZnSO4 10 mg/L,PLP 5 mg/L。
1.2.4 流加培養基
玉米小分子肽7 g/L,VB1、VB3、VB5、VB12 分別為2.5 mg/L,VH 2 mg/L,甜菜堿 3 g/L,氯化膽堿 4 g/L。
1.2.5 斜面培養基
葡萄糖1 g/L,酵母粉 5 g/L,蛋白胨 10 g/L,(NH4)2SO4·7H2O 5 g/L,KH2PO4 4 g/L,MgSO4 1 g/L。
1.3 主要儀器
儀器與設備見表1。
1.4 培養方法
斜面培養:一代試管斜面活化2支,恒溫箱32 ℃培養24 h,二代茄形瓶斜面活化2支,恒溫箱32 ℃培養18 h。
種子罐培養:接種量2支茄形瓶,發酵體積2 L,培養溫度34 ℃,pH 7.0~7.2,溶氧45%以上,轉速與溶氧聯動,種子培養12~15 h。
發酵罐培養:接種量600 mL,發酵體積3 L,培養溫度34 ℃,pH 7.0~7.2,溶氧30%以上,轉速與溶氧聯動,發酵時間36 h。
1.5 檢測方法
1.5.1 pH值測定
梅特勒pH在線檢測,精密pH試紙輔助檢測。
1.5.2 菌體生物量測定
菌體生物量采用OD600 nm吸光度法測定,發酵液用容量瓶稀釋特定倍數后,用可見光分光光度計測定吸光度,吸光度×稀釋倍數得到實際OD值,可以直接表示菌體生物量[4]。
1.5.3 L-賴氨酸含量測定
發酵液中L-賴氨酸及副產物含量采用高效液相色譜法測定,采用Agilent C18色譜柱(15 mm×4.6 mm,3.5 μm),衍生劑為2,4-二硝基氟苯,柱前衍生[5],流動相為50%的乙腈、4.1 g/L的醋酸鈉溶液,柱溫33 ℃,流速1 mL/min,檢測波長360 nm。
1.5.4 蛋白質含量測定
采用考馬斯亮藍法測定。
1.6 糖酸轉化率計算方法
糖酸轉化率SA計算公式:
SA(%)=ρ×Vm×100%。
式中:ρ為L-賴氨酸質量濃度,g/L;V為發酵液總體積,L;m為總耗糖量,g。
所有試驗數據取3次試驗的平均值。單因素方差分析后采用Dunnett-t檢驗來確定數據差異的顯著性(P<0.05)[6]。
1.7 響應面試驗設計
Box-Behnken中心組合試驗:采用Box-Behnken中心組合試驗進行試驗條件優化,以生物素添加量(A)、KH2PO4添加量(B)、接種量(C)為自變量,以L-賴氨酸產量為響應值,通過響應面試驗進行發酵條件優化。
1.8 單因素試驗設計
單因素試驗設計見表2。
2 結果與討論
2.1 玉米漿與玉米小分子肽滅菌后蛋白含量對比分析
玉米漿作為賴氨酸發酵過程中最重要的氮源之一,對菌體生長起著決定性作用。而玉米漿中的雜蛋白是影響發酵過程中菌體代謝和后提取過程中離子交換樹脂壽命的重要因素,雜蛋白的含量越高,賴氨酸成品的良品率越低。所以,選擇成分穩定、蛋白含量低、色素含量少的氮源非常重要。
因生物來源類似,選取玉米小分子肽作為賴氨酸發酵中玉米漿氮源的替代物質。為了對比玉米小分子肽和玉米漿的蛋白質含量,通過凱氏定氮法測定蛋白質含量,結果見圖1。
經過水解處理后的玉米小分子肽蛋白質含量更少。一次滅菌后,玉米漿的蛋白質含量比玉米小分子肽高129%。玉米漿雜蛋白含量過高,在發酵過程中菌體要代償性分解蛋白質以獲取生長必需的氮源[7],剩余的蛋白質在賴氨酸提取過程中容易導致產品質量不合格,雜質超標,故玉米小分子肽作為發酵原料效果更優。
2.2 玉米漿與玉米小分子肽滅菌后不同氨基酸含量對比分析
賴氨酸發酵是典型的生長偶聯型,相對于成分復雜且需要主動分解的蛋白質而言,單分子氨基酸更容易被菌體利用,發酵接種后更容易適應發酵培養基,延滯期更短,對數期生長速率更快,產酸速率更快,能夠有效提高菌體活力。
通過氨基酸分析儀對比玉米漿和玉米小分子肽中各種游離氨基酸含量,結果見圖2。
通過氨基酸分析儀對比分析,玉米漿與玉米小分子肽的氨基酸含量有較大區別[8-9],玉米小分子肽大部分氨基酸含量比玉米漿高,主要是天冬氨酸高41.7%,蘇氨酸高7.3%,絲氨酸高58.4%,甘氨酸高28.6%,苯丙氨酸高69.3%,組氨酸高21.7%,丙氨酸高23.7%,纈氨酸高6.3%,異亮氨酸高8.6%,亮氨酸高11.5%;玉米小分子肽中的谷氨酸、甲硫氨酸、半胱氨酸、賴氨酸相較于玉米漿含量稍低。
2.3 清潔發酵培養基與傳統培養基發酵對比探究
為了確定玉米小分子肽在賴氨酸發酵過程中的效果和對主要發酵參數的影響,從菌體量、產酸量、轉化率、色素含量幾個角度與傳統發酵培養基進行對比,從而確定進一步優化的可行性。
以傳統發酵培養基為對照組,清潔發酵培養基為試驗組。由圖3可知,從菌體的生長曲線看,采用傳統發酵培養基的對照組的生長速率和最高菌體量明顯高于試驗組,對照組的最高OD600 nm為141.6,相較試驗組的107.2高24.29%,在生長穩定期后,試驗組的生物量下降速率比對照組高50%;L-賴氨酸的產量與菌體生長曲線走勢相似,產酸速率隨著菌體生長速率的變化而變化,發酵前期,試驗組的生長速率明顯高于對照組,發酵10 h后,對照組的生長速率明顯高于試驗組,對照組最終產量為159 g/L,試驗組最終產量為124 g/L,相較于對照組下降22.01%。對照組培養基天然成分豐富,且培養基中維生素[10]、微量元素、生長因子等高于試驗組[11]。從產酸曲線可以看出,在發酵前期,試驗組的產酸速率明顯高于對照組,且整體產酸速率相對穩定,說明發酵接種后,菌體能更好、更快地適應組分穩定、均一清潔培養基。賴氨酸的發酵過程是嚴格的生長偶聯型,且生長速率的下降勢必引起產酸速率下降,發酵前期,試驗組的生長速率與對照組相比相差無幾,但在中后期的生長維持上低于對照組,所以解決試驗組發酵中后期的菌體活力維持是主要問題。
由表3可知,試驗組的糖酸轉化率較對照組高3.6%;試驗組發酵上清液透光率較對照組提高了25.6%。盡管選用清潔發酵培養基的產量不及傳統培養基,但是糖酸轉化率有一定的提高,考慮原因可能是用于菌體生長的碳代謝流向產賴氨酸;對照組發酵培養基中含有較多的生物素,生物素的添加量與菌體的合成速度及厚度呈正相關。試驗組發酵培養基中生物素含量較低,有利于賴氨酸的外排,減小胞內反饋抑制,促進碳代謝流向有益于賴氨酸合成方向。試驗組發酵上清液透光率較對照組提高了25.6%,后提取過程中,活性炭和水的用量可大大降低。
2.4 營養流加對賴氨酸清潔發酵的影響
為解決使用玉米小分子肽進行賴氨酸發酵菌體量不足、發酵后期菌體活力不足的問題,采用營養流加的策略進行發酵優化。試驗組從發酵10 h開始進行營養流加,流加速度為100 mL/h,試驗結果見圖4。
由圖4可知,以清潔發酵培養基為對照組,清潔發酵培養基配合營養流加為試驗組。試驗組在2.3的基礎上,于發酵10 h開始流加1.2.4中的流加培養基,以解決發酵中后期營養不足引起的菌體生長停滯及活力下降問題。在發酵0~10 h,試驗組與對照組菌體的生長速率、產酸水平近似相同,開始流加后,試驗組最高OD600 nm為129,試驗組的最終產量為136 g/L,產量較對照組提高9.68%。菌體生長對數期延長約4 h左右,在14 h后,菌體處于低速生長的穩定期,雖產酸速率有所降低,但整體的產酸速率和產量相較于對照組有大幅提高。
2.5 賴氨酸清潔發酵工藝優化單因素試驗
影響谷氨酸棒桿菌發酵生產賴氨酸過程中,菌體的生長周期及生長狀態對賴氨酸產量及產率的影響極為重要,除了碳源、氮源外,影響發酵過程中菌體生長周期及狀態的主要因素為生物素、KH2PO4的添加量和接種量[12]。
圖5中A、B、C、D、E 5個水平分別對應表3中1,2,3,4,5 5個水平。由圖5可知,添加適量的生物素有益于菌體生長,當生物素添加量過高時,可能導致細胞膜生長更致密,不利于賴氨酸外排。生物素添加量為3 mg/L時,賴氨酸產量最優,為127 g/L;KH2PO4能夠為菌體生長提供必需的鉀離子和磷酸根,對菌體滲透壓的維持及細胞膜的合成有重要作用,當磷酸鹽添加量為4 g/L時,賴氨酸產量達到126 g/L;接種量在發酵控制過程中是除溫度、pH、溶氧外最重要的控制參數之一[13-15],合適的接種量對發酵各階段的影響較顯著,是調節發酵各階段所需時間、縮短菌體延滯期、延長對數期的重要策略,對緩解菌體早衰、延長產酸期、提高糖酸轉化率有一定效果,試驗選取5個接種量水平,接種量25%為最優,此時賴氨酸產量為137 g/L。
2.6 發酵條件響應面試驗優化分析
在單因素試驗的基礎上,以生物素添加量(A)、KH2PO4添加量(B)、接種量(C)為自變量,以L-賴氨酸產量為響應值,通過響應面試驗進行發酵條件優化,響應面試驗設計及結果見表4。
以L-賴氨酸產量為響應值,生物素添加量(A)、KH2PO4添加量(B)、接種量(C)為自變量,建立回歸方程Y(賴氨酸)=157.40+5.63A+3.00B+2.63C-4.50AB-4.75AC+2.00BC-11.83A2-10.08B2-9.82C2。
由表5可知,此模型的P值<0.000 1,說明通過該回歸模型的設計所得試驗結果均影響極顯著,失擬項的P值=0.541 3>0.05,說明該回歸曲線的擬合度良好,具有較高的可信度。回歸方程的決定系數R2為0.984 271,回歸方程的校正決定系數RAdj2為0.964 048。由P值可知,該模型的一次項A、B,二次項A2、B2、C2,交互項AB、AC對結果的影響極顯著;一次項C對結果的影響顯著;交互項BC對結果的影響不顯著。由F值可以確定3個因素對賴氨酸產量的影響大小順序為生物素添加量>KH2PO4添加量>接種量。測量信噪比為20.585,信號充足,表明該模型可以應用于試驗設計。
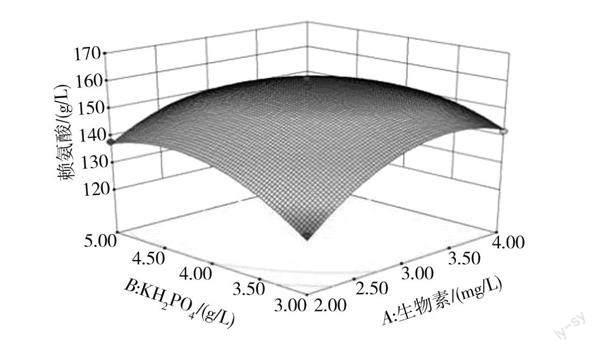
各因素交互作用對賴氨酸產量影響的響應面及等高線見圖6。
由圖6可知,賴氨酸的產量隨著各自變量數值的不斷增加呈現先升高后降低的趨勢,說明兩因素間存在一定交互作用,其中AB曲面彎曲程度最大,等高線更接近橢圓形,說明AB交互作用的影響極顯著。
通過響應面軟件對方程求解,得到最佳發酵條件為生物素添加量(A)3.20 mg/L、KH2PO4添加量(B)4.11 g/L、接種量(C)25.49%。結合分批補料發酵操作過程中的特殊性,接種量調整為25%。在此發酵條件下,賴氨酸的實際產量為160.3 g/L,與優化所得理論值相近,說明該方案具有可行性。
3 結論
通過不同碳源的成分對比分析,采用近似等效替代的方法得到一種效果相近的清潔發酵培養基,通過5 L生物反應器發酵試驗探究清潔發酵培養基的可行性,最終產量為124 g/L,轉化率提高至71.5%;通過營養流加進一步提高產量至136 g/L。
利用單因素試驗結合響應面優化對賴氨酸清潔發酵培養基中對菌體生長及產酸影響顯著的成分添加量進行優化,得到了最優發酵條件:生物素添加量3.20 mg/L、KH2PO4添加量4.11 g/L、接種量25.49%。結合營養流加工藝,賴氨酸的實際產量為160.3 g/L。
試驗得到的賴氨酸的清潔發酵方法在賴氨酸發酵工業有一定的應用前景,不僅能提高獸用賴氨酸的品質,主要優勢表現在后提取過程中能大量節省活性炭和水,而且能提高離子交換樹脂的使用壽命及成品的良品率,符合發酵工業的未來發展方向和“雙碳”發展理念。
參考文獻:
[1]XU J Z, WU Z H, GAO S J, et al. Rational modification of tricarboxylic acid cycle for improving L-lysine production in Corynebacterium glutamicum[J].Microbial Cell Factories,2018,17(1):105.
[2]劉景陽,余子辰,徐慶陽.NH4+雙階段發酵控制谷氨酸工藝研究[J].中國調味品,2022,47(9):7-14.
[3]Alpha-hydroxy-gamma-methylmercaptobutyric-acid/alpha-ketoisocaproic-acid/alpha-ketoisovalerate/calcium/histidine/lysine-hydrochloride/nitrogen/phenylpyruvic-acid/threonine/tryptophan/tyrosine[J].Reactions Weekly,2022,1922(1):22319-22327.
[4]NAGY A, MAGYAR T, JUHSZ C, et al. Phytoremediation of acid mine drainage using by-product of lysine fermentation[J].Water Science and Technology,2020,81(7):1507-1517.
[5]SUVACHAN A, LAL R, NAMPOOTHIRI K M, et al. Valorization of paper industry rejects by combined thermo-chemical pretreatment and biological conversion to L-lysine[J].Environmental Technology & Innovation,2021,24:101882.
[6]熊海波,陳志超,曹華杰,等.全營養流加對谷氨酸棒桿菌發酵產L-異亮氨酸的影響[J].食品與發酵工業,2021,47(6):11-17.
[7]李萬軍.優化L-賴氨酸發酵工藝提高糖酸轉化率[D].銀川:寧夏大學,2021.
[8]曹衛,徐陽鑫,賀婷,等.發酵食品中羧甲基賴氨酸的研究進展[J].中國調味品,2019,44(1):164-168.
[9]于玲紅,李巖,敬雙怡,等.特異性移動床生物膜反應器處理賴氨酸發酵廢水[J].環境污染與防治,2018,40(7):770-773.
[10]洪銘,李巖,張蘇龍,等.利用甜菜堿提升賴氨酸發酵及其代謝通量分析[J].發酵科技通訊,2019,48(3):125-131.
[11]KENTO T, YOSHIKI S, TAKAHIRO T, et al. Data science-based modeling of the lysine fermentation process[J].Journal of Bioscience and Bioengineering,2020,130(4):409-415.
[12]張東.等離子體誘變選育L-賴氨酸產生菌及發酵優化研究[D].鄭州:河南農業大學,2018.
[13]楊帆,王琳琳,時德通,等.響應面法優化賴氨酸發酵生產條件的研究[J].食品工業,2019,40(2):200-203.
[14]BASHIR S, BASHIR R, PERVAIZ M, et al. RSM-based optimization of fermentation conditions and kinetic studies of glutamic acid and lysine production by Corynebacterium glutamicum[J].Journal of Nanomaterials,2022,2022:1-6.
[15]XU Y Q, ZHOU D, LUO R S, et al.Metabolic engineering of Escherichia coli for polyamides monomer δ-valerolactam production from feedstock lysine[J].Applied Microbiology and Biotechnology,2020,104:10939-10946.
