循環水養殖系統中養殖密度對松江鱸生長的影響
2023-08-23 17:42:53高桂明季強黃迪
水產科技情報 2023年2期
關鍵詞:生長
高桂明 季強 黃迪

摘 要:為研究配合飼料條件下循環水養殖系統(RAS)中養殖密度對松江鱸生長的影響,選取體長為(2.97±0.12)cm、體質量為(0.26±0.03)g的松江鱸,分別按40尾/m2(A組)、80尾/m2(B組)和120尾/m2(C組)共3個養殖密度,在RAS中進行了為期240 d的養殖試驗。試驗結果顯示:A組魚的終末體質量、終末體長、體質量日增長量、存活率等均顯著高于其他兩組,A組魚的體長日增長量顯著高于C組(P<0.05);不同密度組間魚體肥滿度無顯著性差異(P>0.05)。試驗組單位面積產量由高到低依次為:C組(2.83 kg/m2)、B組(2.51 kg/m2)、A組(1.72 kg/m2)。試驗組魚體質量與體長均呈冪函數相關(m=aLb,a=0.007 6~0.008 9,b=3.123 6~3.209 4),體長、體質量生長均以三次函數擬合較好。各組間的魚體長、體質量變異系數均差異顯著(P<0.05),其中B組最小。試驗表明:在配合飼料條件下,隨著養殖密度的提高,RAS養殖的松江鱸生長性能逐漸降低,其合理的養殖密度為40~80尾/m2。在本試驗條件下,建議在養殖早期采用高密度養殖模式,同時加強養殖期間對水溫的控制和管理。
關鍵詞:松江鱸;循環水養殖系統;養殖密度;配合飼料;生長
松江鱸(Trachidermus fasciatus Heckel)又名四鰓鱸,隸屬于鲉形目(Scorpaeniformes)、杜父魚科(Cottidae)、松江鱸魚屬(Trachidermus),為肉食性降河洄游性魚類,是我國四大淡水名魚之一,具有悠久的歷史文化底蘊、較高的生態價值和經濟價值。自20世紀80年代起,在水體污染、氣候變暖和洄游受阻等多重影響下,松江鱸資源急劇衰退,并于1988年被正式列入《國家Ⅱ級水生野生保護動物名錄》[1]。為保護和恢復松江鱸種質資源,上海市松江區水產技術推廣站自2006年開始開展松江鱸人工繁育技術研究,于2013年取得松江鱸規模化繁育成功,2017年又攻克了松江鱸轉食馴化技術,突破性地解決了松江鱸只攝食鮮活餌料的難題,為松江鱸規模化、集約化人工養殖奠定了重要基礎。魚類經濟價值的開發離不開集約化的人工養殖。循環水養殖系統(recirculating aquaculture system,RAS)具有穩定可控、資源節約、環境友好等優點,符合水產養殖業綠色發展的時代要求,是我國未來水產養殖發展的重要模式。目前,有關工廠化循環水養殖松江鱸的相關研究只見于零星報道[2],因此,開展松江鱸工廠化循環水養殖的相關研究對其規模化、集約化養殖有著重要的實踐意義。
高密度養殖是RAS的主要特點,也是決定養殖對象生長性能和養殖效益的重要環境因子之一。由于RAS對水質、溶解氧等環境因子具有較強的可控性,且不同的養殖魚類對養殖密度的適應性并不相同,在實際養殖生產過程中,需要選擇合理的養殖密度,以保障在安全養殖條件下產量和養殖效益最大化。諸多研究表明,不合理的養殖密度,會對魚類造成慢性擁擠脅迫,從而影響魚類的生長、免疫等指標[3-5]。為此,本研究基于投喂配合飼料的條件下比較了循環水養殖系統中養殖密度對松江鱸生長的影響,以期為提高松江鱸RAS養殖技術提供參考,為松江鱸資源保護和恢復提供重要的技術支持。
1 材料和方法
1.1 試驗材料
1.1.1 試驗環境
試驗于2021年在上海市松江區水產良種場循環水養殖車間內進行。試驗池為9個玻璃鋼桶(直徑4 m,高0.8 m),水深65~70 cm,池水流速0.5 cm/s,均配備獨立的循環過濾凈化系統和紫外線殺菌設備。養殖系統消毒、清洗、排干后待用。試驗魚入池前15 d,養殖池進水進行預運轉。養殖用水為經消毒、沉淀、曝氣后的井水。
1.1.2 試驗用魚
試驗用魚為上海市松江區水產良種場同批次繁育的松江鱸幼魚,經淡化、轉食馴化成功后,按同一標準從同一苗種培育池挑選出規格整齊、體質健壯、體色正常的個體。試驗魚體長為(2.97±0.12)cm,體質量為(0.26±0.03)g。
1.1.3 飼料營養成分
試驗用飼料為寧波天邦飼料科技有限公司生產的“邦尼”系列全熟化緩降微囊飼料。養殖前期(1~90 d)、中期(90~150 d)、后期(150~240 d)飼料的營養組成(質量分數)分別為:粗蛋白質≥52%,粗脂肪≥10%,粗纖維≤3%,粗灰分≤16%,水分≤10%,總磷≥1.5%,賴氨酸≥2.8%;粗蛋白質≥48%,粗脂肪≥6%,粗纖維≤3%,粗灰分≤16%,水分≤10%,總磷≥0.6%,賴氨酸≥2.6%;粗蛋白質≥45%,粗脂肪≥8%,粗纖維≤5%,粗灰分≤18%,水分≤12%,總磷≥1.0%,賴氨酸≥2.4%。
1.2 試驗方法
1.2.1 試驗設計及魚種放養
試驗設置3個密度梯度,分別為40尾/m2(A組)、80尾/m2(B組)和120尾/m2(C組),每個養殖密度設3個平行。篩選和放養試驗魚時操作要小心細致,以防止幼魚擦傷,減少其應激反應。放養前后,養殖水體潑灑抗應激藥物(“拜激靈”,拜耳動物保健有限公司)各1次。
1.2.2 飼料投喂
每天投喂2次(9:00和15:00),投飼量視魚的攝食情況靈活掌握,以70%~80%魚吃飽為原則,并根據魚體規格合理調整飼料粒徑。
1.2.3 日常管理
試驗期間,保證車間內環境安靜,避免外界干擾。試驗期間全程開啟循環過濾凈化系統,在夜間增開紫外線滅菌設備。每天(8:00、16:00)測量2次水溫,養殖期間水溫在8.4~26.6 ℃。定期檢測水質,試驗期間將水質指標控制在:透明度>60 cm,氨氮<1.0 mg/L,亞硝酸鹽<0.1 mg/L,溶解氧>5 mg/L,pH 7.5~8.5。
試驗自4月25日開始至12月21日結束,歷時240 d。
1.2.4 樣品采集
試驗期間,每30 d取樣1次,取樣時間為上午投飼前。從每個平行隨機取樣20尾,測定其體長與體質量。計算各組的特定生長率(RSG,%/d)、體長日增長量(LDG,cm/d)、體質量日增長量(MDG,g/d)、肥滿度(K,g/cm3)、變異系數(CV,%)和存活率(RS,%)。各指標計算公式如下。
RSG=(lnm2-lnm1)/(t2-t1)×100 (1)
LDG=(L2-L1)/(t2-t1)×100(2)
MDG=(m2-m1)/(t2-t1)×100 (3)
K=m/L3×100(4)
CV=S/X×100(5)
RS=Nt/N0×100 (6)
式(1)~(6)中,m1和m2為t1、t2時所對應的體質量(g),L1和L2為t1、t2時所對應的體長(cm),m為體質量(g),L為體長(cm),S為標準差,X為平均值,N0和Nt分別為試驗魚在試驗開始和結束時的數量(尾)。
1.3 數據處理
試驗數據用EXCEL 2010軟件運算并繪圖,用SPSS 19.0軟件進行分析處理,用單因素方差分析和Duncans多重比較法進行顯著性檢驗,設P<0.05為差異顯著。試驗結果用(平均值±標準差)表示。
2 結果
2.1 不同養殖密度下松江鱸的生長情況
從表1可見,經過240 d的養殖,松江鱸的大部分生長指標呈現隨養殖密度的增加而降低的趨勢。其中,A組試驗魚的終末體質量、終末體長、體質量日增長量和存活率顯著高于B、C組,且各組間差異顯著(P<0.05);A組試驗魚的體長日增長量顯著高于C組(P<0.05),B組試驗魚的體長日增長量與A、C組差異均不顯著(P>0.05);各組間的特定生長率和肥滿度無顯著性差異(P>0.05)。A、B、C組的單位面積產量分別為1.72、2.51、2.83 kg/m2。
2.2 不同養殖密度下松江鱸的生長式型
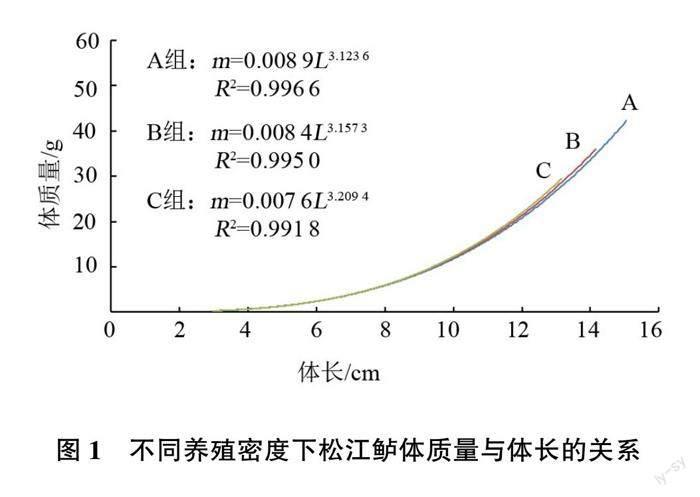
對各組松江鱸的體長、體質量數據用體長和體質量生長方程關系式(m=aLb)進行擬合,結果見圖1。由圖1可以看出,各組松江鱸的體長與體質量均呈良好的冪函數關系。A組、B組和C組的b值分別為3.123 6、3.157 3和3.209 4,均大于3,表明在投喂配合飼料的條件下循環水系統養殖的松江鱸為異速生長類型,魚體質量增長略快于體長增長。
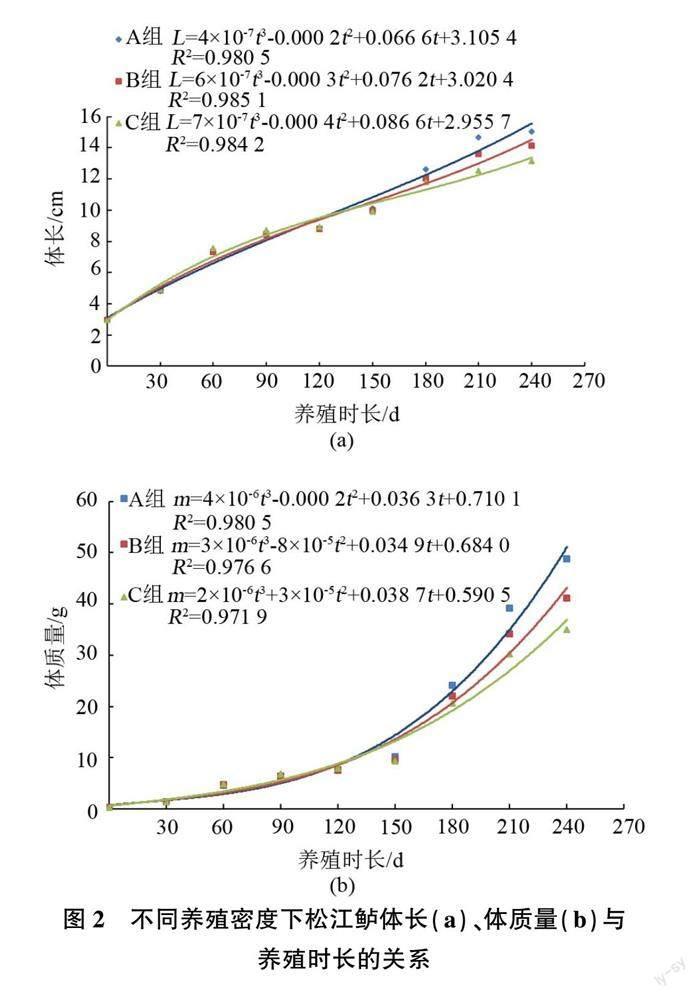
由圖2可見,不同養殖密度下養殖的松江鱸,其體長、體質量與養殖時長呈三次函數關系。
2.3 不同養殖密度下松江鱸的特定生長率
由圖3可見,各試驗組魚的特定生長率在試驗開始后的前120 d,隨著水溫的上升而降低,并在第120天時降到最小值,之后,隨著水溫的逐漸下降呈現先升高后降低的趨勢。
在0~90 d,各組特定生長率由高到低依次為:C組>B組>A組,各組間無顯著性差異(P>0.05);在91~240 d,各組特定生長率由高到低依次為:A組>B組>C組,各組間差異顯著(P<0.05)。
2.4 不同養殖密度下松江鱸的生長離散情況
由表2可見,B組的魚體長與體質量變異系數最小,說明B組松江鱸生長離散小,規格較均勻。各組間體長變異系數和體質量變異系數均存在顯著差異(P<0.05)。
3 討論
3.1 不同養殖密度下松江鱸的生長特征
硬骨魚的體長與體質量一般存在冪函數關系:m=aLb,式中a為條件因子,反映種群所處環境的優劣,b為異速生長因子,反映生長發育的不均勻性,如b<3為負異速生長,即體長增長快于體質量的增加,如b=3為等速生長,即體長和體質量等速增長,如b>3則為正異速生長,即體質量的增加快于體長的增長[6-8]。本試驗中,松江鱸體長與體質量關系可用公式m=aLb表達,式中a值隨養殖密度的增加而逐漸減小,A組(40尾/m2)的a值(0.008 9)最大,表明在此放養密度下松江鱸所處的生長環境更適合其生長。3個組的b值在3.123 6~3.209 4,且隨著養殖密度的增加逐漸增大,這表明養殖密度的提高加大了松江鱸生長的不均勻性,而這種不均勻性是由于體質量和體長的不均勻增長造成的。這與徐獻明等[9]對水泥池養殖松江鱸生長特性的研究有所不同(b=2.858 8)。養殖環境、營養和密度是造成差異的主要因素,這說明養殖環境、營養和密度可能影響松江鱸的生長模式。在今后的試驗中可嘗試通過調控環境、營養和密度等措施來達到體長與體質量增長的均勻性。此推論有待進一步試驗驗證。
擬合魚類生長的模型主要有Von Bertallanffy生長方程、冪指數生長方程、指數函數、直線方程、三次函數等。魚類生長方程能夠直觀地描述體長及體質量的變化情況。本試驗中,試驗魚的生長用三次函數可獲得較好的擬合效果。C組(120尾/m2)松江鱸的體長與體質量生長在試驗前期均高于A組(40尾/m2)和B組(80尾/m2)(見圖2)。隨著試驗的進行,松江鱸對空間和飼料的競爭加劇,各組松江鱸的生長發生變化:A組和B組松江鱸的體長、體質量分別在123.2 d和142.3 d、124.5 d和132.6 d后超過C組,表明本試驗條件下較低的養殖密度更有利于松江鱸的生長。
肥滿度是一種表征動物的生理或營養狀況的形態生理指數,是衡量魚體豐滿程度、營養狀況和環境條件的重要指標[10-11]。本試驗中,松江鱸的肥滿度隨著養殖試驗的進行呈現在波動中增長的趨勢,其數值為0.99~1.45,并在12月份達到最大值。各試驗組間肥滿度沒有顯著性差異(P>0.05)(見表1)。松江鱸屬于一年生魚類,其在養殖中后期所攝取的能量大多用于性腺發育,這導致其體質量的快速增長,因而在12月份達到最大值。另外,本試驗是在一定空間內的人工控制下的養殖狀態,魚體的肥滿度更大程度上是受飼養管理和營養調控質量的影響,肥滿度的變化僅反映了大致的范圍和趨勢。
3.2 養殖密度對松江鱸生長性能的影響
養殖密度是魚類生長過程中主要影響因子之一[12-13]。不同魚類的養殖密度存在一定的閾值,當養殖密度低于閾值,魚類成活率不受影響,而當養殖密度超過閾值,魚類成活率將隨著養殖密度升高呈下降趨勢[14]。本試驗中,C組(120尾/m2)松江鱸的成活率顯著低于A組(40尾/m2)和B組(80尾/m2)(P<0.05)(見表1),這表明松江鱸養殖密度的閾值約為120尾/m2,但具體閾值有待結合松江鱸的生長過程進行驗證。
研究表明,養殖密度是影響魚類生長、攝食及生理功能的重要因素[15]。一般認為,在養殖密度較低時,魚類生長不受密度影響;當達到一定密度后,魚類的生長速度將隨著密度增大而下降[16]。本試驗中,A組魚的終末體質量、終末體長、體質量日增長量、存活率均顯著高于其他兩組,其體長日增長量顯著高于C組(P<0.05)(見表1),且大部分指標呈現隨著養殖密度的增加而降低的趨勢。試驗期間,各組都保持了良好的水質和較高的溶解氧水平,基本可以排除水質條件對松江鱸生長的影響。這表明80尾/m2和120尾/m2的養殖密度對松江鱸的生長、攝食和生理功能產生了負面影響,導致其生長性能變差。因此,在本試驗條件下,松江鱸適宜的養殖密度是40尾/m2。
需要指出的是,在本試驗開始的前90 d,C組的生長速度略高于A組和B組(P>0.05)(見圖2~圖3),表明在此階段,120尾/m2不是松江鱸RAS養殖密度的閾值,該階段的養殖密度閾值仍有待進一步研究,這為松江鱸RAS養殖前期可采用高密度模式提供了數據和理論支撐。主要原因如下:(1)試驗在RAS中進行,水質狀況良好,投飼量充足,且投喂的配合飼料營養均衡;(2)經轉食馴化后的松江鱸在攝食配合飼料時存在集群效應[16],在養殖環境條件良好、養殖密度低于閾值時,較高的養殖密度可幫助其更有效地尋找食物,從而降低維持生存所消耗的能量,將更多的能量用于生長;而在較低的養殖密度條件下,松江鱸個體之間相互作用減少,會在一定程度上影響其攝食與生長。
3.3 水溫對松江鱸生長性能的影響
在養殖水產動物的過程中,水體溫度是影響水產動物生長的關鍵環境因素之一。魚類在一定溫度范圍內其生長速率隨溫度的升高而加快,但當超出其適宜生長溫度范圍時,其生長速率明顯減緩甚至死亡。
本試驗在90~150 d階段的平均水溫在23.5~24.5 ℃,在此期間各組試驗魚的體長特定生長率和體質量特定生長率均較低,并在該階段平均水溫最高(26.6 ℃)時達到最小值(見圖3)。這表明高溫對松江鱸生長的影響比較顯著,松江鱸在該養殖階段的適宜水溫以不超過25 ℃為宜。根本原因可能是水溫影響著松江鱸的攝食強度及代謝活動。研究表明,松江鱸苗種對高溫敏感,當水溫超過28 ℃時全部死亡[17];當養殖水溫超過26 ℃時,其攝食能力明顯下降,水溫超過28 ℃時,魚的活力下降[18];持續熱脅迫(水溫28 ℃以上)可導致其免疫功能下降,機體出現氧化損傷[19]。這與本試驗結果基本一致。另有研究表明,松江鱸耐低溫的能力很強,在較低溫度下仍能較快生長[20-22]。在高溫季節適當降低水溫能促進松江鱸生長[20],顯著增加其生長速度、降低死亡率[18],因此,在實際養殖生產中,建議加強養殖期間對水溫的控制和管理,從而提高松江鱸的生長性能。
3.4 養殖密度對生長離散的影響
養殖密度會影響魚類群體的生長空間和食物資源獲取率,造成養殖環境中生長資源分配不均,導致生長差異。本試驗中,不同養殖密度下松江鱸的體長、體質量變異系數差異性顯著,但B組體長與體質量變異系數均小于其余兩組(見表2)。從B組(80尾/m2)和C組(120尾/m2)在試驗90 d后的養殖情況來看,魚的生長離散程度隨養殖密度的增加而加劇。這可能是因為在高密度養殖水域,魚類對生存空間和食物資源的競爭加劇,競爭能力較弱的個體不能獲得充足的食物,其生長率進一步下降,而占優勢的魚幾乎不受影響或受影響較小,因此導致整個養殖群體的平均生長率下降,生長離散加劇。試驗結果顯示,A組(40尾/m2)魚的體長、體質量變異系數最大。原因可能是該組養殖密度最低,生存空間和食物資源壓力相對較小,魚的終末體長和終末體質量均顯著大于其余兩組,因而A組試驗魚的體長、體質量變異系數大于B組和C組。此外,松江鱸攝食飼料時存在的集群效應導致A組個體之間相互作用減少,影響了其攝食和生理機能,在一定程度上導致個體間生長差異增大。這與對2齡刀鱭[23]和大口黑鱸[24]生長離散的研究結果相似。
在實際生產中,養殖群體的體長、體質量變異系數越小,成魚的出池規格差異就越小,養殖效益就越高[25],并且高密度養殖所追求的高產量和經濟效益并非以高的生長效率為前提,而是以較大的種群個體基數為基礎[26]。本試驗中,B組(80尾/m2)的生長離散最小,且產量高出A組(40尾/m2)45.9%,因此,在RAS中并且投喂配合飼料的條件下,松江鱸養殖密度在80尾/m2左右較為合理。
4 結論
試驗結果表明,本試驗條件下,松江鱸體長與體質量均呈冪函數(m=aLb)相關,各試驗組的異速生長因子b值均大于3,且隨著養殖密度增加而逐漸增大,表現為正異速生長;體長、體質量與養殖天數均呈三次函數關系。
本試驗條件下,松江鱸的生長性能隨養殖密度的增加而降低,A組(40尾/m2)的終末體質量、終末體長、體質量日增長量和存活率均顯著高于B組(80尾/m2)和C組(120尾/m2),其體長日增長量顯著高于C組(P<0.05)。B組(80尾/m2)的體長、體質量變異系數最小,且產量顯著高于A組(40尾/m2)。在實際養殖生產中,群體的變異系數越小,成魚規格差異就越小,且高密度養殖所追求的高經濟效益是以較大的種群個體基數為基礎的。因此,建議配合飼料條件下RAS養殖松江鱸合理的養殖密度為40~80尾/m2。
本試驗條件下,建議養殖早期可采用高密度養殖模式(≥120尾/m2),具體閾值有待進一步研究。隨著魚體增長,應在約第120天前進行密度分稀及規格分篩等操作,將養殖密度降低至40~80尾/m2,以獲得較高產量和效益。建議實際生產中加強養殖期間對水溫的控制和管理,以提高松江鱸的生長性能。
參考文獻
[1]國家環境保護局,中華人民共和國瀕危物種科學委員會.中國瀕危動物紅皮書——魚類[M].北京:科學出版社,1998:240-243.
[2]張偉,徐建榮,徐獻民,等.松江鱸魚工廠化養殖試驗研究[J].漁業現代化,2017,44(1):6-9.
[3]王芳,劉小召,馮騫,等.工廠化養殖條件下大鯢適宜養殖密度的研究[J].淡水漁業,2013,43(6):73-77.
[4]張曉雁,李羅新,危起偉,等.養殖密度對中華鱘行為、免疫力和養殖環境水質的影響[J].長江流域資源與環境,2011,20(11):1348-1354.
[5]姚清華,顏孫安,郭清雄,等.養殖密度對瓦氏黃顙魚幼魚生長品質和生化指標的影響[J].福建農業學報,2018,33(7):670-675.
[6]黃真理,常劍波.魚類體長與體重關系中的分形特征[J].水生生物學報,1999,23(4):330-336.
[7]林學群.粵東近海雄性條尾鯡鯉體長與體重關系研究[J].汕頭大學學報(自然科學版),1999,14(2):64-71.
[8]FROESE R.Cube law,condition factor and weight-length relationships:history,meta-analysis and recommendations[J].Journal of Applied Ichthyology,2006,22(4):241-253.
[9]徐獻明,張志勇,韓曉磊,等.人工養殖松江鱸生長特征研究[J].水產養殖,2013,34(7):46-49.
[10]MURRAY D L.Differential body condition and vulnerability to predation in snowshoe hares[J].Journal of Animal Ecology,2002,71(4):614-625.
[11]BADIANI A,STIPA S,NANNI N,et al.Physical indices,processing yields,compositional parameters and fatty acid profile of three species of cultured sturgeon (Genus Acipenser)[J].Journal of the Science of Food and Agriculture,1997,74(2):257-264.
[12]LARSEN B K,SKOV P V,MCKENZIE D J,et al.The effects of stocking density and low level sustained exercise on the energetic efficiency of rainbow trout(Oncorhynchus mykiss) reared at 19 ℃[J].Aquaculture,2012(324/325):226-233.
[13]LI D P,LIU Z D,XIE C X.Effect of stocking density on growth and serum concentrations of thyroid hormones and cortisol in Amur sturgeon,Acipenser schrenckii[J].Fish Physiology and Biochemistry,2012,38(2):511-520.
[14]廖銳,區又君,勾效偉.養殖密度對魚類福利影響的研究進展Ⅰ.死亡率、生長、攝食以及應激反應[J].南方水產,2006,2(6):76-80.
[15]王威,姜雪照,張漢邦,等.放養密度對池塘循環水養殖翹嘴鱖生長的影響[J].淡水漁業,2020,50(3):94-98.
[16]畢詳,張飛明,張友良,等.松江鱸苗種轉食馴化技術研究[J].水產科技情報,2018,45(3):137-140.
[17]劉玉峰.松江鱸苗種繁育和養成技術的研究[D].保定:河北農業大學,2011.
[18]張飛明,張友良,謝志強.松江鱸魚養殖技術之五松江鱸魚成魚養殖技術[J].科學養魚,2011(12):14-16.
[19]劉佳奇,劉玉峰,何忠偉,等.熱脅迫對松江鱸免疫功能的影響[J].河北漁業,2022(3):1-3.
[20]韋正道,王昌燮,杜懋琴,等.控制松江鱸魚(Trachidermus fasciatus)生長的環境因子的研究[J].復旦學報(自然科學版),1997,36(5):581-585.
[21]李永民,沈宗岳.松江鱸魚網箱越冬的試驗[J].水產科技情報,1985,12(2):8-9.
[22]宋向軍,王忠全,周全利,等.松江鱸魚人工配合飼料投喂實驗研究[J].海洋湖沼通報,2014(1):74-80.
[23]劉永士,王建軍,朱建明,等.養殖密度對2齡刀鱭生長的影響[J].大連海洋大學學報,2021,36(1):66-73.
[24]莫介化,李春枝,王曉斌,等.不同養殖密度大口黑鱸在室內工廠化循環水養殖系統中的生長特性[J].淡水漁業,2022,52(3):98-104.
[25]姜虎成,夏愛軍,張明生,等.人工養殖條件下麥穗魚的生長特性[J].中國農學通報,2017,33(35):137-140.
[26]莊平,李大鵬,王明學,等.養殖密度對史氏鱘稚魚生長的影響[J].應用生態學報,2002,13(6):735-738.
Effects of stocking density on growth of Trachidermus fasciatus in the recirculating aquaculture system
GAO Guiming, JI Qiang, HUANG Di
(Shanghai Songjiang Fisheries Technical Extension Station,Shanghai 201600,China)
Abstract: To investigate the influence of stocking density on the growth of Trachidermus fasciatus in the recirculating aquaculture system(RAS) under the condition of formula feed,T. fasciatus with initial body length of (2.97±0.12)cm and initial body mass of (0.26±0.03)g were cultured in RAS at stocking densities of 40 ind/m2(group A),80 ind/m2(group B) and 120 ind/m2(group C).The test period was 240 days.The results showed that the highest body weight,body length,daily gain and survival rate was observed in group A,and the daily growth of body length in group A was significantly higher than group C(P<0.05).There was no significant difference in coefficient of fatness among groups(P>0.05).Output per unit area of 3 groups:group A(1.72 kg/m2) Key words: Trachidermus fasciatus; recirculating aquaculture system; stocking density; formula feed; growth
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
