β-胡蘿卜素強化劑對鹵蟲體內類胡蘿卜素組成的影響
2023-09-06 16:13:10陳慧聰薛毓才王偉隆黃旭雄
海洋漁業 2023年3期
陳慧聰,王 茜,薛毓才,王偉隆,2,3,黃旭雄,2,3
(1.上海海洋大學,水產科學國家級實驗教學示范中心,上海 201306;2.上海海洋大學,上海市水產養殖工程技術研究中心,上海 201306;3.上海市水產動物良種創制與綠色養殖協同創新中心,上海 201306)
類胡蘿卜素(carotenoids)是廣泛存在于動植物及微生物中的一類基于C40異戊二烯結構的脂溶性色素[1]。在水產養殖中,類胡蘿卜素由微生物合成,再通過浮游動物、甲殼動物、魚類這條食物鏈逐級積累[2]。集約化水產養殖造成群落結構發生變化、食物鏈單一、有效色素源少,是導致養殖對象體內類胡蘿卜素缺乏的重要原因[3]。在飼料中添加類胡蘿卜素可以有效改善水產動物色乏、味寡、香弱、抗病力差及繁殖能力低下等問題,因而受到廣泛關注[4]。目前在水產動物飼料中添加的類胡蘿卜素主要為胡蘿卜素類(結構中不含氧的烴類類胡蘿卜素,以β-胡蘿卜素為主)和葉黃素類(以隱黃質、玉米黃質、海膽酮、角黃素和蝦青素為主)兩大類。
β-胡蘿卜素(β-carotene)作為維生素A的前體,具有很高的生物活性,可預防因維生素A缺乏導致的疾病[5]。水產動物可以直接利用飼料中添加的類胡蘿卜素或將其轉化為其他類胡蘿卜素沉積在不同組織中[6]。作為抗氧化劑,β-胡蘿卜素可以抑制脂質過氧化,保護生殖細胞免受氧化反應的損傷,提高水產動物的繁殖力及孵化率[7]。另外,β-胡蘿卜素的強抗氧化作用可以有效淬滅活性氧和自由基,保護機體白細胞免受損傷,從而提高水產動物的非特異性免疫機能[8]。
鹵蟲(Artemia)幼體和成蟲是海水養殖苗種培育中重要的生物餌料[9]。利用鹵蟲具有非選擇性濾食的特性,將其作為活載體,使鹵蟲腸道包裹并攜帶特定強化劑,可以有效地滿足水產動物對必需脂肪酸、氨基酸、脂溶性維生素等的需求[10]。本研究利用β-胡蘿卜素作為強化劑,測定分析不同強化劑量對鹵蟲體內類胡蘿卜素組成和含量變化的影響,評估鹵蟲作為類胡蘿卜素生物載體的可行性,同時探索鹵蟲體內β-胡蘿卜素的代謝規律,以期提高鹵蟲在水產養殖中的生物餌料價值。
1 材料與方法
1.1 實驗材料
實驗用渤海鹵蟲干卵購自山東濱州前華水產有限公司,鹵蟲卵在25 L孵化桶內進行孵化,孵化密度2 g·L-1,孵化在鹽度25的過濾人工海水中進行,水溫27~28℃,pH =8,連續光照充氣,孵化24 h后分離一部分無節幼體用于后續β-胡蘿卜素強化實驗,另一部分無節幼體在實驗室環境下繼續培養至15日齡成蟲后進行β-胡蘿卜素強化實驗。類胡蘿卜素檢測用的蝦青素、玉米黃質、角黃素、海膽酮和β-胡蘿卜素標準品均購于美國Sigma公司。
1.2 實驗設計
1.2.1 β-胡蘿卜素強化實驗
配制5個濃度梯度的人工合成β-胡蘿卜素強化劑。分別稱取15、45、75、105、135 mg的人工合成β-胡蘿卜素(純度≥98%,Adamas)于1 L燒杯中,加入20 mL吐溫80,充分攪拌,逐漸加入980 mL鹽度25的海水,繼續攪拌直至均勻乳化。并設1個不添加β-胡蘿卜素的對照組,對照組只含有20 mL吐溫80的海水(鹽度25)。
將孵化好的鹵蟲無節幼體/成蟲分別轉移到裝有14 L海水的20 L塑料桶中,鹵蟲無節幼體密度為30尾·mL-1,成蟲密度為1尾·mL-1。每個實驗桶分別添加1 L上述不同濃度(15、45、75、105、135 mg·L-1)的β-胡蘿卜素強化劑以及對照組溶液,使得桶內β-胡蘿卜素濃度分別為0、1、3、5、7、9 mg·L-1,共6個處理組,每組設3個平行。在光照2 000 lx、水溫25℃、pH =8及持續充氣的條件下,強化24 h后收集鹵蟲。
1.2.2 營養強化后饑餓實驗
根據1.2.1方法用不同濃度β-胡蘿卜素強化劑強化鹵蟲無節幼體(成蟲)24 h后,用200目篩絹收集鹵蟲,并用鹽度25的海水沖洗掉表面的β-胡蘿卜素強化劑,轉移至裝有15 L海水的塑料桶中,在與1.2.1相同培養條件下,投喂適量無色素酵母12 h后收集鹵蟲。
1.3 類胡蘿卜素含量測定
1.3.1 鹵蟲樣品收集方法
無節幼體:從強化實驗水體中取1 mL水樣于浮游動物計數框中,用甲醛固定,在體式鏡(LEICA S9E,Germany)下計數,重復6次。然后將水體中的鹵蟲無節幼體濾出,收集于10 mL離心管中,計算總數,冷凍干燥后用于類胡蘿卜素檢測。
成蟲:隨機取50只鹵蟲成蟲收集于10mL離心管中,冷凍干燥后用于類胡蘿卜素檢測。
1.3.2 鹵蟲樣品前處理
將丙酮-甲醇(v∶v,2∶1)萃取劑3 mL加入1.3.1收集的鹵蟲樣本中,充分渦旋振蕩,24 kHz下冰浴超聲處理30 min,再35℃浸提1 h后,4 000 r·min-1離心10 min,取上清液,對殘余料渣重復提取3次。合并上清液后用氮氣吹干,加入3 mL甲醇-乙腈(v∶v,3∶7)溶劑使其完全溶解,經0.22μm親水濾膜過濾后加入進樣瓶中備檢。
1.3.3 類胡蘿卜素檢測標準品標定
5種類胡蘿卜素混合標準溶液:分別稱取5種類胡蘿卜素標準品1mg混合,用正己烷定容為50μg·mL-1的類胡蘿卜素標準儲備液。分別移取上述標準儲備液0.04、0.20、0.40、1.00、4.00 mL于5個10 mL容量瓶中混合均勻,用氮氣吹干后,加入含0.1% BHT的甲醇-乙腈(v∶v,3∶7)溶劑,將其定容為0.2、1.0、2.0、5.0、20.0 μg·mL-1的5個梯度類胡蘿卜素混合標準溶液,經0.22μm親水濾膜過濾后加入進樣瓶中備檢。每組中類胡蘿卜素含量以μg·萬只-1計。
采用超高效液相色譜儀(ultra-high performance liquid chromatography,UPLC,Waters,US)配用ACQUITY H-Class BEH C18色譜柱(1.7μm,2.1 mm ×150 mm)用于類胡蘿卜素的分離和測定。流動相A:100% 超純水;流動相B:甲醇-乙腈(v∶v,3∶7)溶液;流動相C:100% 叔丁基甲基醚溶液。梯度洗脫條件:10% A +90% B 16 min,20% C+80% B 14 min,流速0.3 mL·min-1,進樣量10μL,柱溫25℃,檢測波長475 nm(圖1)。
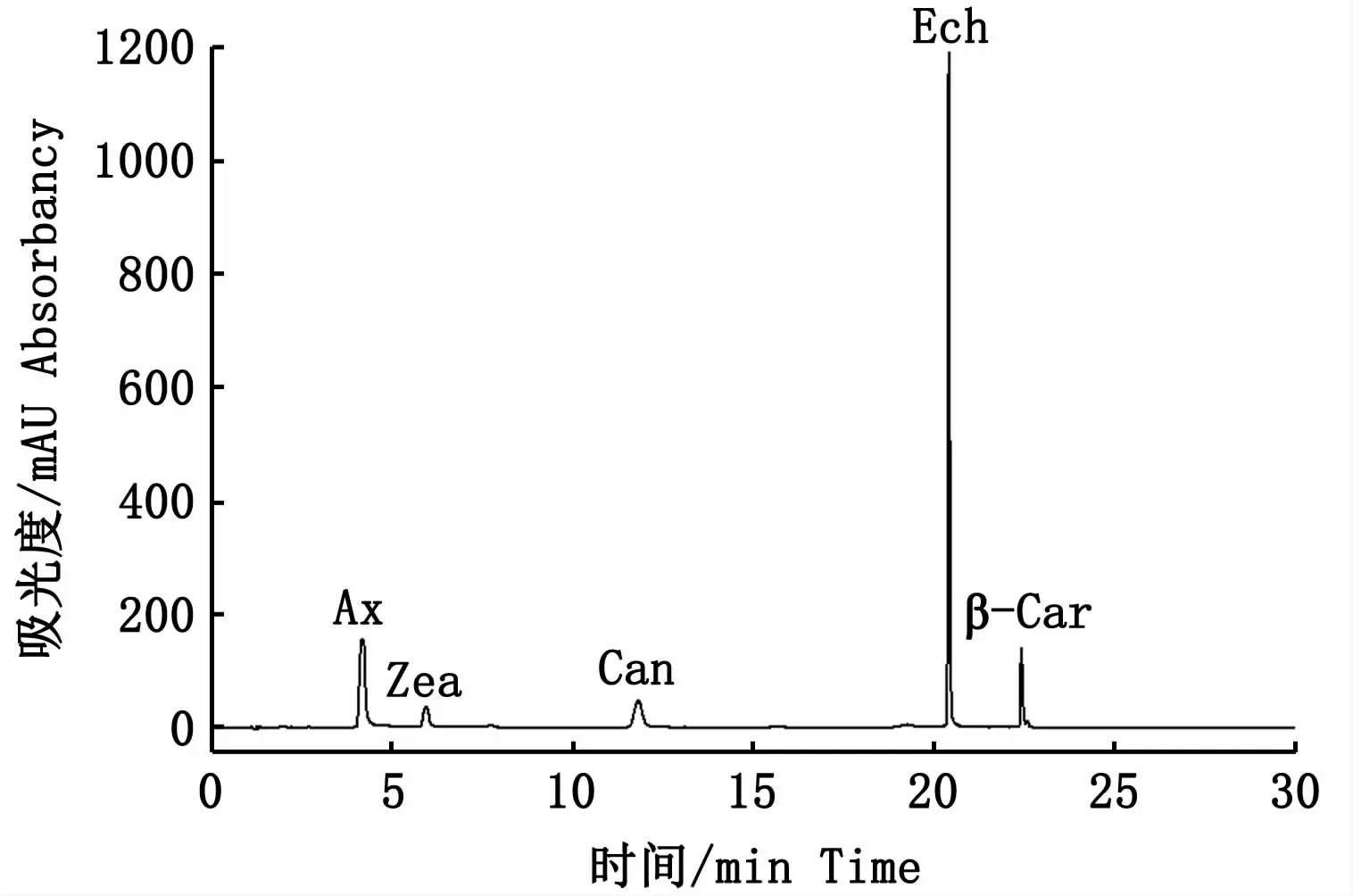
圖1 梯度洗脫條件下5種類胡蘿卜素混標峰圖Fig.1 Peak curves of five carotenoids w ith gradient elution conditions
1.4 數據統計方法
實驗結果用平均值 ±標準差(mean±SD)表示,數據采用SPSS 22.0軟件進行單因素方差分析(one-way ANOVA),用Duncan’s法進行多重比較檢驗,用t檢驗分析饑餓前后鹵蟲體內類胡蘿卜素含量差異。P<0.05表示差異顯著。
2 結果與分析
2.1 鹵蟲無節幼體強化24 h后體內類胡蘿卜素含量的變化
在強化前和對照組的鹵蟲無節幼體中僅檢測到角黃素和海膽酮兩種類胡蘿卜素,未檢測到β-胡蘿卜素。強化24 h后,強化組無節幼體的β-胡蘿卜素含量隨著強化劑濃度的增加顯著上升(P<0.05)(圖2-a)。當β-胡蘿卜素強化劑量大于3 mg·L-1時,無節幼體體內海膽酮含量顯著高于強化前和對照組(P<0.05),其中5 mg·L-1和7 mg·L-1β-胡蘿卜素強化組鹵蟲體內的海膽酮含量最高,且兩組間無顯著性差異(P>0.05)(圖2-b)。經過24 h強化后,對照組無節幼體體內角黃素含量顯著低于3 mg·L-1和5 mg·L-1強化組(P<0.05),其中5 mg·L-1強化組無節幼體體內角黃素含量最高。
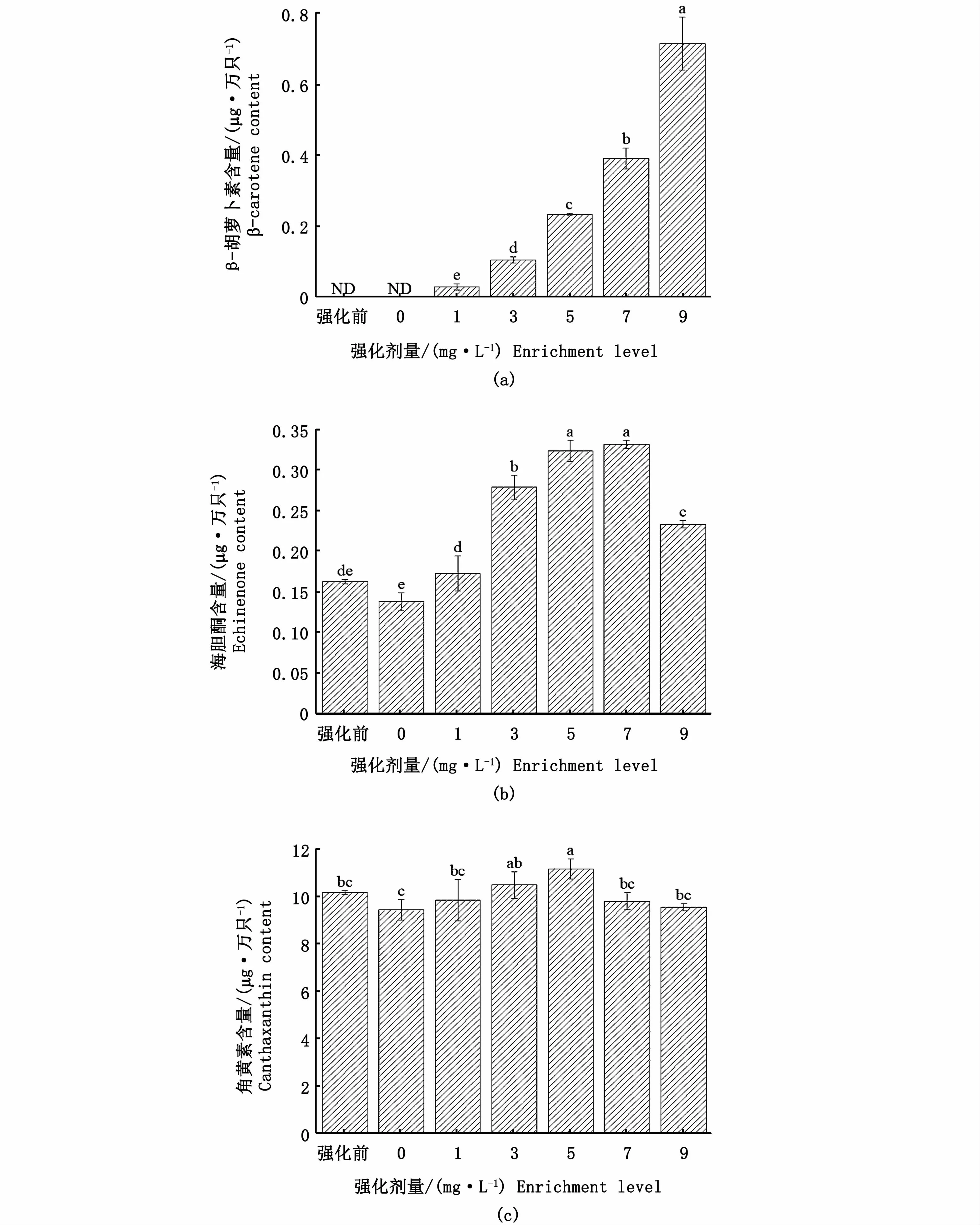
圖2 不同劑量β-胡蘿卜素強化鹵蟲無節幼體24 h后體內類胡蘿卜素含量變化Fig.2 Carotenoid content changes in Artem ia nauplius after different levels ofβ-carotene enrichment for 24 h
2.2 鹵蟲無節幼體強化24 h后再饑餓12 h體內類胡蘿卜素含量變化
由圖3可見,饑餓12 h后,各強化組無節幼體體內β-胡蘿卜素含量均顯著下降(P<0.05),低劑量強化組(1、3、5 mg·L-1)組間差異不顯著(P>0.05)(圖3-a)。饑餓后各組無節幼體體內海膽酮含量也呈現下降趨勢,且對照組和3、5、7、9 mg·L-1強化組海膽酮含量較12 h前顯著降低(P<0.05);饑餓12 h后,5 mg·L-1和7 mg·L-1強化組海膽酮含量顯著高于其他組別,對照組顯著低于其他組別(P<0.05)(圖3-b)。饑餓后各組無節幼體角黃素含量較饑餓前有所下降,但均無顯著性差異(P>0.05);饑餓后5 mg·L-1組無節幼體角黃素含量依然最高,顯著高于其他組別(P<0.05),其他5組間無顯著性差異(P>0.05)(圖3-c)。
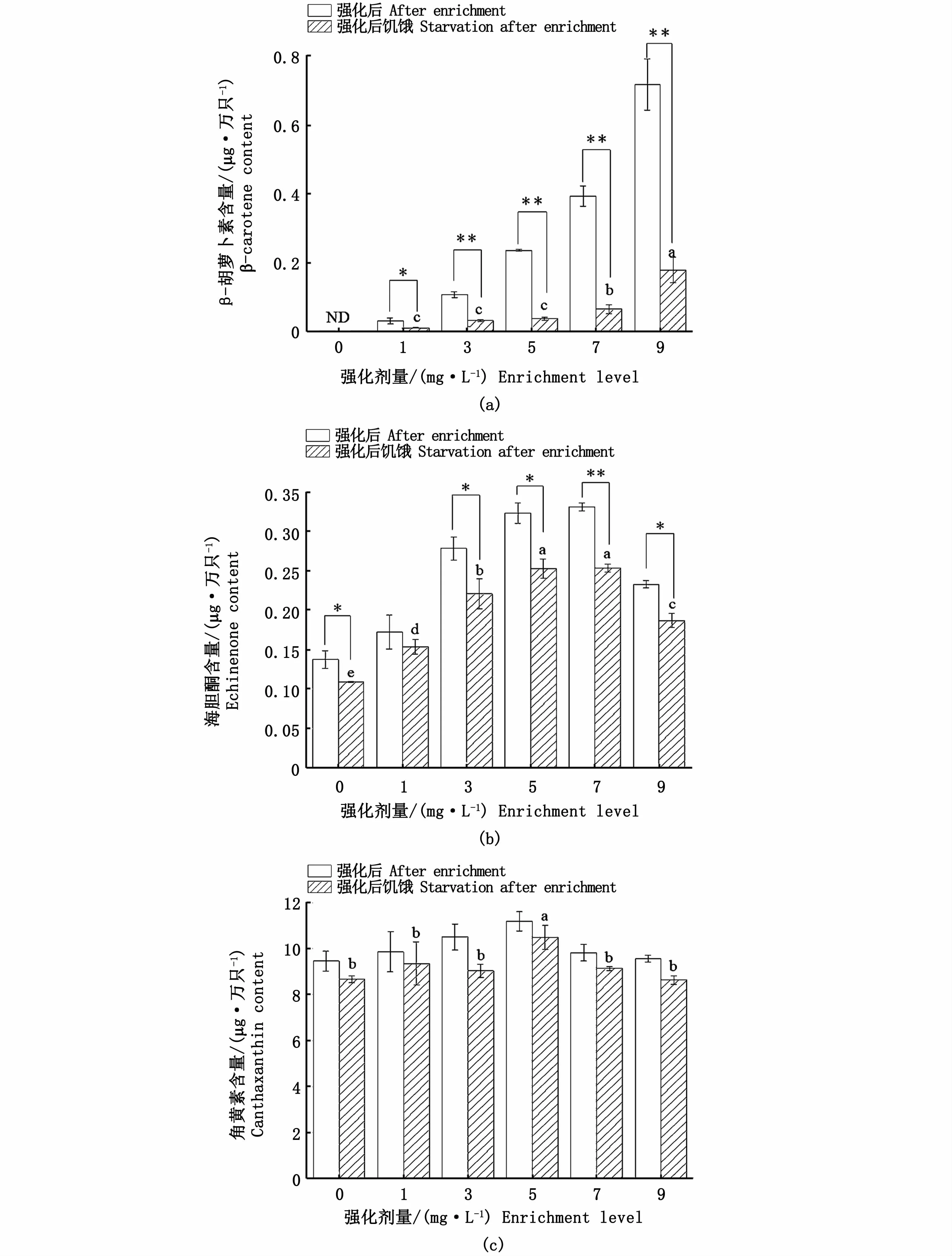
圖3 不同劑量β-胡蘿卜素強化鹵蟲無節幼體24 h后再饑餓12 h體內類胡蘿卜素含量變化Fig.3 Carotenoid content changes in Artemia nauplius after different levels of β-carotene enrichment for 24 h and then starvation for 12 h
2.3 鹵蟲成蟲強化24 h后體內類胡蘿卜素含量變化
在強化前及對照組的鹵蟲成蟲體內僅檢測到角黃素和海膽酮兩種類胡蘿卜素,未檢測到β-胡蘿卜素。β-胡蘿卜素強化后,各組鹵蟲成蟲體內β-胡蘿卜素含量隨著強化劑量增大先呈上升趨勢;當劑量大于7 mg·L-1時,鹵蟲體內β-胡蘿卜素含量則下降(圖4-a)。1、3、5 mg·L-1β-胡蘿卜素強化組在強化24 h后,鹵蟲成蟲體內海膽酮含量降低;而高劑量強化組(7 mg·L-1和9 mg·L-1)鹵蟲成蟲體內海膽酮含量顯著高于其他各組(P<0.05)(圖4-b)。β-胡蘿卜素強化后,強化組鹵蟲成蟲體內角黃素含量顯著高于強化前和對照組(P<0.05);其中,9 mg·L-1強化組又顯著高于其他組別(P<0.05),而低劑量強化組(1、3、5 mg·L-1)之間沒有顯著性差異(P>0.05)(圖4-c)。
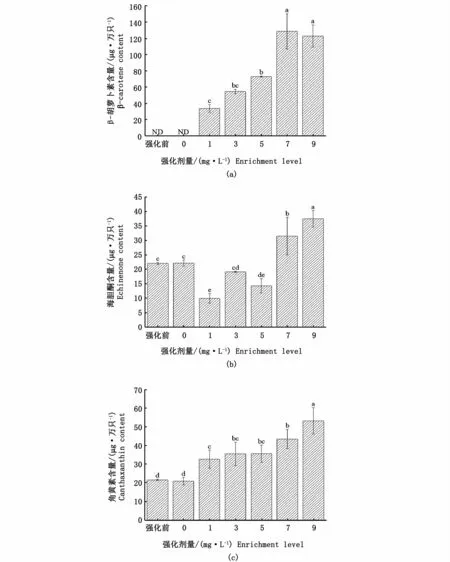
圖4 不同劑量β-胡蘿卜素強化鹵蟲成蟲24 h后體內類胡蘿卜素含量變化Fig.4 Carotenoid content changes in Artem ia adu lts after different levels ofβ-carotene enrichm ent for 24 h
2.4 鹵蟲成蟲強化24 h后再饑餓12 h體內類胡蘿卜素含量變化
由圖5可知,各強化組鹵蟲成蟲饑餓12 h后,體內β-胡蘿卜素含量較饑餓前均極顯著降低(P<0.01);其中,9 mg·L-1強化組鹵蟲饑餓后體內β-胡蘿卜素含量顯著高于其他各組(P<0.05)(圖5-a)。低劑量強化組(1、3、5 mg·L-1)在饑餓12 h后,體內海膽酮含量顯著低于對照組和高劑量強化組(7、9 mg·L-1)(P<0.05)(圖5-b)。對照組角黃素含量在饑餓前后沒有發生顯著性變化(P>0.05),強化組角黃素含量在饑餓后總體上呈上升變化的趨勢,但各組饑餓前后差異均不顯著(P>0.05);其中,9 mg·L-1強化組鹵蟲體內角黃素含量最高,且顯著高于除7 mg·L-1強化組外的其他各組(P<0.05)(圖5-c)。
3 討論
以往研究表明,水產動物自身無法合成類胡蘿卜素,僅可利用飼料中的類胡蘿卜素直接儲存或轉化后沉積于體內[11]。野生水產動物由于其生長環境中富含能夠合成類胡蘿卜素的藻類和細菌,在生長過程中不會出現因類胡蘿卜素營養缺乏導致的體色及免疫活性方面的問題[12]。但在高密度集約化養殖過程中,人工配合飼料組成中易發生有效色素源少且活性不穩定等情況,造成類胡蘿卜素這一必須從外源食物中攝取的功能性代謝產物的嚴重缺乏,而類胡蘿卜素長期不足將直接或間接導致養殖對象體色異常、風味差及抗病能力低于野生品種。
鹵蟲屬于較低等的甲殼動物,在自然環境中主要以細菌和微藻為食,可作為第二營養級向高營養級傳遞能量,滿足水產動物各時期的營養需求[13]。本文研究結果表明,在自然條件下生長的鹵蟲無論是在無節幼體還是成蟲階段,在色譜測定條件無變化和沒有外源性胡蘿卜素干擾的情況下,在鹵蟲體內只檢測到角黃素和海膽酮兩種類胡蘿卜素。這一發現與HSU等[14]在加利福尼亞鹵蟲卵以及初孵無節幼體體內得到的檢測結果一致。GILCHRIST和GREEN[15]的實驗結果表明,鹵蟲體內檢測到的β-胡蘿卜素和蝦青素等類胡蘿卜素很可能來源于生長環境中富含能夠生產相應類胡蘿卜素的藻類(如杜氏鹽藻Dunaliellasalina、雨生紅球藻Haematococcuspluviali等)。為了進一步驗證以上結論,本實驗選用不同梯度的β-胡蘿卜素分別對鹵蟲無節幼體及成蟲進行營養強化。實驗結果表明,在β-胡蘿卜素強化24 h后,隨著β-胡蘿卜素強化劑濃度增加,鹵蟲體內β-胡蘿卜素含量總體上呈現顯著上升變化趨勢。這一結果可能是由于強化后鹵蟲腸道內β-胡蘿卜素沒有排空,充分發揮了鹵蟲作為生物載體對營養物質的包裹富集作用,也論證了鹵蟲成為類胡蘿卜素載體的可行性。
甲殼動物體內的類胡蘿卜素包括胡蘿卜素類(carotenes)和葉黃素類(xanthophylls)兩類。胡蘿卜素類以β-胡蘿卜素為主;葉黃素類則是胡蘿卜素的含氧衍生物,主要包括隱黃質、玉米黃質、海膽酮、角黃素和蝦青素等[13-14]。其中,隱黃質、玉米黃質、海膽酮、角黃素和蝦青素的化學結構與β-胡蘿卜素相似,主要由中央多聚鏈和位于兩側的芳香環組成,區別在于每個芳香環上的3,3’和4,4’端具有不同數量的羥基和酮基。眾所周知,物質結構決定其功能,這些類胡蘿卜素以蛋白共軛或者脂肪酸酯合形式在甲殼動物組織中的選擇性積累,不僅表明結構差異性對水產動物的生長發育和健康產生的不同生理作用,也增加了類胡蘿卜素與其他生物分子之間相互作用的復雜性[15-16]。已有研究表明,甲殼動物可以通過代謝轉化不同外源類胡蘿卜素以滿足機體正常發育過程中對類胡蘿卜素的營養需求[4]。傳統的“對蝦模型”代謝轉換方式,可以將β-胡蘿卜素、隱黃質、角黃素和玉米黃質轉化為蝦青素,而酮基化酶和羥基化酶的功能特性決定了這兩種酶在類胡蘿卜素代謝轉化過程中的關鍵作用[8,17]。而在代謝轉化過程中,兩種酶的交替作用也會產生多條不同的中間產物的補充途徑[4]。迄今為止,類胡蘿卜素代謝轉化途徑中所需關鍵酶基因及其作用機制的研究僅在少數自身能夠合成蝦青素的微生物中有比較系統與深入的開展,涉及甲殼動物的相關研究還較為匱乏[18-19],僅在測定橈足類高通量的轉錄組文庫后,通過生物信息學分析篩選到代謝轉化途徑中差異表達的β-胡蘿卜素羥基化酶基因[20],但對其作用機制的研究還未見報道。在觀察強化后鹵蟲體內角黃素和海膽酮含量變化的過程中發現,強化24 h后的鹵蟲成蟲體內角黃素含量較強化前總體上呈顯著上升趨勢,且在饑餓12 h后鹵蟲腸道β-胡蘿卜素排空的情況下,其體內角黃素含量不降反升,充分證實鹵蟲體內存在從β-胡蘿卜素到角黃素的轉化途徑。分析海膽酮(4-keto-β-carotene)和角黃素(4,4’-diketo-β-carotene)兩種物質結構,發現這兩種類胡蘿卜素都屬于酮基類胡蘿卜素。而且在鹵蟲卵、初孵無節幼體以及成蟲中均未檢測出玉米黃質(3,3’-dihydroxy-β-carotene)和蝦青素(3,3’-dihydroxy-4,4’-diketo-βcarotene)等羥基類胡蘿卜素中間代謝產物,可以推測鹵蟲體內類胡蘿卜素代謝轉化過程中,可能只有β-胡蘿卜素酮基化酶的參與,而缺少羥基化酶的作用。實驗結果還表明,鹵蟲成蟲的轉化能力較無節幼體更強,能夠在胡蘿卜素傳遞過程中滿足多級營養需求。
為提高營養強化效率,今后研究還可從以下幾個方面開展:選擇合適的外源類胡蘿卜素強化劑;加強鹵蟲類胡蘿卜素營養動力學研究;細化品系篩選和強化條件;發掘鹵蟲體內參與代謝轉化的功能酶基因等。
