近江牡蠣受精過程、胚胎及早期幼蟲發育特征
2023-09-06 16:13:10葛廣玉孫秀俊周麗青劉志鴻
海洋漁業 2023年3期
王 巖,吳 彪,葛廣玉,4,陳 夕,孫秀俊,周麗青,劉志鴻
(1.農業農村部海洋漁業可持續發展重點實驗室,中國水產科學研究院黃海水產研究所,山東青島 266071;2.青島海洋科學與技術試點國家實驗室,海洋漁業科學與食物產出過程功能實驗室,山東青島 266071;3.水產科學國家級實驗教學示范中心,上海海洋大學,上海 201306;4.國家海洋設施養殖工程技術研究中心,浙江海洋大學,浙江舟山 316022)
近江牡蠣(Crassostreaariakensis)隸屬于雙殼綱(Bivalvia),牡蠣科(Ostreidae),巨牡蠣屬,是廣溫、廣鹽性貝類,自然種群多棲息于河口區,在我國南北方均有分布。近江牡蠣是我國重要的牡蠣養殖品種之一,具有較高的營養價值,也是構成牡蠣礁的主要種類,在濾除水體污染物、維持生物多樣性等方面發揮重要的生態功能,素有“生態系統工程師”之美譽[1-6]。近年來,因海洋環境變化、過度捕撈、人工苗種補充有限等因素影響,我國沿海地區,尤其是北方海區近江牡蠣自然資源量不斷衰減[7-9]。開展人工苗種規模化繁育可為補充自然苗種和資源增殖提供支撐和保障,同時也是進行種質改良的必要前提。
受精及早期胚胎發育過程是水產經濟動物苗種繁育、遺傳育種領域的重要研究內容。充分了解養殖對象的受精和胚胎發育特點,可為人工苗種培育、細胞工程育種等工作提供重要參考。采用普通光學顯微鏡和熒光顯微觀察技術進行觀察具有簡便、清晰、直觀等優點,已在多種經濟貝類受精過程的核行為變化和胚胎早期發育觀察相關研究中應用。例如,有國外學者利用熒光顯微觀察技術對合浦珠母貝(Pinctada martensii)、侏儒蛤(Mulinialateralis)等經濟貝類在受精過程中的核相變化進行了詳細的觀察研究[10-11]。已報道的對貝類受精和早期卵裂過程進行觀察研究的品種還包括文蛤(Meretrix meretrix)、毛蚶(Scapharcasubcrenata)、斧文蛤(M.lamarkii)、縊蟶(Sinonovaulaconstricta)、青蛤(Cyclina sinensi)、管角螺(Hemifusus tuba)等[12-17],這些研究為貝類人工苗種繁育提供了重要參考。本研究以近江牡蠣北方群體作為親本,在溫度22~24℃、鹽度20~22的條件下進行人工授精,利用普通光學顯微鏡和熒光顯微觀察技術對其受精及早期胚胎發育過程進行了觀察研究,以期為近江牡蠣的苗種繁育、遺傳育種等研究提供基礎資料。
1 材料與方法
1.1 實驗材料
實驗用近江牡蠣親貝采自山東省丁字灣海域(36°37′N、120°51′E),平均殼長(21.73±4.01)cm。將親貝沖洗干凈后,在溫度22~24℃、鹽度20~22的充氣水體中暫養2 d,繁養期間每4 h 投喂小新月菱形藻(Nitzschia closteriumf.minutissima)1次,餌料的單次投喂量為每1 mL水體中投喂1×105個。
1.2 實驗設計
1.2.1 近江牡蠣性腺切片觀察
選用性腺飽滿的近江牡蠣親本進行解剖。解剖后,從性腺部位取少量樣品涂于載玻片通過顯微鏡判斷性別,并取一部分性腺組織置于Bouin氏液(Solarbio公司)中固定24 h后,用于后續切片制作,以進一步確定親貝的性腺發育狀況。切片參考芮菊生等[18]組織切片方法進行,分別取雌、雄近江牡蠣1 cm2性腺組織置于Bouin氏液中固定24 h,之后更換為70%酒精,酒精每日更換,共更換2~3次,樣品經梯度脫水、浸蠟包埋、切片、蘇木素-伊紅(H.E)染色后,使用LEICA DM4000光學顯微鏡觀察并拍照。
1.2.2 人工授精、早期胚胎材料固定及顯微觀察
確定雌、雄性別后,取性腺發育良好的個體,用干凈海水分別將精、卵沖洗收集至盛有砂濾水(鹽度20)的燒杯中浸泡20 min,以激活卵子和精子。利用300目篩絹過濾多余組織塊后進行人工授精,保持顯微鏡視野下1個卵子周圍有5~8個精子,精、卵混合后需不斷攪拌使其均勻分布,并在受精完成后轉入100 L的塑料桶中進行充氣孵化。
受精后,分別于20、40、60、80、100 min和2、3、6、9、12、18、24 h取樣。取樣時,過濾掉多余海水,將樣品轉入1.5 mL的離心管中(每個時間點取3管平行),加入4%多聚甲醛溶液(pH 7.4、0.1 mol·L-1PBS配制)固定,每隔2 h更換1次固定液,更換2次后,置于4℃冰箱內保存備用。
用于普通光學顯微鏡觀察的樣品直接滴在載玻片上,加蓋玻片后,于LEICA DM4000光學顯微鏡下觀察并拍照。用于熒光觀察的樣品在鏡檢前用0.1 mol·L-1PBS(pH 7.4)沖洗3次,再用Hoechst 33258熒光染料(江蘇碧云天生物技術研究所)在黑暗環境下染色15 min后,用PBS沖洗3次,然后滴于載玻片上,滴加抗熒光淬滅劑后,用蓋玻片封片,在LEICA DM4000熒光顯微鏡下觀察、拍照。
1.2.3 胚胎早期發育
胚胎發育至D形幼蟲時,停止曝氣1 h,用300目篩絹選取上浮幼蟲進行培養。幼蟲培育環境條件為:水溫22~24℃,鹽度20~22,pH 8.3~8.4,每天換水2次,每次換水量50%,并投喂球等鞭金藻(Isochrysisgalbana),每4 h投喂1次,單次投喂量為每1 mL水體中投喂5×103個。在此期間,跟蹤幼蟲的生長和發育情況,方法如下:用篩絹篩取幼蟲,用一次性滴管吸取樣品置于透明載玻片上,于LEICA DM4000光學顯微鏡下拍照,并通過顯微鏡自帶軟件Toupview 3.7測量幼蟲的殼長,用于幼蟲日平均生長率的計算與分析,幼蟲日平均生長率=(平均殼長增長量/發育天數)×100%。
2 結果與分析
2.1 近江牡蠣親本性腺外觀及切片觀察
本實驗獲取的近江牡蠣性腺飽滿,整個內臟團被乳白色性腺包圍著,內臟團周圍性腺厚度較厚,解剖刀刺破性腺表面,有白色、粘稠狀液體流出,表明性腺已發育成熟(圖1)。組織學觀察結果表明,性腺濾泡體積大,濾泡中的大多數生殖細胞處于成熟期。雄性個體濾泡內存在大量成熟精子,排列緊密,濾泡壁周圍有精原細胞出現,此時期的生殖帶清晰可見,成熟的精子位于生殖帶上(圖2-A)。雌性個體的濾泡相互擠壓,相互之間基本無空隙,濾泡呈多邊形,其中排滿大量成熟卵子(圖2-B)。結合外觀和組織切片觀察,繁育用牡蠣性腺已達到成熟期。
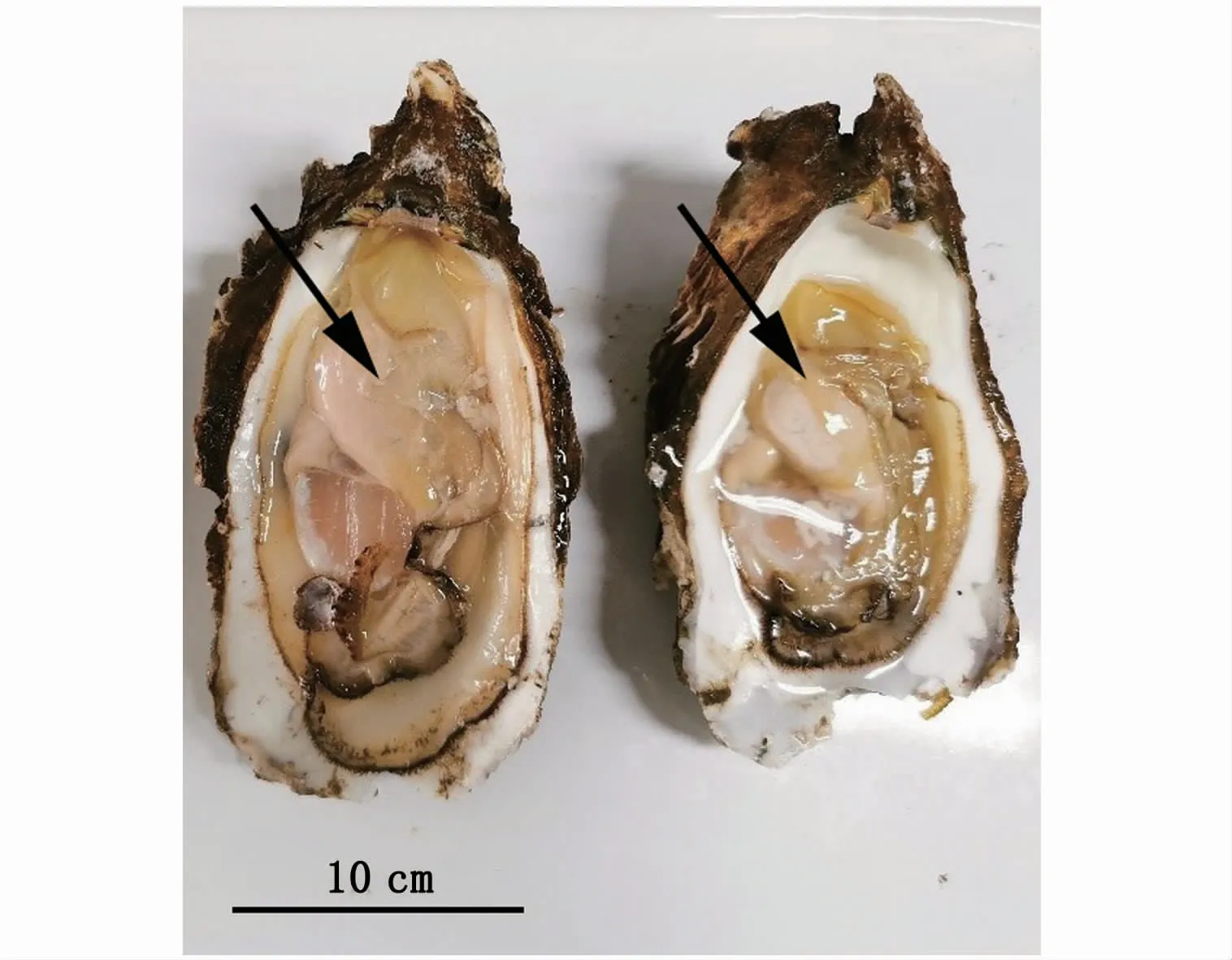
圖1 近江牡蠣親本性腺外觀(×20)Fig.1 Appearance of gonads in parents of C.ariakensis(×20)
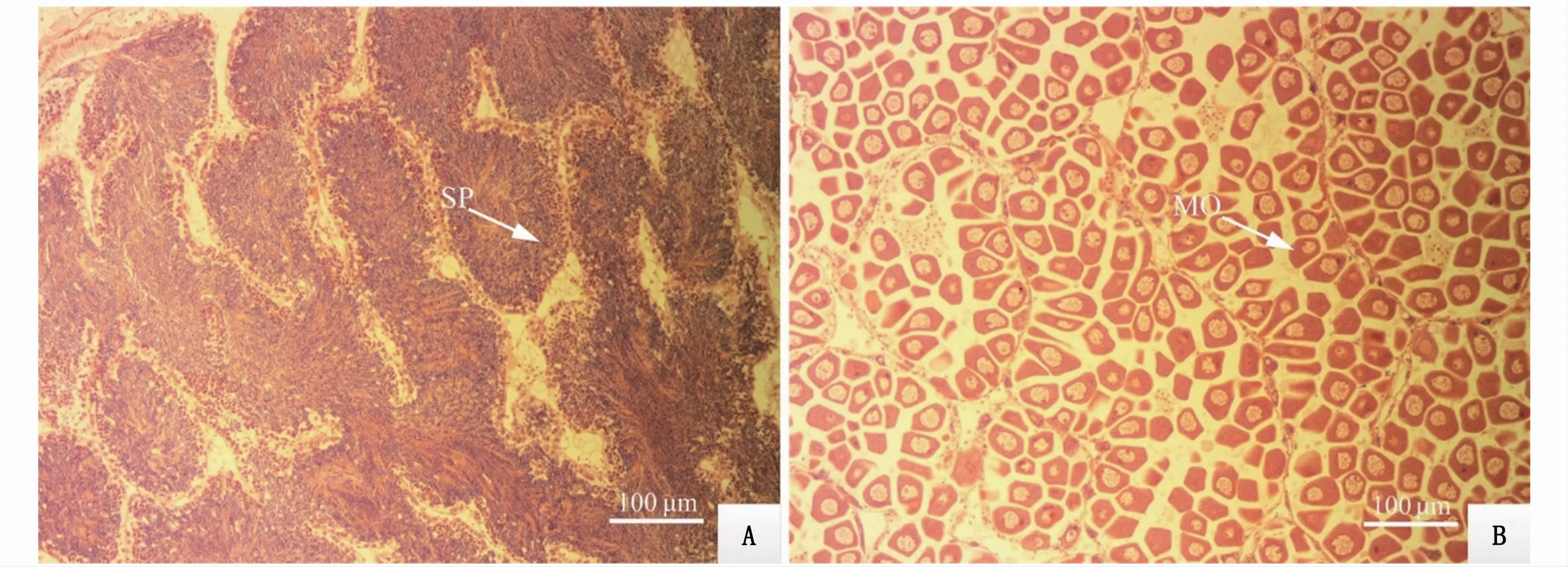
圖2 近江牡蠣育苗親本性腺切片顯微觀察(×10)Fig.2 Gonad section of parent C.ariakensis under m icroscope(×10)
2.2 近江牡蠣受精和早期發育過程
表1為近江牡蠣受精和早期發育過程中各個階段的時間。

表1 近江牡蠣受精和早期發育過程中各個階段的時間Tab.1 Time of insem ination and stages of early development for C.ariakensis
2.2.1 受精和早期卵裂過程
2.2.1.1 精子入卵過程
未受精的成熟卵細胞在未激活狀態下呈卵圓形或梨形,卵徑為50μm左右(圖3-A)。在熒光顯微鏡下可以看到,成熟卵細胞染色體呈亮藍色,卵質則為淺藍色,染色體整齊地排列在赤道板上(圖3-B);成熟精子頭部的染色質呈亮藍色熒光(圖3-C)。精子在和卵細胞混合后擺動尾巴至卵子周圍,通過頂體吸附在卵子表面(圖3-D)。受精后5~10 min,精子頭部在進入卵子細胞質后膨大成直徑為4~5μm的圓球形精核,此時第一次成熟分裂開始(圖3-E)。
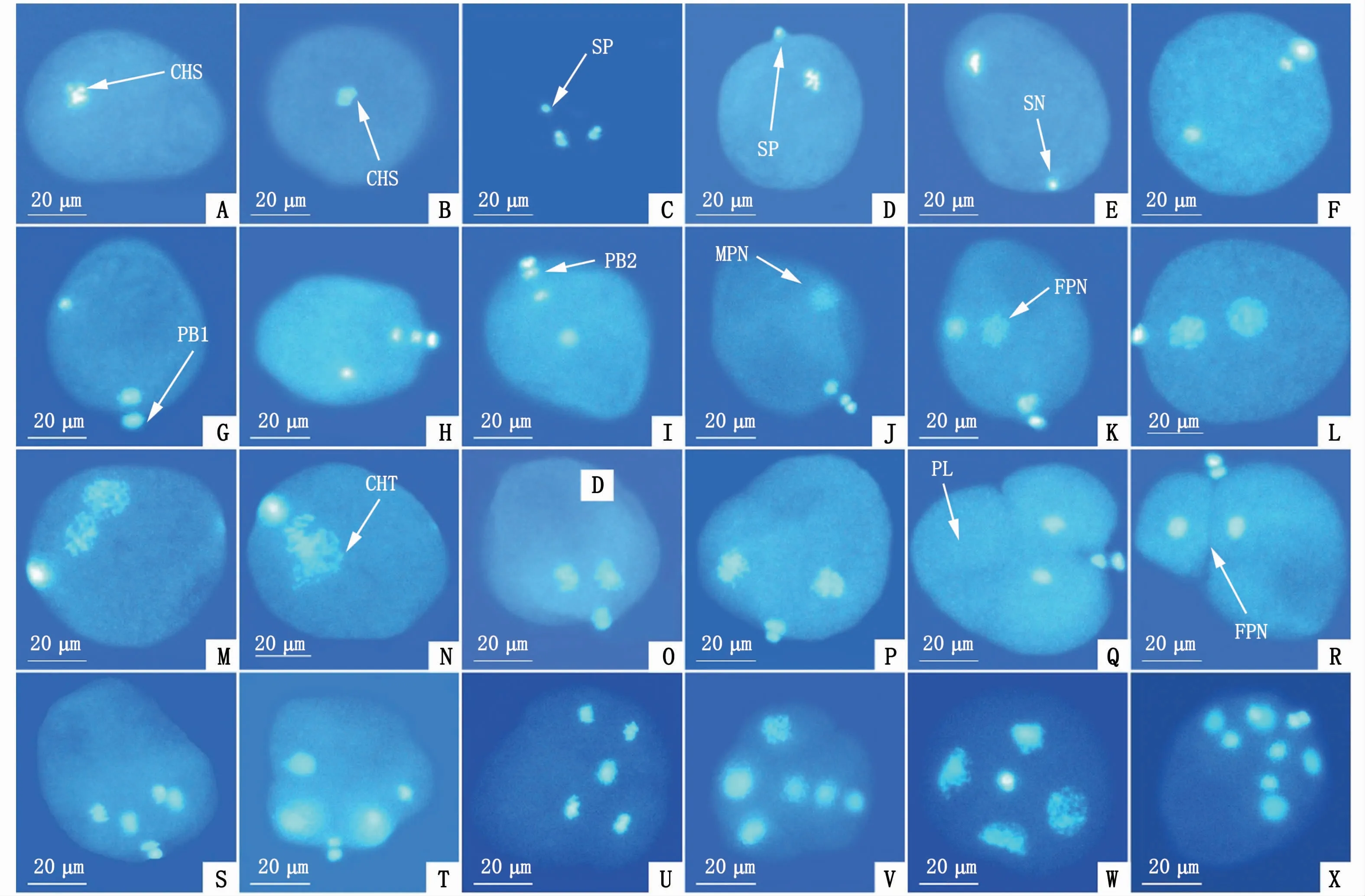
圖3 近江牡蠣受精和早期卵裂過程核行為的熒光顯微鏡觀察(×20)Fig.3 Nuclear behavior of fertilization and early cleavage in C.ariakensis under fluorescentm icroscope(×20)
2.2.1.2 兩次成熟分裂過程
卵子在受精后10~15 min,其內位于赤道板上的同源染色體會在兩側紡錘絲的牽引下向卵膜移動,此時處于第一次成熟分裂后期(圖3-F);受精后15~20 min,靠近卵膜一側的染色體組不斷向卵膜處移動,最終使卵膜拱起,第一極體形成并排出卵膜外,第一次成熟分裂完成(圖3-G)。受精后20~25 min,在卵子赤道板上出現新排列的染色體組,染色體組在紡錘絲的牽引下向卵膜和反方向移動,第二次成熟分裂啟動;受精后25~30 min,姐妹染色體分離后,卵膜一側的染色體組濃縮形成第二極體,并向卵外排出,完成第二次成熟分裂,該過程產生的核相變化如圖3-H,I所示。在整個過程中,精核位置、大小變化不明顯。
2.2.1.3 雌、雄原核的形成
近江牡蠣卵細胞在受精后30 min左右,精核開始膨脹,最終形成松散的團狀核物質,即雄原核,且雄原核體積遠大于精核(圖3-J)。核相卵子中的另一組染色體也開始膨脹形成松散的團狀核物質,即雌原核(圖3-K)。雌原核的位置靠近極體,它的形成要晚于雄原核。
2.2.1.4 第一次卵裂
受精后45 min左右,雌、雄原核相互靠近(圖3-L),并向卵子中間位置移動(圖3-M),但不融合,雌、雄原核染色絲凝集,呈現出兩個獨立的染色體組,并在赤道板上以染色體聯合的方式結合,此時處于第一次卵裂中期(圖3-N)。
然后,排列于赤道板上的染色體組在紡錘絲的牽引下向受精卵兩極移動,此時受精卵處于第一次卵裂后期(圖3-O)。與此同時,受精卵形狀也發生顯著變化,細胞向兩極橫向拉伸,極葉伸出(圖3-P),隨著染色體繼續分離,極葉突出,染色體處于兩個卵裂溝內,受精卵呈三葉狀,進入三葉期(圖3-Q)。
受精后60 min,極葉收縮,卵細胞自極體生成處形成一大一小2個卵裂球,其中大卵裂球上還可以看到未完全縮回的極葉,最終極葉全部回縮消失,卵裂溝清晰可見(圖3-R),此時第一次卵裂完成,受精卵處于2細胞時期(圖3-R)。
2.2.1.5 第二、三次卵裂
受精后75 min,2個卵裂球開始分裂(圖3-S),染色體組向兩極移動,至第二次卵裂中期分裂相形成(圖3-T),各自分裂出一大一小的卵裂球(圖3-U)。受精后1.5 h,卵裂球分裂基本結束,最終形成4個卵裂球,卵裂溝明顯,此時進入4細胞時期(圖3-V)。4個卵裂球內染色體逐漸變成染色質,并形成新的核膜,為第三次卵裂做準備。
2.2.1.6 多精入卵現象
在近江牡蠣受精發育過程中,觀察到一些多精入卵現象。在多精受精的卵子中,入卵精子數多在2~5個,甚至更多。當2~3個精子同時進入卵細胞,大多不影響后續的成熟分裂過程,但后期發育無法進行。而當入卵精子數目較多時,分裂就會被抑制。多個精子入卵后會形成精核,精核膨脹為雄原核,多個雄原核各自形成染色體組,同時與雌性原核進行染色體聯合,最終導致多個染色體組因無法平均分到兩個子細胞中,發生染色體分離紊亂,該現象被稱為多極分離(圖3-W,S)。
2.2.2 早期胚胎發育光學顯微鏡觀察
2.2.2.1 桑葚期、囊胚期和原腸期階段
在受精后6 h左右,受精卵經過多次有絲分裂進入桑葚期(圖4-E),此時細胞界限相較于2細胞期、4細胞期較為模糊,胚胎內部無空腔。受精后的8~10 h,胚胎發育進入囊胚期(圖4-F),胚胎上開始逐漸出現微小的纖毛,纖毛擺動時胚胎會有小幅的運動現象。隨后進入原腸期(圖4-G),胚胎的植物極細胞內陷,胚胎僅能作小范圍的順時針旋轉運動。
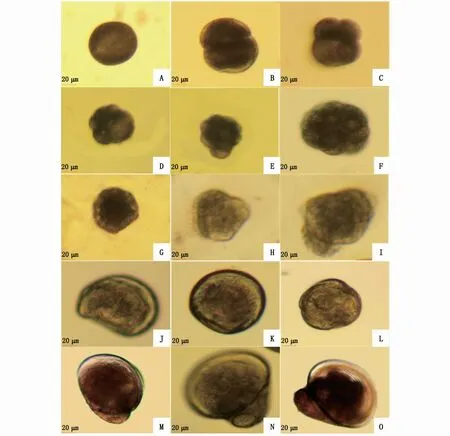
圖4 近江牡蠣早期胚胎和幼體發育普通光學顯微鏡觀察(×20)Fig.4 Development of early embryos and larvae of C.ariakensis under ordinary opticalm icroscope(×20)
2.2.2.2 擔輪幼蟲
受精后12~15 h,胚胎發育成擔輪幼蟲,擔輪幼蟲前期形狀近圓形,一側纖毛增多,通過纖毛擺動做回旋運動(圖4-H),隨后頂板逐漸變為橢圓形,生出的殼腺分泌物質形成殼,包裹在幼蟲外(圖4-I)。
2.2.2.3 D形幼蟲
受精后27~30 h,進入到D形幼蟲階段(圖4-J)。
2.2.2.4 殼頂幼蟲
經過5 d左右的發育,幼蟲發育至殼頂幼蟲階段,此時殼長達(82.40±4.48)μm。殼頂幼蟲前期,殼頂稍微突起(圖4-K),開始長出纖毛。當發育到殼頂幼蟲中期時,殼頂突出明顯,纖毛多而密;受精15 d左右進入殼頂幼蟲后期,此時幼蟲的殼頂突出更明顯,殼長已達到(143.20±5.11)μm(圖5),兩殼形狀大小不對稱,一側稍大于另一側(圖4-L)。

圖5 近江牡蠣幼蟲殼長生長情況Fig 5 Grow th in shell length of C.ariakensis larvae
2.2.2.5 眼點幼蟲
受精后17 d左右,部分幼蟲開始出現眼點,此時幼蟲發育至眼點幼蟲時期(圖4-M)。受精后20 d左右的眼點幼蟲殼長為(276.60±15.70)μm(圖5)。
2.2.2.6 附著幼蟲及稚貝
受精后22 d左右,胚胎發育至附著幼蟲時期,個體基本成型,開始營底棲爬行生活(圖4-N)。底棲后,附著幼蟲很快變態為附著稚貝(圖4-O)。到受精后的33 d時,幼蟲已大多變態為稚貝,殼長達(374.80±12.99)μm。
2.2.2.7 幼蟲殼長生長狀況
近江牡蠣幼蟲殼長在前10 d變化較小,10日齡幼蟲平均殼長為93.4μm,該時期的日平均生長率為3.16μm·d-1(圖5)。10~20 d幼蟲快速生長,其中15~20 d殼長變化最快,日平均增長率達26.68μm·d-1。20日齡的眼點幼蟲的平均殼長為276.60μm。20~30日齡幼蟲殼長增速放緩,日平均增長率為9.82μm·d-1。
3 討論
3.1 近江牡蠣受精細胞學特點
在雙殼貝類的受精過程中,根據受精發生時卵細胞所處的狀態,可將其大致分為兩類:第一類是,排放的卵子處于第一次成熟分裂前期或中期,受精后卵細胞的生發泡消失,而后再進行減數分裂,屬于該種受精方式的有大西洋浪蛤(Spisula solidissima)、寬殼全海筍(Barnea dilatata)、長牡蠣(C.gigas)等[19-21];另一類則是受精發生在第一次減數分裂中期,生發泡已破裂,精子入卵后減數分裂過程重新啟動,屬于這一類的有泥蚶(Tegillarcagranosa)、文蛤(Meretrix meretrix)、櫛孔扇貝(Chlamysfarreri)、皺紋盤鮑(Haliotisdiscushannai)、紫貽貝(Mytilusedulis)、合浦珠母貝(Pinctada martensii)和毛蚶(Scapharcasubcrenata)等[12,15,22-25]。本研究觀察發現,近江牡蠣屬于第一種類型,其未受精成熟卵子呈梨形,核相處于第一次成熟分裂中期,生發泡破裂發生在受精后,精子入卵前生發泡完整,精子入卵后分裂啟動。近江牡蠣精子入卵無固定位置,其是通過頂體反應在卵表面隨機入卵。實驗中還觀察到,近江牡蠣的卵子為均黃卵,在受精后才能區分具體的動植物極。
雌、雄原核形成的先后順序在不同雙殼貝類中存在一定的差異。在櫛孔扇貝、合浦珠母貝、毛蚶中,雄原核的形成要早于雌原核,而太平洋牡蠣(Crassostreagigas)的雌、雄原核幾乎是同時形成的[26]。本研究中,近江牡蠣的雄性原核形成早于雌性原核且比雌性原核的體積要大一些。雙殼貝類的雌、雄原核的結合方式也可分為兩種,一種是原核聯合的結合方式,另一種為原核融合的結合方式。本研究發現,近江牡蠣受精卵細胞雌、雄原核形成于第二次成熟分裂后期,兩原核相互靠攏時,有核膜存在,因此推斷其雌、雄原核結合方式為原核聯合。
3.2 多精入卵現象
雖然近江牡蠣本身具有防止多精入卵的機制,即精子入卵后,卵子會阻止其他精子附著,但本研究發現,近江牡蠣北方群體在溫度22~24℃、鹽度20~22的環境下進行人工授精的過程中,仍存在少數多精入卵的現象。多個精子進入卵子后,受精卵中的雄原核與雌原核會發生多個原核聯合,此時多個染色體組共存,細胞內同時形成多級紡錘體,導致卵細胞不能正常分裂,最終造成胚胎發育畸形乃至死亡。研究表明,雙殼貝類多精入卵現象與精卵質量、精卵濃度以及外界的受精環境因素等有很大關系[12]。近江牡蠣作為廣溫廣鹽性經濟貝類,人工育苗正處在溫度和鹽度變化較大的時期,因此開展溫度、鹽度對近江牡蠣影響的研究可以推動其人工繁育技術的發展。
3.3 近江牡蠣胚胎發育階段特點
在適宜溫度范圍內,不同品種的牡蠣幼蟲發育速度存在一定的差異。近江牡蠣在水溫22~24℃、鹽度20~22的環境條件下,受精卵經27~30 h發育到D形幼蟲,比太平洋牡蠣(22~23 h,20~22℃)、熊本牡蠣(C.sikamea)(17 h,27~28℃)、葡萄牙牡蠣(C.angulata)(16 h,25.3℃)、巖牡蠣(C.nippona)(18~20 h,22℃)發育到D形幼蟲所需的時間長[26-29]。在其他雙殼貝類中,幼蟲發育速度也不同,如縊蟶和小刀蟶(Phaxasattenuatus)在水溫為24~26℃環境中,經過24 h發育至D形幼蟲時期[30-31];青蛤在水溫28℃時,經16 h發育成D形幼蟲[30]。綜上所述,近江牡蠣的幼蟲發育速度低于大多數雙殼貝類。
眼點幼蟲作為幼蟲發育和附著變態的關鍵時期,因其生活水體環境復雜,可能受溫度、鹽度和餌料等因素的影響。如長牡蠣在20~22℃的條件下,經18 d幼蟲出現眼點,殼長為270~320 μm[24];熊本牡蠣在溫度26~27℃、鹽度28~30條件下,發育到16 d時,殼長達到320~330μm,有個別幼蟲出現眼點[27];巖牡蠣幼體在25℃下發育到14~16 d,殼長達到290~320μm,開始出現眼點[29];葡萄牙牡蠣幼蟲培育14~21 d,殼長達到310~330μm,開始出現眼點[28]。在本研究中,近江牡蠣在水溫22~24℃、鹽度20~22的環境條件下,經17 d左右幼蟲開始出現眼點,與上述貝類相似。推測貝類發育時間可能與育苗的溫度和鹽度有一定的關系。已有研究證明,鹽度和溫度均會影響牡蠣幼蟲的存活與生長,幼蟲存活率隨鹽度的升高而降低,而高溫會造成幼蟲的大量死亡,低溫則會影響幼蟲的生長[33-35]。在本研究中,近江牡蠣幼蟲培育前期較后期生長緩慢,這與呂曉燕[27]和劉魯雷[36]的研究結果類似,說明近江牡蠣在發育至眼點幼蟲時期后進入快速生長階段。
