
短枝木麻黃半同胞家系苗期遺傳變異及選擇
2023-09-20 11:23:02徐惠昌聶森余錦林尤龍輝葉功富張航曉程分生蘇親桂
熱帶作物學報 2023年8期
徐惠昌 聶森 余錦林 尤龍輝 葉功富 張航曉 程分生 蘇親桂
關鍵詞:短枝木麻黃;半同胞家系;遺傳變異;苗期選擇
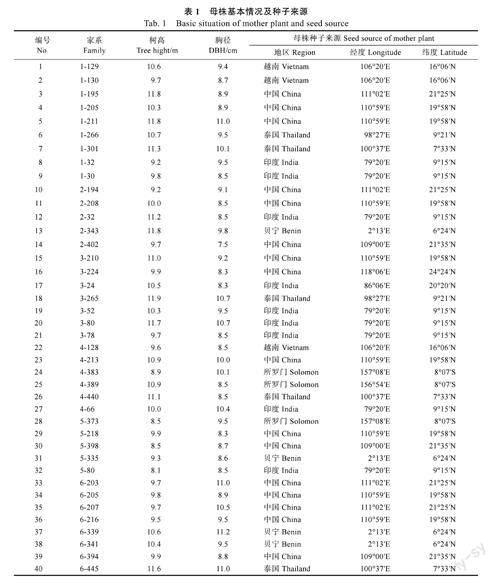
短枝木麻黃生長迅速,具有良好的抗風沙、耐旱及耐鹽堿等特性,是我國東南沿海沙地最重要的生態防護樹種之一[1-2]。20世紀80年代營造的木麻黃防護林,因林分逐漸衰退更新,而被短枝木麻黃優良無性系惠1、閩平2號、木麻黃粵501等無性系所替代,但是由于無性系品種來源單一、種質資源貧乏、遺傳基礎狹窄,導致林分抗性下降、易感病蟲害等問題逐漸凸顯,木麻黃防護林生態防護功能不斷下降[3-4]。因此,木麻黃實生種子苗所具有的生長期長、根系發達、抗性強等優點得到相關研究者、林業生產和管理單位重新審視和重視。
“有性選育,無性利用”是木麻黃遺傳改良的指導方針[5-6]。我國自20世紀80年代中期,以中澳、中法等國際合作項目為契機,先后引進短枝木麻黃不同地區國際種源73個[7],并于福建、廣東和海南等地建立了多個種源試驗點,從而揭開了國內短枝木麻黃種源試驗的序幕,并對其遺傳變異規律進行了較為全面、系統的研究,選育出了多用途的優良種源[2]。但是,關于短枝木麻黃優良家系遺傳選育的研究卻較為滯后。傅玉獅[8]于1996年,通過對34個短枝木麻黃半同胞家系的11年生子代測定林進行分析,結果表明其生長和抗性性狀遺傳變異豐富,并篩選出8個速生豐產的優良家系,其在一定程度上豐富了短枝木麻黃優良種質資源。然而,此后關于短枝木麻黃家系遺傳改良育種工作的研究鮮有報道。
亞群體層次(如家系層次)遺傳改良相較于群體層次,可以消除后者對有益配子不平衡的負面效應,并避免加性基因方差的下降,提高子代育種值,林業發達國家在采用現代苗木生產技術的同時,也正從種源層次的苗木培育逐步轉為家系層次育種,以充分發揮亞群體層次遺傳改良的潛在效益[9]。鑒于良種選育和苗木培育對發掘變異、研究變異、利用變異的要求,我國林木育種工作者對紅皮云杉[10]、云南松[11]等樹種進行了大量研究,并證實在苗期進行遺傳改良選擇,在提高林木良種選育效率、加速育種進程的同時,還可獲得較高的增益。故本研究以40個短枝木麻黃半同胞家系1年生幼苗為研究對象,結合前人對短枝木麻黃的相關研究[2,12],觀測分析其苗期苗高、地徑、干重、根冠比、側枝數、側枝角及側枝長等生長與分枝性狀的遺傳變異,探尋短枝木麻黃家系層次的苗期遺傳變異規律,以期為后續木麻黃科樹種更好地開展家系層次遺傳改良工作提供參考依據。
1材料與方法
1.1材料
1.1.1試驗地點試驗地位于福建省惠安赤湖國有防護林場后山工業區(118°54′29″E,24°54′45″N),平均海拔約12m。屬南亞熱帶海洋性季風氣候,干濕季明顯,夏季多臺風,秋冬盛行東北風,該區域年均氣溫為20.7℃,年均降雨量1029.0mm,年均蒸發量達2056.7mm。試驗地設在林場場部苗圃,基質土壤類型為沙壤土,肥力較低,日照充足。
1.1.2試驗材料2019年8月,對短枝木麻黃國際種源試驗林(5年生)的表型性狀進行觀測分析,篩選出樹高、胸徑生長勢及健康狀況良好的優良母株進行采種、保存。2021年3月取出種子,于林場苗圃進行播種育苗,共計培育40個短枝木麻黃半同胞家系(表1)。2021年5月,當幼苗長至10cm左右時,按照完全隨機區組設計,采用雙列10株小區排列方式,將幼苗移栽至塑料營養缽(規格為8cm×15cm),一株一缽,基質為沙壤土,共4個區組(重復)。適時進行澆水、除草等常規育苗管理。
1.2試驗指標觀測
2022年3月對短枝木麻黃40個半同胞家系幼苗的生長和分枝性狀進行測定。測定指標包括苗高(SH)、地徑(GD)、干重(WD)、根冠比(R/S)、側枝數(LBN)、側枝角(LBA)、側枝長(LBL)等7個指標。其中苗高、地徑分別采用卷尺和電子游標卡尺進行測量;側枝角和側枝長則采用量角器和直尺進行測量。根據測量結果,計算各區組每個家系平均苗高和平均地徑;各區組每個家系再隨機選取4株幼苗,清理根部泥土后,分根、莖、小枝帶回實驗室,置于鼓風干燥箱中,105℃烘至恒重并稱重,計算根冠比=地下部分干重/地上部分干重。
1.3數據處理
采用Excel2020和SPSS25.0軟件進行數據整理和統計分析;使用R語言程序包“HalfsibMS”計算遺傳參數[13];利用Origin2021進行聚類分析和增益柱狀圖的繪制。
1.3.1生長與分枝性狀遺傳力、遺傳變異系數、相關系數和增益的數學模型[14-15]
1.3.2短枝木麻黃優良家系的選擇通過聚類分析和布雷金多性狀綜合評價法開展優良家系的綜合選擇。布雷金多性狀綜合評價法數學模型為[16]:
2結果與分析
2.1生長與分枝性狀變異
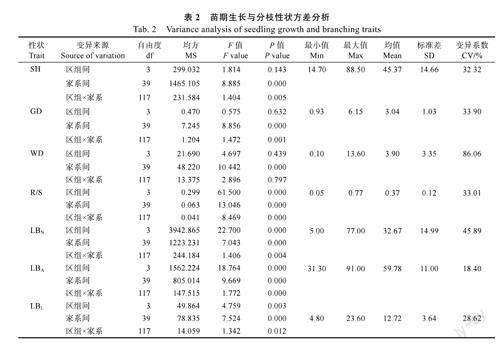
由表2可知,7個苗期性狀變異系數為18.40%~86.06%,表明參試群體生長與分枝性狀分化較明顯。方差分析結果則顯示,40個短枝木麻黃半同胞家系間苗期各生長和分枝性狀差異均極顯著(P<0.01,下同),且方差來源以家系為主。
2.2生長與分枝性狀的遺傳參數估算
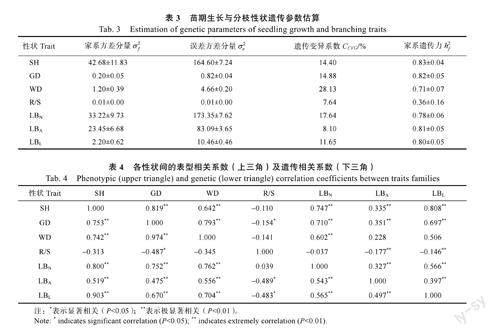
如表3所示,參試群體生長與分枝性狀的遺傳變異系數為7.64%~28.13%,其中根冠比和側枝角的遺傳變異系數均低于10%。在遺傳力方面,各表型性狀的家系遺傳力變化幅度為0.36~0.83。
2.3生長與分枝性狀的表型和遺傳相關分析
對短枝木麻黃半同胞家系苗期生長與分枝性狀,進行表型和遺傳相關分析,結果(表4)表明,根冠比、側枝角與其他表型性狀指標相關性不明顯或呈弱相關關系。苗高、地徑、干重、側枝數及側枝長相互間表型和遺傳相關性較高,且均達到極顯著水平。
2.4優良家系選擇
由于根冠比、側枝角的遺傳變異系數較低,因此,僅以苗高、地徑、干重、側枝數和側枝長這5個性狀作為評價指標,對參試家系進行聚類分析,劃分類群(圖1),再計算各類群的遺傳增益和現實增益。
圖1顯示,當歐式距離達到15時,可將參試家系劃分為4個類群:其中第Ⅰ類群包含1-129、2-402、2-32等10個家系;第Ⅱ類群則包含1-211、3-80、1-32和3-224等4個家系;第Ⅲ類群包含1-130、1-30、2-194等15個家系;第Ⅳ類群則包含1-205、4-128、2-343等11個家系。圖2顯示,第Ⅰ、Ⅱ類群家系苗期生長和分枝性狀的現實增益和遺傳增益均為負值,因此將其進一步歸為苗期慢生家系;第Ⅲ類群家系除干重的現實增益和遺傳增益高于10%,其余4個性狀的現實增益為2.01%~7.86%,遺傳增益為1.57%~6.44%,將其歸為苗期較速生家系;第Ⅳ類群家系與參試群體相比,其側枝長的現實增益和遺傳增益分別為6.28%和5.02%,而苗高、地徑、干重和側枝數的現實增益和遺傳增益均超過10%,可歸為苗期速生家系。
以苗高、地徑、干重、側枝數及側枝長為評價指標,采用布雷金多性狀綜合評價法對參試家系進行評價(表5)。按10%的入選率進行篩選,共有4個家系入選,分別為3-265、5-373、1-266和2-343家系,其苗高、地徑、干重、側枝數和側枝長平均值分別為54.59cm、3.61mm、5.48g、39.04和15.10cm,現實增益分別為21.14%、19.10%、41.83%、19.97%和18.88%,遺傳增益分別為17.55%、15.66%、29.70%、15.57%和15.10%。
3討論
遺傳和變異是林木遺傳改良的重要基礎[17]。遺傳力表示親本將某性狀遺傳給子代的能力,遺傳力愈大,愈能穩定遺傳給子代[18-19]。本研究中,苗高、地徑、根冠比、側枝數、側枝角及側枝長等性狀的家系遺傳力為0.36~0.83,在家系水平上,受到中度或強度遺傳控制[20],顯著高于PRASAD等[21]在印度安得拉邦對短枝木麻黃家系苗期生長性狀遺傳力的估算結果。這可能是因為短枝木麻黃花期以3—5月為主,而受西南暖濕氣流與冷空氣交綏作用的影響,供試樣地此時降雨較為頻繁,在一定程度上阻礙了花粉傳播,同一母株的種子材料中可能包含較大比例的全同胞子代,間接導致參試家系的遺傳力有所提高[22-24]。此外,方差分析結果顯示,各性狀在家系間差異極顯著,變異系數為18.40%~86.06%,這與胡盼等[12]對幼苗時期不同種源的短枝木麻黃表型性狀差異評價結果及陳珍等[25]對25個短枝木麻黃無性系進行苗期選擇研究的結果相似,表明短枝木麻黃生長和分枝性狀分化明顯,變異較豐富,具有較高的遺傳改良潛力。
遺傳變異系數反映了性狀遺傳變異的相對大小,其數值越大,選取獲得優良遺傳型的潛力越大[26]。本研究中,參試群體根冠比和側枝角的遺傳變異系數較低,僅為7.64%和8.10%,表明這2個性狀的選擇潛力較低。相關系數代表了性狀間的關聯程度,可用來確定目的性狀之間的相關關系[27-28]。本研究發現,側枝數、側枝長與苗高、地徑、干重間的表型相關性和遺傳相關性均達到了極顯著水平,這與胡盼等[12]的研究結果相似,說明側枝數和側枝長可作為短枝木麻黃優良半同胞家系篩選的參考因子。
遺傳增益是人工選擇群體所取得改良效果的重要評價指標,受到遺傳力、變異系數和選擇強度的直接影響[24]。本研究以苗高、地徑、干重、側枝數和側枝長作為評價指標,結合聚類分析和多性狀綜合評價法,以10%入選率,篩選出4個優良家系,增益明顯,選擇效果較為顯著。然而,隸屬于較速生類群的1-266入選其中,可能是因為該家系平均側枝長較大,因此,在苗期選擇時,也可適當考慮第Ⅲ類群的較速生家系。入選優良家系為后續短枝木麻黃遺傳改良和林分營建提供了較好的種質材料。但受基因與環境的共同作用,林木在不同生長發育階段,其遺傳和表型參數是動態變化的[29],故今后還需結合造林試驗進行多年觀測,進一步探究短枝木麻黃半同胞家系的變異及其穩定性,以便更加科學地指導木麻黃育種和營林生產實踐。
