硫鐵礦介導的自養反硝化研究進展
2023-10-14 08:01:08許中碩周盼盼王宇暉黃威宋新山
化工進展 2023年9期
關鍵詞:生物
許中碩,周盼盼,王宇暉,黃威,宋新山
(東華大學環境科學與工程學院,上海 201600)
水資源短缺、水環境破壞以及水生態破壞是制約我國高質量發展的突出瓶頸和生態文明建設的突出短板,污水資源化利用是破解上述問題的有效措施和多贏途徑。目前,我國城市各類污(廢)水年排放總量近700 億噸,是城市水環境的主要污染源,同時也是一種來源穩定、具有潛在利用價值的可再生資源[1]。但是城市排水中具有高濃度的氮磷等營養物質,危害生態系統和人類健康,制約了其資源化再生梯級分質利用。如何有效控制城市排水中氮磷污染物以保障再生水的安全合理利用,是城市污水資源化過程中亟需解決的問題。
城市排水中氮磷營養物質的深度控制目前已引起廣泛關注。隨著我國生態文明建設的推進,許多污水處理廠在常規水處理工藝后添加反硝化深度處理單元,以保證氮磷營養物質的高效去除,從而利于再生水回用于景觀生態補水、工業利用和市政雜用等,推進污水處理廠從“治污單功能”向“治污供水雙功能”轉變[2]。針對城鎮排水中氮磷濃度高而碳源含量少的特征,工程實踐表明高效脫氮除磷尚需外加碳源和除磷藥劑,從而使城鎮排水深度脫氮除磷的成本提高,成為城鎮排水深度處理領域可持續發展的制約性因素之一。相較于傳統異養反硝化,硫鐵礦自養反硝化是一種經濟、高效和綠色的脫氮處理技術,具有節省外加有機碳源、同步脫氮除磷、減少污泥產量和CO2排放量等優勢[3],是近年來深度處理領域研究的前沿和焦點。
1 硫鐵礦自養反硝化技術概述
1.1 硫鐵礦介導自養反硝化的發現
作為自然界普遍存在且儲量豐富的天然礦,硫鐵礦主要包括硫化亞鐵(FeS)、磁黃鐵礦(Fe1-xS)、膠黃鐵礦(Fe3S4)以及黃鐵礦(FeS2)等[4]。20 世紀,研究者已證實在缺氧和中性pH 條件下微生物可以驅動硫鐵礦介導的自養反硝化,是地下含水層、海洋和湖泊沉積物等自然水生環境中硝態氮還原的重要過程[5-7]。Golterman[8]于1991年發現在淡水系統中硫鐵礦物的氧化過程需要借助硝酸鹽的消耗。Garcia-Gil 等[5]于1992 年發現了海洋沉積物中FeS介導的脫氮作用。J?orgensen等[9]證明砂質含水層沉積物中黃鐵礦可以作為反硝化細菌的電子供體;Torrentó 等[10]也發現黃鐵礦的添加可以增強受硝酸鹽污染的地下水和沉積物中的反硝化作用,并提高反硝化細菌的豐度。Vaclavkova 等[11]也發現FeSx的添加可以提升缺氧含水層沉積物反應器中硝酸鹽的去除,而未添加硫鐵礦物的反應器中幾乎未觀察到硝酸鹽的減少。由上可知,自然生境中存在微生物驅動的硫鐵礦氧化與硝酸鹽還原的耦合現象。
盡管上述研究表明硫鐵礦可以作為硝態氮還原的有效電子供體,然而部分研究者認為這可能是由于硫鐵礦中殘存的還原態硫單質造成的,主要成分FeSx是否可以作為有效電子供體依然存在爭議[12]。Zhang 等[13]利 用 多 同 位 素(15N-NO-3、18N-NO-3、34S-SO2-4、18O-SO2-4以及34Spyrite)發現黃鐵礦氧化約占脫氮區硫酸鹽的70%,證實黃鐵礦是砂質含水層中脫氮的主要電子供體;其中,16S rRNA 基因測序顯示存在與硝酸鹽還原相結合的硫化物氧化細菌。Jakus 等[14]利用56Fe、57Fe 以及32S 等鐵硫同位素證實了黃鐵礦介導的自養反硝化中FeS2和殘存硫單質對硝氮去除率的貢獻率分別為26%和73%,直接證實了微生物可以驅動黃鐵礦的氧化。綜上所述,除了硫鐵礦中殘留的單質硫外,硫鐵礦(FeSx)本身亦可以作為反硝化過程的電子供體。
1.2 基于硫鐵礦的生物處理技術
基于自然生境中微生物驅動硫鐵礦氧化與硝態氮還原的耦合作用,研究者開始利用硫鐵礦(黃鐵礦、磁黃鐵礦等)同時作為電子供體和生物膜基質,開發相應的生物技術用以處理各類污水。目前基于硫鐵礦開發的生物處理技術主要包括生物濾池[15-17]、人工濕地[18-20]以及生物滯留池[21]等。Capua等[22]構建循環式黃鐵礦生物濾池,當水力停留時間大于8h 時,硝氮和磷的去除率分別超過了90%和70%。Liang等[23]發現相較于人工濕地空白組,添加了磁黃鐵礦的人工濕地系統處理效能大幅度提升,草甘膦、總磷以及總氮的平均去除率分別高達90.3%±6.1%、 88.2%±5.1% 以 及 60.4%±5.60%。Kong 等[24]開發了生物炭-黃鐵礦雙層生物滯留池,相較于傳統砂基或秸稈基生物滯留池,其對雨水徑流中碳氮磷組分的去除具有更好的穩定性和高效性。同時大量研究也發現,硫鐵礦介導微生物反硝化過程中磷污染物也得到了有效地去除[25-27],這是由于硫鐵礦的生物和化學氧化過程中無定形Fe(OH)3和γFeOOH 等產物具有高比表面積和正表面電荷,可吸附沉淀去除負電性的磷污染物[22,26]。由此可見,基于硫鐵礦的生物技術可通過“生物驅動反硝化-非生物化學除磷”雙重途徑協同作用實現氮磷的同步去除。
2 硫鐵礦自養反硝化技術的影響因素
硫鐵礦自養反硝化技術主要是由硫鐵礦與微生物的相互作用主導,可歸結于固相電子供體與反硝化細菌間的界面反應。該界面反應的影響因素主要包括硫鐵礦特性、添加量、運行pH以及溫度等。
2.1 硫鐵礦特性
目前廣泛研究的代表性硫鐵礦物主要包括硫化亞鐵、磁黃鐵礦以及黃鐵礦等。大量研究表明硫化亞鐵[5,28-29]、磁黃鐵礦[23,30-31]以及黃鐵礦[32-34]均可以作為電子供體以增強系統的反硝化性能;然而,他們結構上的差異會影響其作為電子供體的性能。其中,硫化亞鐵是鐵硫化物之間反應形成的第一種硫化鐵相,呈四方層狀結構,是黃鐵礦或磁黃鐵礦的前體物質[35]。磁黃鐵礦非化學計量組成顯示為Fe1-xS(0≤x≤0.125),其結構隨著不同的缺鐵量而呈現不同的形式[36]。黃鐵礦非化學計量組成顯示為FeS2,其呈NaCl 型結構,堅固的S—S 鍵和對稱的立方晶體結構使得其具有更高的穩定性[37]。研究發現黃鐵礦的反硝化能力可能弱于其前體物(如FeS),在相同培養實驗條件下FeS和FeS2組的硝態氮去除率分別為61.5%和54.1%[38]。Trouve等[39]也發現了脫氮硫桿菌對FeS的親和力大于FeS2,說明不同硫鐵礦在生物利用性上的差異可能是影響其作為電子供體時脫氮性能的主要因素。整體而言,典型硫鐵礦的結構特性差異會影響其作為電子供體的生物利用性。
研究發現硫鐵礦的晶面結構直接決定了其是否可以被反硝化細菌利用。Haaijer等[29]發現當高度結晶的黃鐵礦作為電子供體時,微生物無法驅動其生物氧化以實現硝態氮的還原。Yan 等[40]也證實了純結晶的黃鐵礦不能為反硝化菌提供有效電子供體,而合成的低結晶度的FeS2則可以實現以硝態氮為電子受體的生物氧化。最新研究也表明,相較于多晶面結構黃鐵礦,單晶面結構的黃鐵礦具有更高的表面能,穩定性更高,從而不利于反硝化細菌的利用,甚至無法作為電子供體[41]。李雅倩等[42]發現相較于六方磁鐵礦,低結晶度的單斜磁鐵礦表現出更優異的脫氮除磷活性。基于硫鐵礦晶面結構對其生物利用性的影響作用,研究者嘗試通過預處理手段改變硫鐵礦的物理結構特性以增強其生物利用性。Aquilina等[43]發現普通的礦物表面呈現低反應活性,而粉碎可通過誘發礦物表面缺失使其產生新的潛在的高反應活性表面,從而有利于含水層中基于黃鐵礦的自養反硝化。Yang等[27]利用無氧焙燒研制了納米晶粒組成的黃鐵礦,其具有蜂巢狀結構,可大大降低生物濾池出水氮和磷的濃度。綜上所述,硫鐵礦的物理化學特性是影響硫鐵礦自養反硝化是否發生及效率的重要因素。
2.2 硫鐵礦的添加量
硫鐵礦的添加量會影響系統的電子供給能力和鐵產物生成量,從而影響系統的脫氮除磷效能。一般而言,硫鐵礦的添加量與系統脫氮除磷能力存在正相關關系。Tong等[44]利用BBD模型和響應面法對黃鐵礦反硝化性能進行了研究和優化,當黃鐵礦劑量從50g/L 提高到125g/L 時,批次系統的反硝化速率顯著提高。Si等[26]發現在低溫條件下當人工濕地系統中黃鐵礦/火山巖的質量比從1∶10 提高到2∶1 時,NO-3-N 的去 除率從47.6% 增加到61.1%,PO3-4-P 去除率從25%升至74.7%。Chu 等[45]構建了黃鐵礦和石英不同體積比(0∶100%、25%∶75%、50%∶50%、75%∶25%、100%∶0)的垂直流人工濕地,發現隨著黃鐵礦含量的增加,TN和TP去除率呈現遞增趨勢。由此可知,隨著黃鐵礦添加量的增加,系統的脫氮和除磷能力均呈現明顯的上升趨勢。然而,微生物和氧氣驅動硫鐵礦的氧化作用均會導致SO2-4副產物的生成。Li等[46]研究表明在磁黃鐵礦自養生物濾池中,當水力停留時間為72h 時,出水SO2-4平均濃度高達(196.69±12.12)mg/L[進水SO2-4平均濃度僅為(34.6±1.3)mg/L]。基于污水排入城市下水道水質標準(SO42-<600mg/L)和飲用水標準(<250mg/L),該體系出水中硫酸根離子濃度的同步增加是后續研究中值得注意的問題。
2.3 pH
pH是影響硫鐵礦自養反硝化的重要因素之一,主要體現在其可以影響反硝化細菌活性以及硫鐵礦的溶解,從而影響其對該體系脫氮效能的調控[47]。一般而言,反硝化細菌的最適pH為6.5~8.0,過高或過低的pH都會抑制反硝化細菌的正常生命活動,從而對反硝化過程產生不利影響[39];然而,硫鐵礦介導的自養反硝化對pH 范圍的響應有所不同。Vaclavkova 等[11]發現在初始pH 為5.5(整個過程中4.5<pH<6)的條件下,砂質含水層中缺氧NO-3-N還原耦合FeSx氧化作用依然可以發生,并導致NO-3-N濃度逐漸降低。同時,研究發現從富含黃鐵礦的石灰巖含水層中分離的硝酸鹽還原依賴鐵氧化菌群,可以在pH為6.0~6.5條件下實現硝酸鹽的還原[48]。Torrentó 等[33]證實了酸性pH(<6.5)不會導致脫氮硫桿菌驅動的硫鐵礦自養反硝化速率的降低。此外,Li 等[16]發現在FeS 自養反硝化系統中,當初始pH 從7 下降到5 或者從7 升高至8 時,反硝化速率均呈現加快趨勢,但pH 大于8 時反硝化速率顯著下降。由此可知,硫鐵礦自養反硝化可以在較寬的pH 范圍內(5~8)發生,特別是在酸性pH條件下(5~6)。這可能是由于:①驅動硫鐵礦生物氧化的反硝化菌株可能具有低pH 的適應值[49];②較低的pH 可以促進硫鐵礦的溶解,從而提升其反硝化速率[16];③硫鐵礦可以作為一種有效的緩沖劑維持系統的pH穩定。
2.4 溫度
溫度是微生物新陳代謝的重要因素,它可以通過影響微生物增殖速率、氮還原酶活性等來控制反硝化過程[50]。一般而言,反硝化是一個溫度依賴的過程,其在低溫條件下易受到抑制。其中,自養反硝化菌的適宜溫度為20~40℃,而最佳生長溫度約為30℃;因此微生物驅動的硫鐵礦自養反硝化通常在室溫條件下發生。蒲驕陽[51]發現當運行溫度從30℃下降為20℃時,硫鐵礦反硝化系統的硝酸鹽去除率從99%下降至50%,且亞硝酸鹽含量明顯上升。Li 等[16]發現FeS 介導的反硝化作用可以在10~35℃范圍內發生,然而系統在5℃時幾乎不發生任何反硝化作用。Xu 等[52]發現黃鐵礦自養反硝化系統的NO-3-N 去除速率隨著溫度的降低而逐漸下降;其中,當溫度從20℃下降到10℃時,NO-3-N 去除率從49.2%下降到15.2%。由此可見,過低的溫度會抑制硫鐵礦介導的自養反硝化效能,如何在低溫條件下提升該系統的脫氮除磷效能還值得探究。
3 硫鐵礦自養反硝化技術的生物機制
3.1 硫鐵礦自養反硝化功能菌
研究表明硫氧化反硝化菌和亞鐵氧化反硝化菌是驅動硫鐵礦介導的自養反硝化過程的關鍵功能微生物[9,20,32]。目前典型的硫氧化反硝化菌主要包括脫氮硫桿菌(Thiobacillus denitrificans,T.denitrificans)、脫氮硫單胞菌(Sulfurimonas denitrificans,S.denitrificans)等[53];常見的鐵氧化反硝化菌主要包括微桿菌屬(Microbacteriumsp)[54]、氧化亞鐵硫桿菌(Thiobacillus ferrooxidans)[55]、氧化亞鐵假單胞菌(Pseudogulbenkiania ferrooxidans)[56]、腸桿菌(Enterobactersp.CC76)[57]、食酸菌(Acidovorax sp)[58]等。
目前硫鐵礦自養反硝化技術中優勢自養反硝化菌種多鑒定為硫氧化菌,其中以T.denitrificans為主。2006 年Haaijer 等[59]在富含硫鐵礦的天然生態系統中分離出β變形桿菌群Thiobacilli,并發現其可以在缺氧環境中氧化FeSx并耦合硝酸鹽還原。Torrentó 等[60]和Bosch 等[61]分別證實了T.denitrificans可以硝酸鹽為電子受體驅動納米級黃鐵礦或純黃鐵礦的生物氧化。周翔等[62]發現T.denitrificans可以FeS 為電子供體進行反硝化。朱良等[63]證實水熱法合成的FeS2粉末也可以作為硫自養厭氧反硝化污泥系統的電子供體,其中優勢菌屬為T.denitrificans。此外,S.denitrificans也是硫鐵礦介導的自養反硝化體系的優勢菌屬,其可在中性條件下利用無機硫化物和硫代硫酸鹽作為電子供體[64]。劉斌等[65]發現黃鐵礦生物濾池主要功能微生物除T.denitrificans(相對豐度27.6%)外,S.denitrificans(相對豐度11.8%)也占主要優勢。Li 等[46]發現磁黃鐵礦自養反硝化生物濾池處理缺碳廢水時,系統中最優勢菌種為S.denitrificans,其相對豐度高達68.05%。綜上所述,T.denitrificans和S.denitrificans等硫氧化反硝化菌屬在該體系中發揮重要作用。
除了硫氧化反硝化菌,亞鐵氧化反硝化菌被證實也可以利用硫鐵礦進行反硝化。研究者發現硝酸鹽還原依賴亞鐵氧化菌大多數不是專性自養生物,需要額外的有機碳作為能源進行持續生長[55-66],并多屬于嘉利翁氏菌科(Gallionellaceae)、嗜酸菌屬(Acidovoraxsp)、花生根瘤菌屬(Bradyrhizobiumsp)以及諾卡氏菌屬(Nocardioidessp)[67]等。同時,假單胞菌屬(Pseudogulbenkianiasp)、氧化亞鐵副球菌(Paracoccus ferrooxidans) 和弗氏檸檬酸桿菌(Citrobacter freundii)已被證明兼具亞鐵氧化和硝酸鹽還原的能力[68]。目前上述典型的硝酸鹽還原依賴亞鐵氧化細菌并未在硫鐵礦介導的反硝化體系中得到鑒定;同時,受限于微生物學的發展,尚有部分硝酸鹽還原依賴的亞鐵氧化菌處于富集混合培養而未分離或未鑒定完整種屬階段。然而,Senko等[69]分離出了硝酸鹽還原依賴亞鐵氧化細菌(命名為FW33AN),并證實其可以驅動FeS 氧化協同硝酸鹽還原。Jakus 等[48]從富含黃鐵礦的石灰含水層中富集培養了自養型硝酸鹽還原依賴的亞鐵氧化菌群,16S rRNA 基因測序表明其歸屬Gallionellaceae科;同時證實其可以同時驅動硝態氮的還原和Fe(Ⅱ)的氧化,NO-3-N/Fe(Ⅱ)的化學計量系數約為0.2。上述研究進一步說明了硝酸鹽還原依賴亞鐵氧化細菌可以驅動硫鐵礦的生物氧化,有助于推動對硫鐵礦自養反硝化體系微生物群落結構的全面認識。
3.2 硫鐵礦介導自養反硝化的原理
由上述研究可知,硫鐵礦介導的自養反硝化是通過“生物驅動反硝化-非生物化學除磷”雙重途徑協同作用實現氮磷的同步去除。以黃鐵礦為例,當其作為電子供體時,總反硝化反應方程式如式(1)所示。總體而言,該過程可能還涉及殘存硫單質自養反硝化[式(2)]、鐵化學除磷[式(3)和式(4)]等[4,70-71]。
盡管黃鐵礦自養反硝化是一個熱力學自發過程(ΔGo'=-2439kJ/mol),但是該反應只能在微生物催化作用下發生,且動力學比較緩慢[72]。目前研究雖然證實了微生物可以驅動硫鐵礦介導的反硝化作用,但其微觀機制仍不清楚,比如不溶性硫鐵礦與微生物的初始反應是如何發生的。目前部分研究認為硫鐵礦受到酸性溶解或氧化攻擊后,Fe和S之間的鍵被裂解,釋放出Fe2+和S2-2,之后Fe2+被迅速氧化成Fe3+并在中性pH 下生成鐵(氫)氧化物,S2-2被氧化生成各種無機硫中間體(如S2O2-3等)最終被氧化成SO2-4[73]。其中,對于黃鐵礦而言,其表面的S—S 鍵比Fe—S 鍵弱,S—S 更先斷裂,故而硫元素先被氧化生成SO2-3、S2O2-3等并最終轉化為SO2-4;鐵則主要以氧化物或者氫氧化物形式穩定存在[74]。部分研究認為微生物與礦物的相互作用可由胞外聚合物中酶(如鐵氧化酶MtoA和細胞色素c等)驅動[75];Pang等[32]研究發現在黃鐵礦自養反硝化過程中,參與電子穿梭體生物合成的menC/E/G等基因得到顯著富集,認為微生物衍生的電子穿梭體是黃鐵礦氧化的第一步。同時,Fe3+、鐵(氫)氧化物等中間產物通過沉淀吸附作用則可以同步去除PO3-4-P[76-77];此外,部分含鐵沉淀物有可能通過胞外聚合物中的金屬結合位點沉淀至生物膜。綜上所述,硫鐵礦介導的自養反硝化同步脫氮除磷潛在原理見圖1。
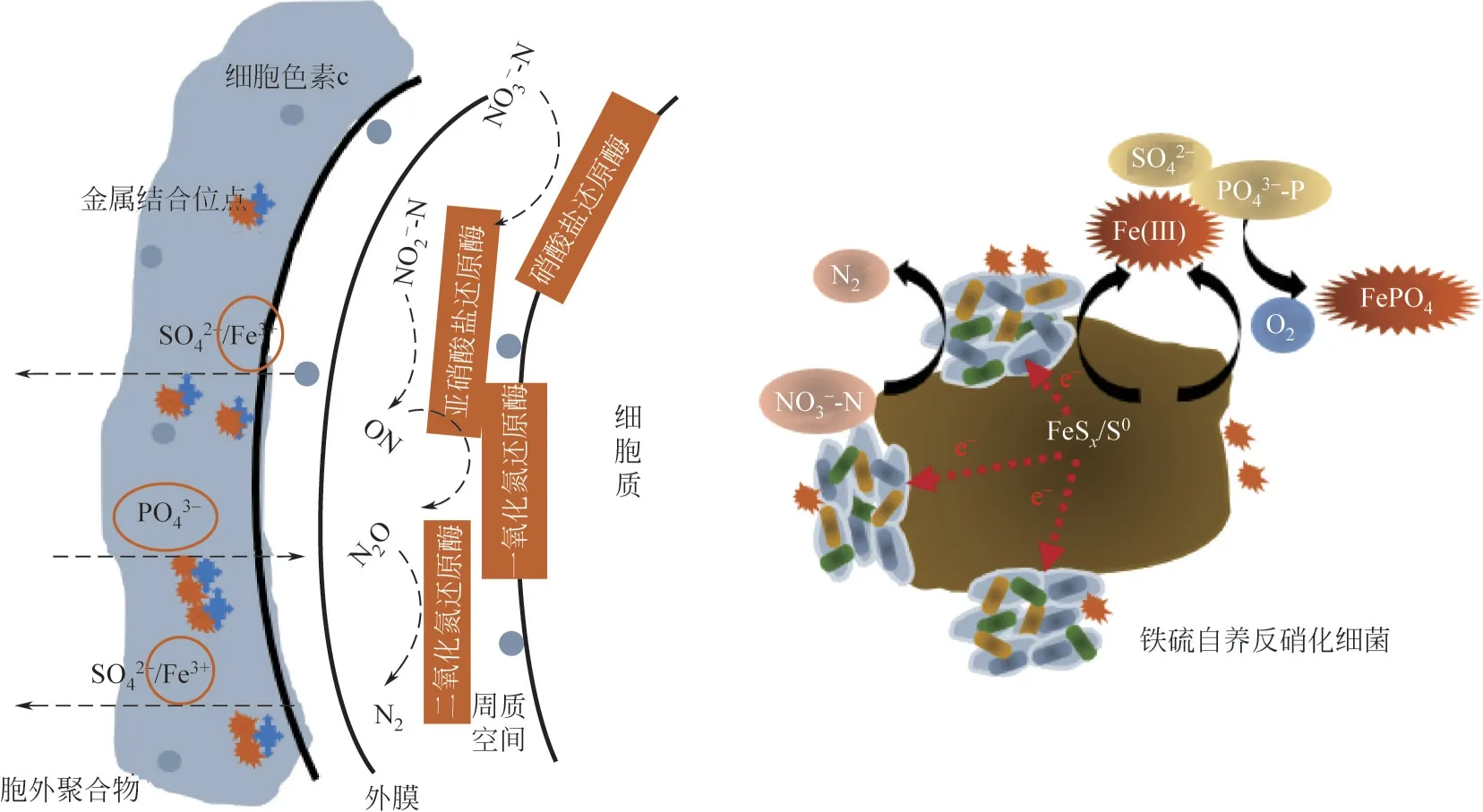
圖1 硫鐵礦自養反硝化同步脫氮除磷的潛在機制
4 技術的關鍵難點
4.1 硫鐵礦的生物利用性
由于微生物無法將硫鐵礦固體顆粒直接攝入到細胞中,硫鐵礦界面的電子直接轉移和溶解是被生物利用的潛在途徑[3]。在生物浸出過程中,根據硫鐵礦物的電子結構,存在酸性溶解和氧化溶解兩種溶解途徑。其中,價帶來源于鐵和硫化物原子軌道的硫鐵礦物(如FeS、Fe3S4、Fe1-xS、Fe7S8等)通常是酸溶性的,即容易受到質子攻擊而產生溶解生成Fe2+和H2S[78],這也與較低pH 下產生的高反硝化速率一致。然而,與單硫化物有所不同,黃鐵礦具有高度結晶性,其價帶完全來源于其金屬軌道,導致晶體對酸性或質子促溶作用呈現惰性,這與黃鐵礦的生物利用性低于FeS 存在關聯[79]。由此可知,相較于其他硫鐵礦,黃鐵礦的生物利用性則主要取決于直接電子轉移或氧化劑溶解。在黃鐵礦自養反硝化過程中,200mV 左右是黃鐵礦轉化為生物可利用有效形態的電化學閾值。從熱力學角度來看,中性條件下微生物中細胞色素a、c和d是可以氧化黃鐵礦的,即為微生物-礦物的直接電子轉移;但是這種直接作用是建立在微生物和礦物的接觸基礎上,意味著與礦物界面可用性有很大關系。同時,研究證實Fe3+作為強氧化劑,可與硫鐵礦發生化學反應加速其溶解;Moses等[80]發現相較于O2,Fe3+對黃鐵礦的溶解速率高出兩個數量級。Liu 等[81]也證實了有機配體EDTA 的添加通過形成Fe3+-EDTA 配合物,可以顯著加速黃鐵礦氧化并提升其反硝化性能。綜上所述,硫鐵礦的結晶度、反應pH、界面可用性、氧化劑(如Fe3+等)以及非還原有機配體都是影響硫鐵礦生物利用性的重要因素。
4.2 鐵沉積物的抑制作用
亞微米級高價態鐵產物在基質及微生物界面沉積聚集形成鐵鈍化膜,導致硫鐵電子轉移和氮磷物質傳輸受阻,進而可能誘發微生物活性惡化。一方面,通過化學氧化途徑生成的鐵氧化物和磷酸鐵等高價態鐵產物會沉積在黃鐵礦表面,限制甚至阻止反硝化菌和黃鐵礦的直接接觸,從而阻止硫鐵電子從礦物轉移到細胞外膜蛋白Cytc2 或鐵氧化酶MtoA[75]。另一方面,通過亞鐵胞外生物氧化(主要途徑)和亞鐵胞內化學氧化途徑形成的高價態鐵礦物微粒,會在胞外和周質空間等氧化位點聚集形成結殼現象[14,82-83],阻礙物質的傳輸和代謝,導致微生物活性降低而誘發系統惡化。化學納米成像顯示微生物分泌的黏液基質-胞外聚合物(EPS)在控制細胞高價態鐵礦物微粒的賦存形態起主導作用[84],且EPS中多糖為高價態鐵礦物微粒的潛在成核沉淀位點[85]。相關研究進一步證實EPS 中腐殖質、羧基、巰基、胺基以及膦基等金屬吸附位點會與高價態鐵礦物微粒緊密結合形成復合物[86]。因此,如何調控界面高價態鐵沉積物鈍化問題是實現硫鐵礦同步脫氮除磷長效運行面臨的難點。研究發現有機配體絡合法可以去除細胞鐵結殼物[84,87],進而恢復微生物新陳代謝,說明細胞鐵結殼現象屬于可逆的抑制作用。有機配體與黃鐵礦氧化產生的Fe3+相互作用形成較強的絡合物,不僅可以干擾(氫)氧化鐵的晶體結構、表面電荷和電勢等以阻止鐵(氫)氧化物的沉淀,而且降低了Fe(Ⅱ)/Fe(Ⅲ)循環的氧化還原電位。Liang等[88]發現天然有機物(絡合物)可以穩定地下水中Fe(Ⅱ)濃度并增加Fe(Ⅲ)濃度,說明有機配體通過增溶Fe3+進而阻止鐵(氫)氧化物形成上有一定的積極作用。其次,研究表明利用微超聲波可以促進厭氧氨氧化菌和硝酸鹽還原依賴的亞鐵氧化細菌等微生物細胞附著的金屬物質解吸,實現微生物與金屬物質的分離[86,89],從而解除金屬物質包裹的抑制作用而恢復微生物的活性。此外,具有鐵還原能力的菌種能夠以Fe(Ⅲ)礦物為電子受體將高價態鐵沉淀物溶解為Fe(Ⅱ),從而減緩或消除鐵鈍化膜的生成。近期研究也發現Fe(Ⅱ)/Fe(Ⅲ)循環可通過由NO-2-N 誘導的Fe(Ⅱ)氧化和NH+4-N 誘導的Fe(Ⅲ)還原,推動總氮的去除[90]。由此推斷,如果向該系統提供外部電子受體(如氨),Fe(Ⅱ)/Fe(Ⅲ)循環將得到極大的促進,這不僅可以減輕鐵沉積物鈍化問題,還可以提高總氮的去除率。綜上所述,硫鐵礦自養反硝化中鐵沉積物鈍化問題仍需要進一步深度探究。
5 結語與展望
硫鐵礦介導自養反硝化具有節省外加有機碳源、同步脫氮除磷、且減少污泥產量和CO2排放量等優勢,是一種經濟、高效和綠色的脫氮技術。目前基于硫鐵礦的各類生物處理技術已證實可以高效處理城鎮廢水、地下水以及雨水等,在低碳廢水處理領域具有優勢的應用前景。然而,目前研究多聚焦于該技術的脫氮除磷性能、運行參數以及微生物群落結構等;而對硫鐵礦介導自養反硝化的微觀生物機制研究較少,硫鐵礦生物氧化與反硝化耦合過程中的電子傳遞機制仍不清楚。同時,結合該技術的發展難點,該技術未來的研究方向與發展重點應包括:①繼續深入探究硫鐵礦介導反硝化的微生物機制,尤其是微生物胞外聚合物中細胞色素c等電子穿梭體;②如何增強硫鐵礦的生物可利用性;③如何解除硫鐵礦基質和生物膜界面鐵沉積物的抑制作用。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
