不同放牧模式對(duì)祁連山高寒草甸植物群落特征的影響
2023-10-08 08:57:00于澤航董全民張春平劉文亭楊曉霞劉玉禎張振祥呂衛(wèi)東楊增增
草地學(xué)報(bào) 2023年9期
于澤航, 董全民*, 曹 銓, 俞 旸, 張春平, 劉文亭,楊曉霞, 劉玉禎, 張振祥, 馮 斌, 呂衛(wèi)東, 楊增增
(1.青海大學(xué)畜牧獸醫(yī)科學(xué)院, 青海 西寧 810016; 2.青海省高寒草地適應(yīng)性管理重點(diǎn)實(shí)驗(yàn)室, 青海 西寧 810016;3.三江源區(qū)高寒草地生態(tài)教育部重點(diǎn)實(shí)驗(yàn)室, 青海 西寧 810016)
高寒草甸作為青藏高原廣泛分布的草地類型,具有涵養(yǎng)水源、碳固持、生物多樣性保護(hù)和緩減氣候變化等多種生態(tài)功能[1-3],對(duì)維持區(qū)域經(jīng)濟(jì)發(fā)展和生態(tài)平衡具有重要作用[4-5]。上世紀(jì)末,隨著全球氣候變暖、高寒牧區(qū)人口激增、人為活動(dòng)加劇和過(guò)度放牧,青藏高原高寒草甸出現(xiàn)不同程度的退化[6-7]。過(guò)度放牧是造成高寒草甸退化的重要因素之一,該方式破壞了植物組織和功能,減弱了植株再生能力,降低了植物生物量,加之牧民不予管理的態(tài)度,高寒草甸進(jìn)一步退化[8-12]。
草甸植物群落的穩(wěn)定性和生產(chǎn)力是植被對(duì)環(huán)境變化響應(yīng)的指示器,能充分反映植被對(duì)于環(huán)境和人類干擾的變化特征[13]。近年來(lái),我國(guó)學(xué)者針對(duì)放牧導(dǎo)致高寒草甸持續(xù)退化的問(wèn)題,通過(guò)比較放牧強(qiáng)度、放牧制度及放牧季節(jié)等管理方式,探討了放牧對(duì)高寒草甸植物群落特征的影響。王向濤等[14]基于放牧強(qiáng)度的研究發(fā)現(xiàn),草甸地上總生物量、物種豐富度在圍封樣地中最高,而可食牧草的生物量和蓋度在中度放牧中最高。劉玉禎等[15]關(guān)于放牧方式的研究表明,在基于中度強(qiáng)度放牧條件下,藏羊、牦牛1∶2混合放牧對(duì)草地凈初級(jí)生產(chǎn)力和家畜采食量最高。不同放牧管理方式對(duì)植物群落物種產(chǎn)生的影響不同,而物種又是組成群落的基本單位,物種的變化直接影響植物群落的變化,不同的物種在植物群落生態(tài)恢復(fù)過(guò)程中并非同等重要,往往關(guān)鍵種的損失會(huì)導(dǎo)致生態(tài)系統(tǒng)更大甚至不可逆的改變,且關(guān)鍵種在生態(tài)系統(tǒng)中發(fā)揮功能的重要性遠(yuǎn)大于其它物種[16]。關(guān)鍵種的確定有移除法和對(duì)比法,這2種方法對(duì)關(guān)鍵種的確定存在諸多主觀因素,限制性較強(qiáng),其準(zhǔn)確性也有待商榷。網(wǎng)絡(luò)分析廣泛應(yīng)用于微生物群落中關(guān)鍵種的識(shí)別,Zhang等[17]利用網(wǎng)絡(luò)分析評(píng)價(jià)不同圍封年限對(duì)高寒草甸植物群落結(jié)構(gòu)及穩(wěn)定性的影響中發(fā)現(xiàn),基于物種重要值構(gòu)建的網(wǎng)絡(luò)圖最具代表性,為確定植物群落的關(guān)鍵種提供了新方法,本研究植物群落關(guān)鍵種計(jì)算也使用此方法。綜上所述,系統(tǒng)評(píng)價(jià)植物群落特征應(yīng)從不同方面分析,多方面的研究結(jié)果更有利于高寒草甸植物群落穩(wěn)定和健康發(fā)展。
本研究在青海省祁連縣野牛溝鄉(xiāng)開(kāi)展試驗(yàn),通過(guò)設(shè)置不同的牦牛放牧模式(放牧、放牧+補(bǔ)飼、圍封),系統(tǒng)性分析植物群落結(jié)構(gòu)、關(guān)鍵種及功能群特征,旨在明確不同放牧模式對(duì)高寒草甸植物群落特征的影響,以期為祁連山冬季草場(chǎng)的科學(xué)合理利用提供理論依據(jù)。
1 材料與方法
1.1 研究區(qū)概況
研究區(qū)位于青海省海北藏族自治州祁連縣野牛溝鄉(xiāng)(38°39′N,99°19′E),平均海拔3 300 m左右,年均溫—1.3℃左右,年降水約在420 mm,降雨多集中于5—9月,氣候類型屬于典型的高原大陸性氣候。全年無(wú)絕對(duì)無(wú)霜期,多以冷暖區(qū)分,冷季平均氣溫—25.6℃,暖季平均氣溫13.4℃,年均日照時(shí)數(shù)約2 900 h。研究區(qū)草地類型為高寒草甸,土壤類型為高山草甸土[18],研究點(diǎn)的優(yōu)勢(shì)物種有西北針茅(Stipasareptana)、矮生嵩草(Carexalatauensis)、洽草(Koeleriamacrantha)、珠芽蓼(Bistortavivipara)、雞冠茶(Sibbaldianthebifurca)、多莖委陵菜(Potentillamulticaulis)等。
1.2 研究方法
1.2.1樣地設(shè)置 于2019年5月,通過(guò)對(duì)當(dāng)?shù)啬羺^(qū)進(jìn)行實(shí)地走訪和調(diào)查,選取地形一致且植被特征較均一的冬季草場(chǎng)試驗(yàn)樣地,放牧?xí)r間為每年11月—次年5月,其余時(shí)間禁牧,以牦牛為研究家畜,牦牛體重為(110±5) kg,樣地設(shè)置3種不同放牧模式:放牧(M1)、放牧+補(bǔ)飼(M2)以及圍封(M3)。放牧(0.35 AU·hm-2)指冬季草場(chǎng)連續(xù)放牧,不進(jìn)行補(bǔ)飼;放牧+補(bǔ)飼(0.38 AU·hm-2)指冬季草場(chǎng)連續(xù)放牧,同時(shí)每頭牛每日補(bǔ)飼1.2 kg干草;圍封地禁牧不利用。
1.2.2植物樣品采集方法 植被群落特征的測(cè)定是在2022年8月牧草生長(zhǎng)期進(jìn)行,在不同放牧模式的樣地中以樣帶的方式進(jìn)行取樣,每個(gè)處理設(shè)置3條相距100 m樣帶,每條樣帶等距設(shè)置3個(gè)取樣點(diǎn),采用50 cm×50 cm的樣方進(jìn)行,分種測(cè)定了各物種的蓋度、高度(每個(gè)樣方內(nèi)記錄5株,不足5株的按實(shí)際株數(shù)記錄)、頻度、密度,并對(duì)牧草分種齊地面進(jìn)行刈割裝入信封,根系采集利用內(nèi)徑5 cm的土鉆采集0~10 cm土層土樣,將根系清洗干凈后裝入信封,最后把地上植物和根系帶回實(shí)驗(yàn)室在烘箱中65℃恒溫烘至恒重。
1.3 數(shù)據(jù)處理與分析
物種重要值(Important value,IV)和多樣性采用以下公式計(jì)算[19]:
物種重要值(IV):IV=(Hr+Br+Dr)/3
(1)
Patrick豐富度指數(shù)(R):R=S
(2)
Shannon多樣性指數(shù)(H):H=-∑(VilnVi)
(3)
Pielou均勻度指數(shù)(D):D=H/lnS
(4)
式中:Hr為相對(duì)高度;Br為相對(duì)物種生物量;Dr為相對(duì)頻度;S為物種數(shù);Vi為物種i的重要值。采用單因素方差分析(One-way ANOVA)不同放牧模式下植物群落特征指標(biāo)的變化。
其次,應(yīng)用網(wǎng)絡(luò)分析(Network analysis)確定植物群落關(guān)鍵種。關(guān)鍵種確定分為3個(gè)步驟[20]:計(jì)算每個(gè)物種的重要值(IV)[21];基于物種重要值計(jì)算物種相互間的Pearson相關(guān)系數(shù)r,用于構(gòu)建網(wǎng)絡(luò)圖的邊[22];把物種當(dāng)作節(jié)點(diǎn),將相關(guān)性系數(shù)r絕對(duì)值小于0.8的邊且顯著性水平大于0.05的值剔除。根據(jù)上述的節(jié)點(diǎn)和邊,利用Gephi 0.9.2制作不同放牧模式下的無(wú)向隨機(jī)網(wǎng)絡(luò)圖。在網(wǎng)絡(luò)圖中,特征向量中心度(Hub value)的大小能夠代表物種(節(jié)點(diǎn))在網(wǎng)絡(luò)中的重要性[23],在網(wǎng)絡(luò)圖中節(jié)點(diǎn)最大的即為該模式下植被群落的關(guān)鍵種[24]。平均連通度(Average connectivity)來(lái)權(quán)衡物種間的競(jìng)爭(zhēng)關(guān)系,數(shù)值越高說(shuō)明該群落競(jìng)爭(zhēng)越強(qiáng)、復(fù)雜性越高[25],平均聚類系數(shù)(Average clustering coefficient)來(lái)權(quán)衡植被群落的組織程度,數(shù)值越高說(shuō)明該群落的組織程度越高[26]。
以上全部統(tǒng)計(jì)、繪圖都在R 4.0.3中進(jìn)行。
2 結(jié)果與分析
2.1 不同放牧模式對(duì)植物群落結(jié)構(gòu)的影響
由圖1可知,M2,M3模式下植物地上生物量和高度顯著高于M1(P<0.05),但M2,M3之間無(wú)顯著差異;M3模式下植物地下生物量和蓋度顯著高于M1,M2(P<0.05);M1,M2,M3模式下植物豐富度差異顯著(P<0.05);M1,M2,M3模式下植物群落的Shannon-Wiener指數(shù)和Pielou均勻度指數(shù)無(wú)顯著差異。
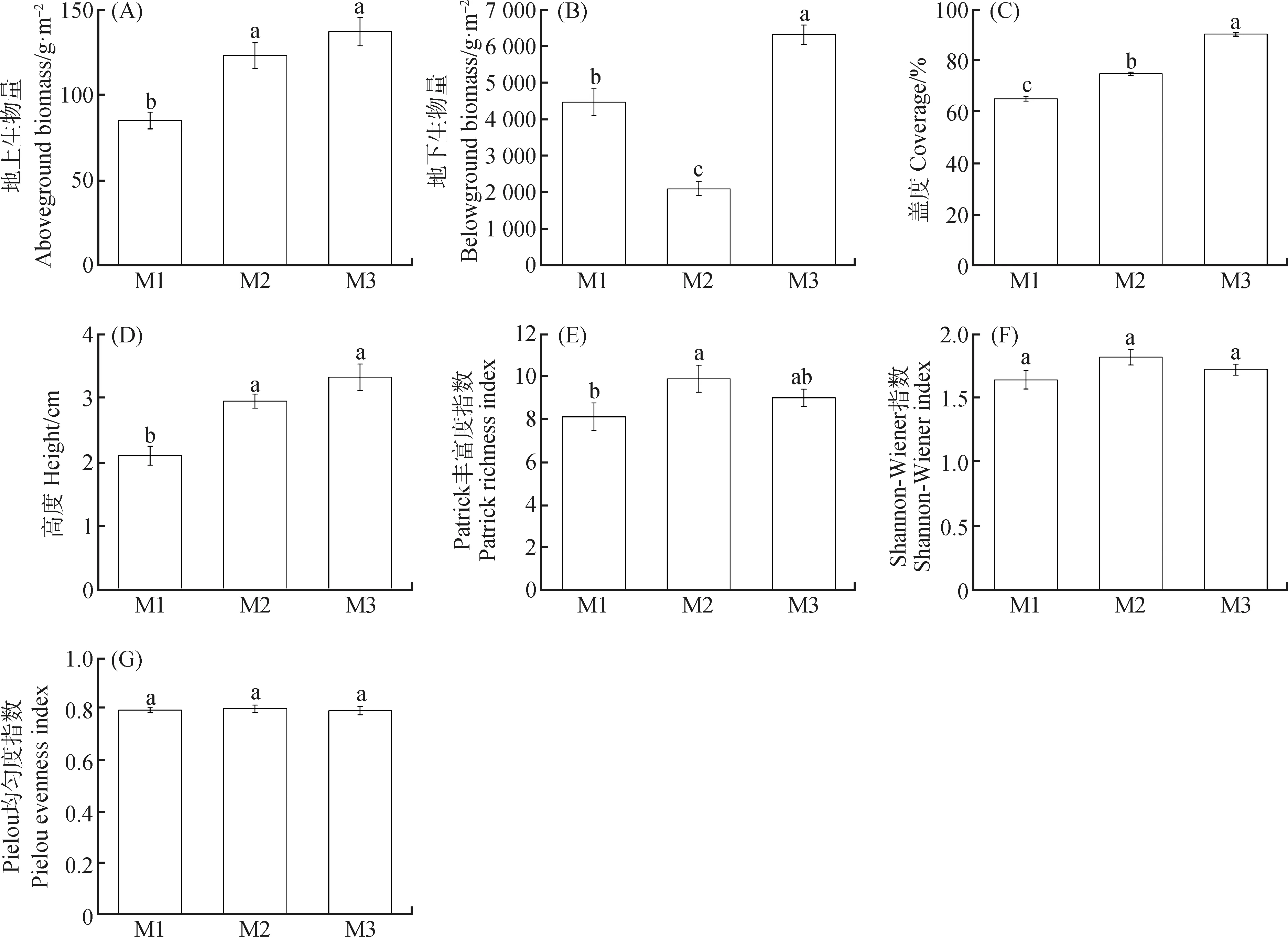
圖1 不同放牧模式對(duì)植物群落結(jié)構(gòu)的影響
2.2 不同放牧模式對(duì)群落網(wǎng)絡(luò)結(jié)構(gòu)及關(guān)鍵種的影響
由圖2A可知,M1模式下的植物群落網(wǎng)絡(luò)圖中有13個(gè)節(jié)點(diǎn)26條邊,植物群落關(guān)聯(lián)性較差;M2模式下的植物群落網(wǎng)絡(luò)圖中有18個(gè)節(jié)點(diǎn)28條邊,呈現(xiàn)出較好的植物群落關(guān)聯(lián)性;M3的植物群落網(wǎng)絡(luò)圖中有16個(gè)節(jié)點(diǎn)23條邊,植物群落關(guān)聯(lián)性中等。M1,M2,M3放牧模式下關(guān)鍵種依次為雞冠茶、洽草、多莖委陵菜(圖2B)。
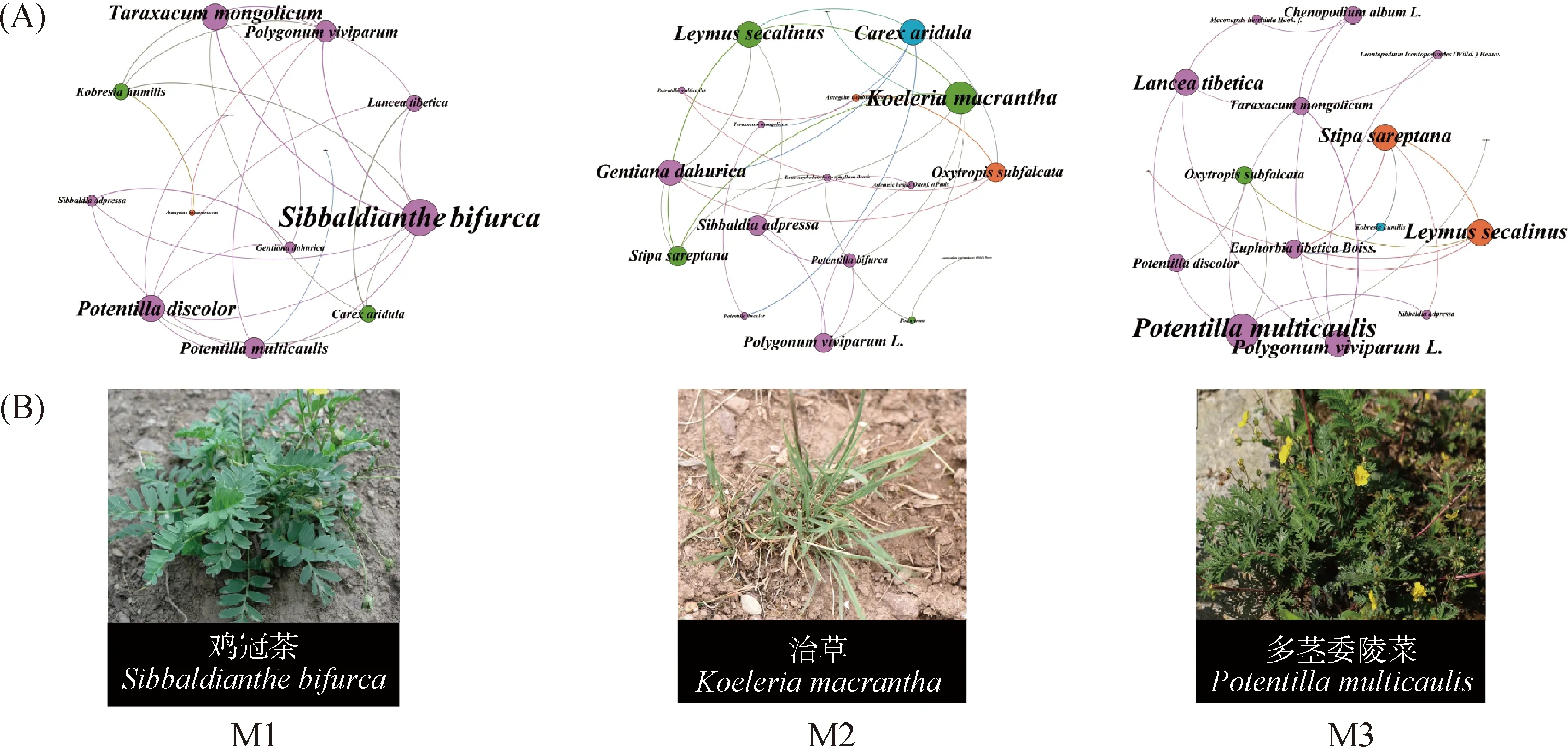
圖2 不同放牧模式下的植被群落網(wǎng)絡(luò)圖
由圖3可知,M2模式具有最高的平均連通度,表明群落物種間競(jìng)爭(zhēng)平衡性、復(fù)雜度最高;M3的平均連通度次之,M1的平均連通度最差。不同放牧模式下的平均聚類系數(shù)排序?yàn)?M2>M1>M3,表明在放牧?xí)r添加補(bǔ)飼能夠維持植被群落物種間競(jìng)爭(zhēng)力和復(fù)雜度。
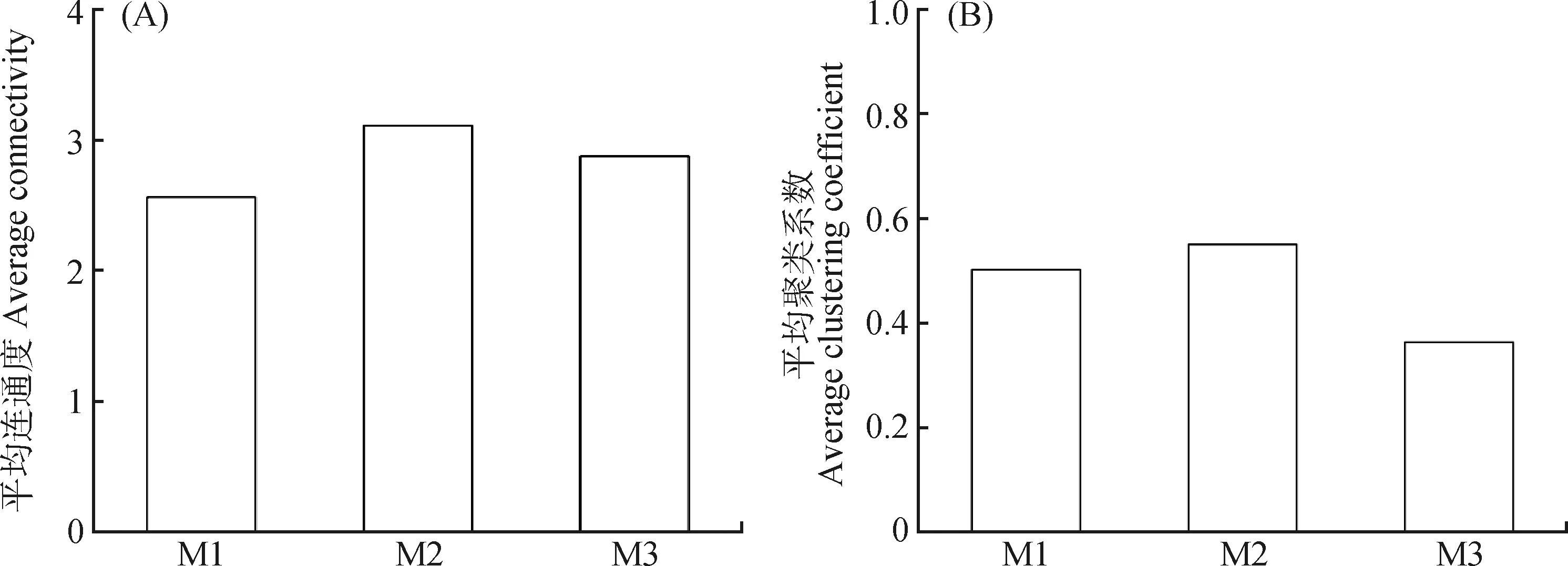
圖3 不同放牧模式下平均連通度和平均聚類系數(shù)的變化
由圖4可知,M1模式的擬合程度最高,群落間的連通度和Hub值較聚集,物種間的競(jìng)爭(zhēng)強(qiáng),復(fù)雜性高,因此,群落穩(wěn)定性高,關(guān)鍵種更替速度慢;M2模式擬合度處于M1和M3之間,物種競(jìng)爭(zhēng)和穩(wěn)定性相對(duì)較平衡;M3模式的擬合程度最低,群落間的連通度和Hub值差異較分散,物種間競(jìng)爭(zhēng)弱,復(fù)雜性低,使得群落穩(wěn)定性低,從而關(guān)鍵種更替速度快。

圖4 不同放牧模式下平均連通度與特征向量中心的關(guān)系
2.3 不同放牧模式對(duì)植物功能群特征的影響
M2和M3模式下禾本科的生物量高于M1模式,M1模式下莎草科的生物量高于M2,M3,而豆科和雜類草的生物量在3種模式下差異不大(圖5A)。
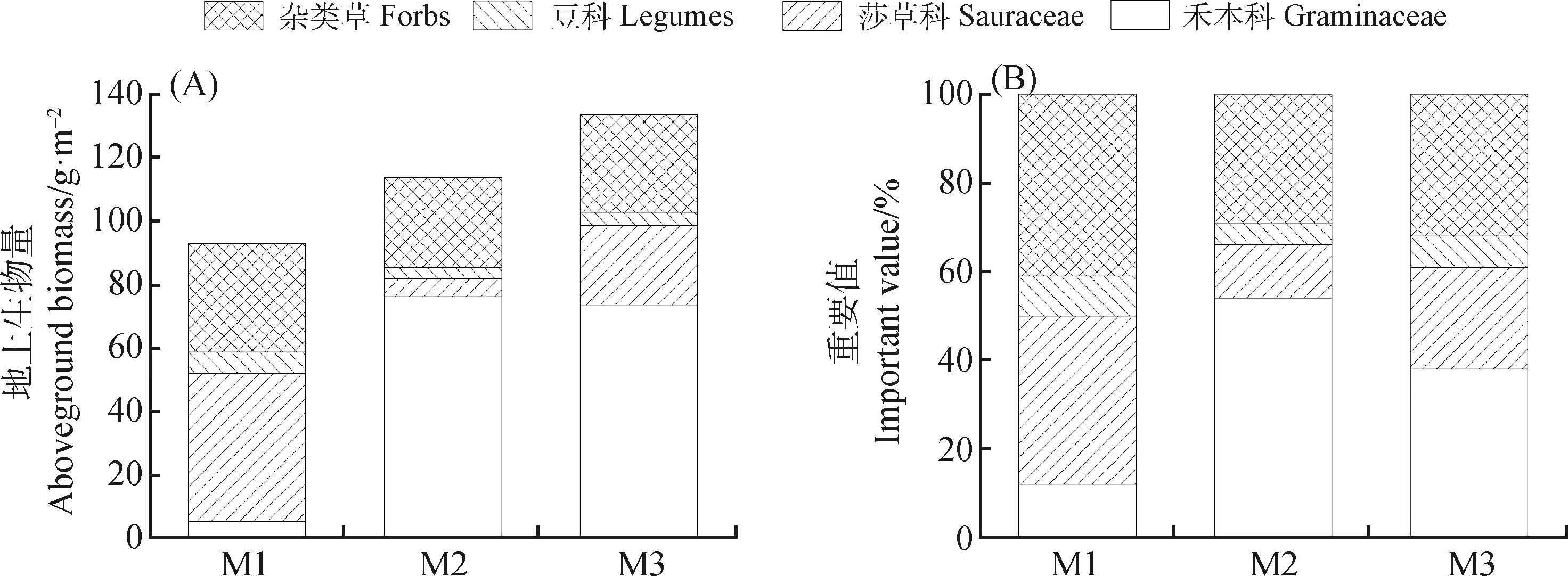
圖5 不同放牧模式下植物功能群特征
不同放牧模式下,各植物功能群占比不同(圖5B)。禾本科功能群重要值呈M2>M3>M1的趨勢(shì)變化,莎草科、豆科和雜類草功能群重要值呈M1>M3>M2的趨勢(shì)變化。
3 討論
3.1 不同放牧模式對(duì)植被群落結(jié)構(gòu)的影響
植被群落結(jié)構(gòu)是評(píng)價(jià)高寒草甸生態(tài)系統(tǒng)健康的重要指標(biāo),其變化過(guò)程與變幅顯著影響高寒草甸生態(tài)系統(tǒng)功能與生物多樣性[27]。本研究發(fā)現(xiàn),不同放牧模式會(huì)顯著改變植被的地上生物量、高度和蓋度。連續(xù)放牧導(dǎo)致群落蓋度、高度、生物量和物種數(shù)減少,且可食牧草占比減少,進(jìn)一步降低了高寒草甸植物群落的穩(wěn)定性,這和唐劉燕等[28]的研究結(jié)果相似。一方面由于家畜的放牧,使得植物光合組織和生長(zhǎng)組織長(zhǎng)期被家畜采食,牧草結(jié)構(gòu)被反復(fù)損傷,導(dǎo)致牧草的生長(zhǎng)速率變緩,受損傷的植被組織易遭受病菌侵染,增大了牧草的死亡率。此外,放牧還導(dǎo)致牧草的繁殖組織被轉(zhuǎn)移,牧草繁殖受到影響,進(jìn)一步延緩了牧草的繁殖期[29]。另一方面可能是資源分配權(quán)衡的作用[30],當(dāng)嚴(yán)重放牧脅迫到植物時(shí),其更傾向于避牧防御而不是非補(bǔ)償生長(zhǎng),會(huì)把更多的資源分配到地下部分來(lái)促進(jìn)根系的生長(zhǎng)。從地下生物量角度分析,M1模式下,牦牛不間斷的踐踏和采食,使得植被補(bǔ)償性生長(zhǎng)持續(xù)進(jìn)行,促進(jìn)了根系的生長(zhǎng);M2模式下,植被采食減少,更多的資源向地上部分聚集;M3模式下由于不受家畜干擾,土壤緊實(shí)度下降,土壤養(yǎng)分逐步改善,也促進(jìn)了地下生物量的積累,這一結(jié)果與張曉艷等[31]和趙景學(xué)等[32]的研究相似。植被群落物種的多樣性和豐富度是群落的重要性質(zhì),其對(duì)不同的環(huán)境異常敏感,容易受外界環(huán)境影響發(fā)生改變[33]。本研究發(fā)現(xiàn),三種模式下的物種豐富度有著顯著的差異,而Shannon-Wiener指數(shù)和Pielou指數(shù)沒(méi)有顯著差異。在M2模式下,物種的豐富度最高。依據(jù)植物群落構(gòu)成過(guò)程及過(guò)濾機(jī)制(Community assembiy and filtering mechanism),在生存環(huán)境空間相對(duì)較好的情況下,由于物種競(jìng)爭(zhēng)時(shí)間漫長(zhǎng)及存在短暫的排斥現(xiàn)象,加入適當(dāng)放牧干擾,能刺激植株再生,促進(jìn)植被補(bǔ)償生長(zhǎng),使群落的物種數(shù)增多[34]。
3.2 不同放牧模式對(duì)關(guān)鍵種及穩(wěn)定性的影響
關(guān)鍵種對(duì)支撐草地生態(tài)系統(tǒng)結(jié)構(gòu)有著重要的作用,倘若關(guān)鍵種丟失,生態(tài)系統(tǒng)結(jié)構(gòu)會(huì)發(fā)生巨大改變[35]。關(guān)鍵種的存在削弱了物種之間的競(jìng)爭(zhēng),進(jìn)而維持了生態(tài)系統(tǒng)的穩(wěn)定性。關(guān)鍵種在維持生態(tài)系統(tǒng)穩(wěn)定性的同時(shí),進(jìn)一步影響其他物種的分布與數(shù)量,最終決定了植物群落的物種多樣性、穩(wěn)定性和其它一連貫的生態(tài)效益[36]。本研究中,M1和M3模式下的關(guān)鍵種都為雜類草,其耐牧性和耐干擾性相對(duì)較高,可以有效延緩高寒草甸退化[37],但由于M1模式下長(zhǎng)時(shí)間的放牧導(dǎo)致了禾本科牧草減少,牧草質(zhì)量下降,進(jìn)而高寒草甸群落發(fā)生了改變。在M2模式下,由于補(bǔ)飼效應(yīng)的存在,降低了牦牛對(duì)牧草的采食率,進(jìn)而對(duì)草地植物群落結(jié)構(gòu)產(chǎn)生一定的調(diào)節(jié)作用[38]。Newman等[39]研究發(fā)現(xiàn),饑餓與飽腹?fàn)顟B(tài)下,不僅家畜的采食時(shí)間和采食率有差異,而且牧草中禾本科植物比例也有較大變化,這是由于不同狀態(tài)下家畜的采食量不同。由于M2模式下存在補(bǔ)飼效應(yīng),相對(duì)減少了家畜對(duì)禾本科植物的采食,使得長(zhǎng)勢(shì)較高的禾本科植物獲得更多的光能資源,在物種競(jìng)爭(zhēng)中占據(jù)優(yōu)勢(shì)[40-41]。從植被群落的平均連通度和平均聚類系數(shù)分析,M2模式下植被群落物種競(jìng)爭(zhēng)力和復(fù)雜度較高,這是由于在適當(dāng)放牧和補(bǔ)飼下,物種的互作強(qiáng)度提高,這與Gao等[42]的研究較為一致。
3.3 不同放牧模式對(duì)植物功能群特征的影響
草甸植被群落中功能群生物量及占比可直接反映草地的分布格局,不同放牧模式下的地上生物量占比可作為判斷草地狀況、生產(chǎn)力和演替趨勢(shì)的重要指標(biāo)[43]。放牧過(guò)程中,家畜采食和踐踏直接作用于草地,草甸土壤發(fā)生了變化,從而嚴(yán)重干擾植物的生長(zhǎng)發(fā)育,導(dǎo)致植物群落的高度和蓋度下降,生物量大幅度減少,其中禾本科牧草的生物量變化最為顯著[44-45]。不同放牧模式下的禾本科植物功能群生物量及重要值發(fā)生顯著變化,禾本科絕大多植物屬于家畜喜食的牧草,放牧過(guò)程中,其被家畜大量采食,使得禾本科牧草生物量大幅降低,這與張娜等[46]的研究結(jié)果一致。M1模式下的莎草科功能群占比最大,這是由于家畜優(yōu)先采食上層優(yōu)質(zhì)牧草后,從而使得作為下層植物的莎草科牧草獲得了較多的光熱資源,使其在群落功能群占比提高[47-48];M2模式由于存在補(bǔ)飼效應(yīng),家畜采食率降低,禾本科植物高大直立的特征使其獲得更多的光合優(yōu)勢(shì),冠層低矮的莎草科植物得不到更好的生長(zhǎng),所以在該模式下占比最低。在M1模式下豆科和雜類草植物重要值最高,因?yàn)檫m度放牧刺激植物產(chǎn)生補(bǔ)償生長(zhǎng),未被采食的植物光合效率增大,增強(qiáng)了植物的適應(yīng)能力,使得在群落中占比最高。M3模式下植物功能群組成發(fā)生改變,禾本科和莎草科植物增多。原因是消除了家畜對(duì)植物的采食和踐踏作用,禾本科植物和莎草科植物因其形態(tài)特征高大,較其它植物擁有更充足的環(huán)境優(yōu)勢(shì),從而生長(zhǎng)較快,在群落中占據(jù)主要地位,這與周華坤等[49]研究相似。
4 結(jié)論
通過(guò)對(duì)高寒草甸冬季牧場(chǎng)的不同利用發(fā)現(xiàn),草甸植物群落的物種組成和結(jié)構(gòu)發(fā)生了變化,但對(duì)植物群落的Shannon-Wiener指數(shù)和Pielou均勻度指數(shù)無(wú)顯著影響。“放牧”“放牧+補(bǔ)飼”“圍封”的關(guān)鍵種分別為雞冠茶、洽草和多莖委陵菜。從植物功能群生物量分配角度來(lái)看,“放牧+補(bǔ)飼”下植物群落生物量分配更為合理,能有效增強(qiáng)群落的穩(wěn)定性。因此,高寒草甸冬季牧場(chǎng)利用中,建議采用“放牧+補(bǔ)飼”方式,從而降低物種之間的競(jìng)爭(zhēng),使植物具有更好的生存空間,更有利于維持冬季牧場(chǎng)草地群落穩(wěn)定性,能對(duì)高寒草甸生態(tài)系統(tǒng)的健康發(fā)展產(chǎn)生積極影響。
猜你喜歡
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
中學(xué)生數(shù)理化(高中版.高考理化)(2021年6期)2021-07-28 06:21:04
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·少年版(2015年1期)2015-07-07 17:15:12
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
NBA特刊(2014年7期)2014-04-29 00:44:03
