水熱預處理對餐廚垃圾厭氧發(fā)酵產戊酸的影響
2023-10-14 08:01:54王雪婷顧霞徐先寶趙磊薛罡李響
化工進展 2023年9期
王雪婷,顧霞,徐先寶,趙磊,薛罡,李響
(1 東華大學環(huán)境科學與工程學院,上海 201620;2 維爾利環(huán)保科技集團股份有限公司,江蘇 常州 213125)
戊酸是含有五個碳原子的飽和脂肪酸[1],已廣泛應用于化工、制藥、食品、飲料和紡織等行業(yè)[2],可用于生產防腐劑、稀釋劑、藥物、增塑劑、香料、纖維、添加劑以及可生物降解塑料聚羥基烷酸酯[3-4]。目前,戊酸生產主要依賴于石油化工以及化學合成[5],然而化工合成能耗大、成本高且會產生環(huán)境污染,因此,拓展綠色的戊酸生產方式具有廣闊的前景和意義。
近年來,利用碳鏈延長方法將有機廢棄物轉化為有機酸[6]的方式越來越受到研究者的關注。碳鏈延長技術是特定的碳鏈延長微生物利用乙醇、乳酸等能源物質作為電子供體,以乙酸、丙酸等短鏈羧酸作為電子受體[7],在生物酶的催化作用下,通過反向β 氧化循環(huán)反應將短鏈羧酸進行碳鏈延長[8]。早期利用碳鏈延長方法產戊酸的研究多集中于純底物和純菌,Grootscholten 等[9]和Ganigué 等[10]在純底物體系投加外源乙醇并接種克氏梭菌(Clostridium kluyveri)生產戊酸,最終戊酸的產量分別可達10.61g COD/L 和10.81g COD/L。但是純菌體系操作煩瑣,耐沖擊負荷能力弱,且運行成本高,不利于工業(yè)化的大規(guī)模生產[11]。因此研究者嘗試利用混菌體系生產戊酸,餐廚垃圾因其有機質豐富成為研究中常用的底物。在前期的研究中選取了兩種常用的菌源(剩余污泥和酒曲),結果表明在接種剩余污泥的發(fā)酵體系中戊酸產量為5.01g COD/L,而接種酒曲的發(fā)酵體系中戊酸產量僅為2.83g COD/L[12],混菌體系戊酸的產量遠低于純菌體系。此外,分析得知,酒曲體系中產生了高濃度的乙醇,而研究證實醇酸比高時易產生高濃度的己酸和庚酸,醇酸比低時易產生大量戊酸[13],因此調控乙醇的產量有利于提高發(fā)酵體系中戊酸的產量。
Taherzadeh 等[14]和Taue 等[15]的研究發(fā)現呋喃、糠醛和羥甲基糠醛會降低發(fā)酵體系中乙醇的生成,可使乙醇的產量降低80%,而餐廚垃圾水熱預處理過程中會發(fā)生美拉德反應,產生類黑精類物質、呋喃、糠醛和胺類物質等[16]。因此利用水熱法預處理餐廚垃圾抑制乙醇的產量,從而提高戊酸的產量是一種可行的方法,然而尚缺乏水熱預處理對餐廚垃圾厭氧發(fā)酵產戊酸的效果驗證。
本文基于前期的研究基礎[12],選取了戊酸產量高的剩余污泥體系和戊酸產量低的酒曲體系作為菌源,以批次發(fā)酵的方式,研究了在不同水熱預處理溫度下,餐廚垃圾厭氧發(fā)酵產戊酸的效能,探究了產酸代謝機制,同時研究了微生物群落組成變化,為餐廚垃圾高值資源化提供技術支撐。
1 實驗部分
1.1 實驗材料
餐廚垃圾,東華大學食堂,以蔬菜、米飯和肉為主,揮發(fā)性固體(VS)值為0.38g/g,經手工篩選和攪拌機攪碎后放置在4℃的冰箱內保存?zhèn)溆茫皇S辔勰啵虾J心澄鬯幚韽S,VS 值為11.10g/g,室溫下保存?zhèn)溆茫痪魄虾J心尘茝S,VS值為0.78g/g,經攪拌機混勻放置在4℃的冰箱內保存?zhèn)溆谩?/p>
1.2 實驗裝置與方法
發(fā)酵罐運行之前,將餐廚垃圾分別在80℃、100℃、120℃、140℃、160℃、180℃條件下進行水熱預處理,隨后將不同水熱預處理溫度下得到的發(fā)酵底物分成兩組:其中一組接種酒曲(DY-B、DY-80、DY-100、DY-120、DY-140、DY-160、DY-180);另一組接種剩余污泥(WAS-B、WAS-80、WAS-100、WAS-120、WAS-140、WAS-160、WAS-180)。發(fā)酵罐有效容積為200mL,底物濃度為(42.0±0.5)g VS/L(餐廚垃圾∶菌源=6∶1),投加20g/L 的CaCO3緩沖pH。將厭氧發(fā)酵罐置于磁力攪拌器上,放置于37℃恒溫箱內。
發(fā)酵周期為20天,每隔2天進行一次取樣,將5mL 發(fā)酵液置于離心管中經8000r/min 離心10min。取上層清液經0.45μm 濾膜過濾后,用于分析發(fā)酵體系中的戊酸、乙醇、乙酸、丙酸、丁酸、己酸和庚酸的變化情況;下層固體物質標記后置于-20℃的冰箱中保存,用于體系中微生物的分析。
1.3 組分分析方法
揮發(fā)性固體量采用馬弗爐重量法測定;戊酸、乙醇、乙酸、丙酸、丁酸、己酸和庚酸的測定采用氣相色譜儀GCSmart(GC-2018)測定,檢測器類型為FID 檢測器,檢測器溫度設為250℃,進樣口溫度設為220℃,載氣為N2,載氣流速設為50mL/min,測定時間為15min,利用外標法進行定量;餐廚垃圾水熱預處理后溶解性有機物的測定采用熒光光度計(Hitachi F-7000)進行掃描測定,利用Matlab軟件對掃描結果進行分析。
1.4 微生物測序方法
選擇發(fā)酵12天時的DY-B、DY-100、DY-140、DY-180 和 WAS-B、 WAS-100、 WAS-140、WAS-180的8個樣品進行微生物群落分析。微生物群落樣品選擇高通量測序技術進行分析,測樣公司為上海派森諾生物科技有限公司,PCR 擴增引物序列為338F(5'-ACTCCTACGGGAGGCAGCA-3')和806R (5'-GGACTACHVGGGTWTCTAA T-3'),擴增區(qū)域為V3~V4區(qū),測序平臺為Illumina Miseq。
2 結果與討論
2.1 水熱預處理對餐廚垃圾發(fā)酵產戊酸的影響
由圖1(a)結果可知,接種酒曲體系下,空白組戊酸最大產量為3.47g COD/L,水熱預處理后,戊酸最大濃度分別提升至13.03g COD/L (80℃)、12.05g COD/L (100℃)、12.68g COD/L (120℃)、14.81g COD/L(140℃)、12.67g COD/L(160℃)和16.19g COD/L(180℃)。結果表明,水熱預處理顯著提高了酒曲體系中戊酸的產量,其中DY-180組戊酸產量與DY-B 相比提高了3.66 倍。由圖1(b)結果可知,接種剩余污泥體系下,空白組戊酸最大產量為10.64g COD/L,高于接種酒曲的空白組,與郭志超等[12]的研究結果一致。水熱預處理后,戊酸最大濃度提升至12.57g COD/L(80℃)、12.73g COD/L(100℃)、12.63g COD/L (120℃)、13.11g COD/L(140℃)、15.75g COD/L(160℃)和18.55g COD/L(180℃)。結果表明,水熱預處理同樣提高了剩余污泥體系中戊酸產量,WAS-180 組戊酸產量與WAS-B 組相比提高了0.74 倍。研究表明,以粗甘油為底物時戊酸的產量僅為6.9g COD/L[17];以剩余污泥為底物時戊酸的產量僅為2.82g COD/L[2]。本文以成分復雜的餐廚垃圾作為發(fā)酵底物,并利用水熱預處理的方法顯著提高了體系中戊酸的產量,最高產量可達18.55g COD/L。
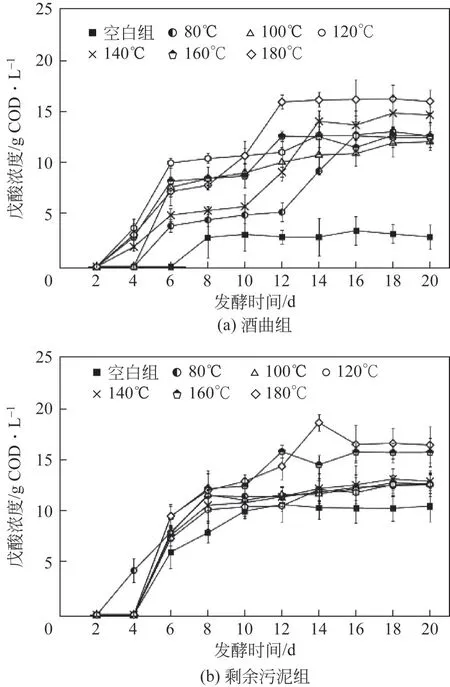
圖1 不同水熱預處理發(fā)酵體系下戊酸的產量
2.2 水熱預處理對餐廚垃圾發(fā)酵產有機酸及乙醇的影響
2.2.1 水熱預處理對戊酸代謝過程的影響
戊酸的產生和消耗主要是通過反向β 氧化途徑,微生物利用體系中乙醇作為電子供體,丙酸作為電子受體通過一次反向β循環(huán)產生戊酸,同時戊酸也可作為電子受體與乙醇通過反向β循環(huán)產生庚酸[18],因此關注發(fā)酵體系內丙酸和庚酸的變化對戊酸的產生和積累尤其重要。如圖2所示,在酒曲體系中,空白組丙酸的最大產量是2.16g COD/L,水熱預處理組丙酸的最大產量可達到10.58g COD/L(180℃);在剩余污泥體系中,空白組丙酸的最大產量是12.10g COD/L,水熱預處理組丙酸的最大產量可達到14.63g COD/L(120℃)。結果表明,水熱預處理后接種酒曲及污泥體系中的丙酸產量均有顯著提高。研究發(fā)現,腐殖酸可以提高水解酶的活性,增強電子轉移效率,促進發(fā)酵體系產短鏈羧酸[19]。如圖3 所示,隨著水熱預處理溫度的提高,餐廚垃圾中腐殖酸的比例逐漸增加,體系中丙酸濃度的提高可能與腐殖酸濃度的增加有關。同時研究發(fā)現,水熱預處理會提高體系中乳酸的產量,部分乳酸會通過丙烯酸途徑轉化為丙酸,實現了體系中丙酸的積累[20]。丙酸與乙醇氧化形成的乙酰輔酶A縮合形成戊酸,因此經水熱預處理后的酒曲體系和剩余污泥體系中戊酸的產量會增加,該過程的反應方程式見式(1)、式(2)、式(4)、式(5)[21]。在戊酸的消耗方面,隨著發(fā)酵時間的延長,在電子供體充足的情況下,戊酸會作為電子受體通過一次反向β循環(huán)生成庚酸,該過程的反應方程式見式(6)[21]。酒曲和剩余污泥體系中,空白組中庚酸的產量可分別達到1.84g COD/L 和2.33g COD/L,經水熱預處理后體系中庚酸的產量均下降,其中最低產量分別為0g COD/L(DY-120)和0.71g COD/L(WAS-160),水熱預處理減少了戊酸的消耗,實現了戊酸的積累。
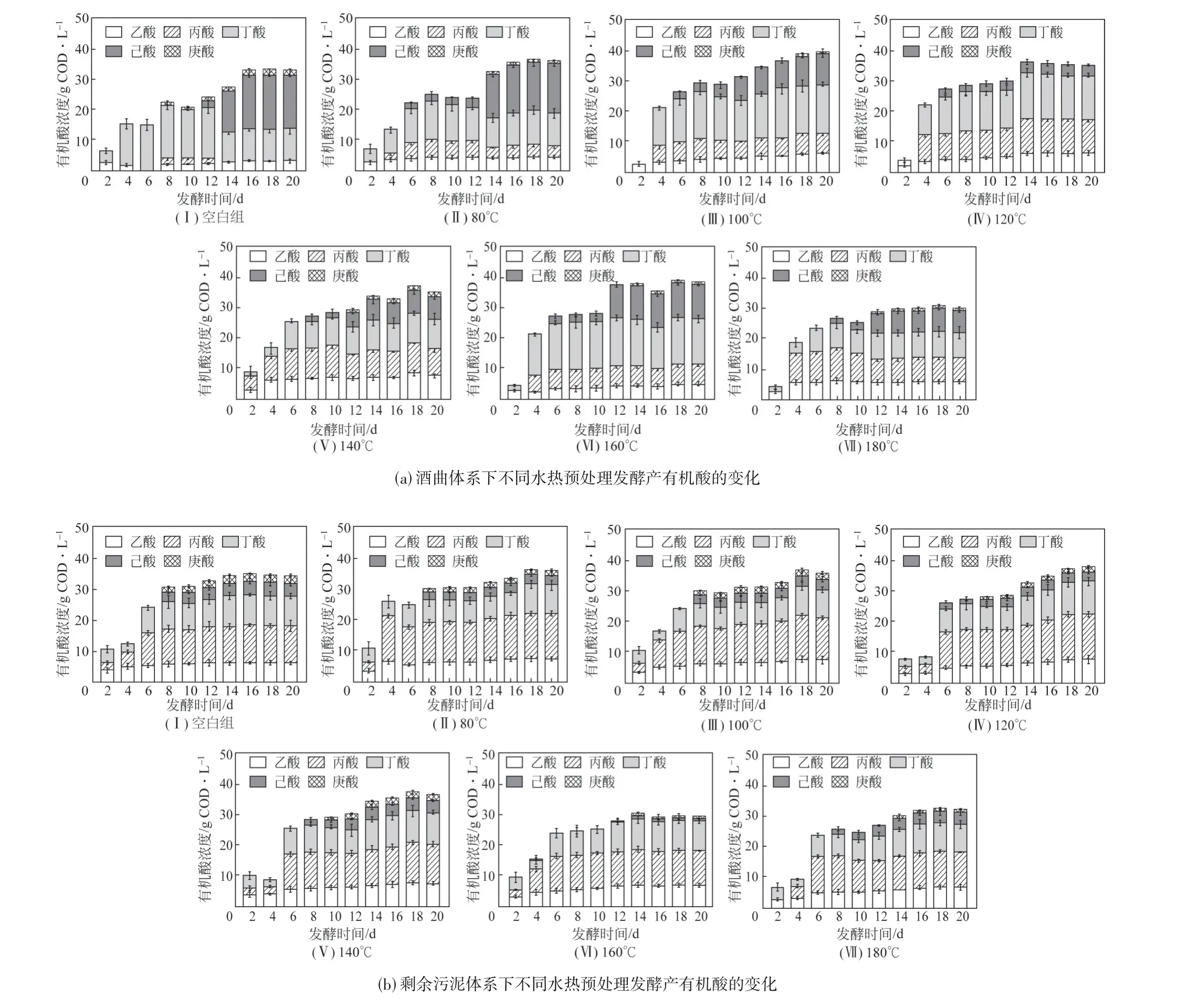
圖2 不同水熱預處理發(fā)酵體系下有機酸的變化
水熱預處理對乙酸、丁酸和己酸的產量如圖2所示,在酒曲和剩余污泥體系中,水熱預處理后乙酸產量與空白組相比均有提高,而水熱預處理后丁酸和己酸的產量與空白組相比顯著下降。研究發(fā)現水熱預處理會產生類黑精類物質(類腐殖酸區(qū)),而類黑精類物質會抑制發(fā)酵過程中乙酸向丁酸的轉化[22],因此水熱預處理組乙酸產量會有明顯的積累,該過程的反應方程式見式(3)和式(7)[21]。體系中己酸產量的降低可能與電子受體丁酸不足,微生物無法利用乙醇進行碳鏈延長反應有關,該過程的反應方程式見式(8)[21]。丁酸和己酸濃度的降低減少了碳鏈延長過程中乙醇的消耗,為生產戊酸積累了充足的電子供體。
2.2.2 水熱預處理對產乙醇的影響
乙醇是富含能量的還原性物質,為碳鏈延長提供能量,可氧化產生乙酰輔酶A[23]進而發(fā)生縮合反應促進碳鏈延長反應,是產戊酸的重要電子供體。低濃度的乙醇會導致碳鏈延長不充分,而乙醇濃度過高會對微生物有毒害作用[24]。如圖4所示,在酒曲和剩余污泥體系中,空白組乙醇濃度最高可分別達到14.90g COD/L 和3.96g COD/L,水熱預處理組乙醇濃度與空白組相比有明顯的下降,這與空白組產生高濃度的中長鏈羧酸(己酸和庚酸)而水熱預處理組產生高濃度的戊酸現象一致。
研究表明,醇酸比對發(fā)酵體系中有機酸的組成和產量有重要影響[25],醇酸比高時易產生高濃度的己酸和庚酸,醇酸比低時易產生大量戊酸[13]。乙醇和丙酸可以通過一次反向β循環(huán)產生戊酸,本研究中乙醇/丙酸的值如表1 所示,空白組醇酸比分別為6.48(酒曲體系)和1.55(剩余污泥體系),水熱預處理后醇酸比與空白組相比有明顯的下降。在酒曲體系中,DY-B(6.48)和DY-80(4.66)醇酸比高,這與體系中高濃度己酸和庚酸現象一致,而DY-100、DY-140、DY-160 和DY-180 的醇酸比均處于較低水平(0.14~1.30),因此產生了高濃度的戊酸。在剩余污泥體系中,所有反應罐中醇酸比均處于較低水平(0.40~1.55),故剩余污泥組戊酸的產量高。上述現象與苑榮雪[26]研究的醇酸比對厭氧發(fā)酵中產有機酸的影響一致,醇酸比為3時主要以產己酸和庚酸為主,而醇酸比為1 和0.5 時,戊酸的產量與高醇酸比組相比有顯著升高。另外,如圖5所示,在酒曲和剩余污泥體系中,戊酸產量較高的兩組(DY-180和WAS-180)發(fā)酵前期(2~8 天),乙醇、乙酸、丙酸和丁酸的濃度均呈現上升趨勢,醇酸比較高,電子受體乙酸易與電子供體乙醇反應產生丁酸,消耗了電子供體乙醇,此階段戊酸的產量緩慢增長。在發(fā)酵第8~12 天,隨著戊酸產量的快速提高,乙醇和丙酸分別作為電子供體和電子受體被消耗,濃度呈快速下降趨勢。當戊酸的產量達到峰值時,此時乙醇濃度突然降為0。在發(fā)酵后期第16~20天時,戊酸、丁酸、己酸和庚酸的產量保持不變,這可能與體系中缺乏電子供體乙醇有關。水熱預處理可以降低發(fā)酵體系中乙醇的濃度,調控醇酸比,促進餐廚垃圾厭氧發(fā)酵體系中戊酸的產生和積累。
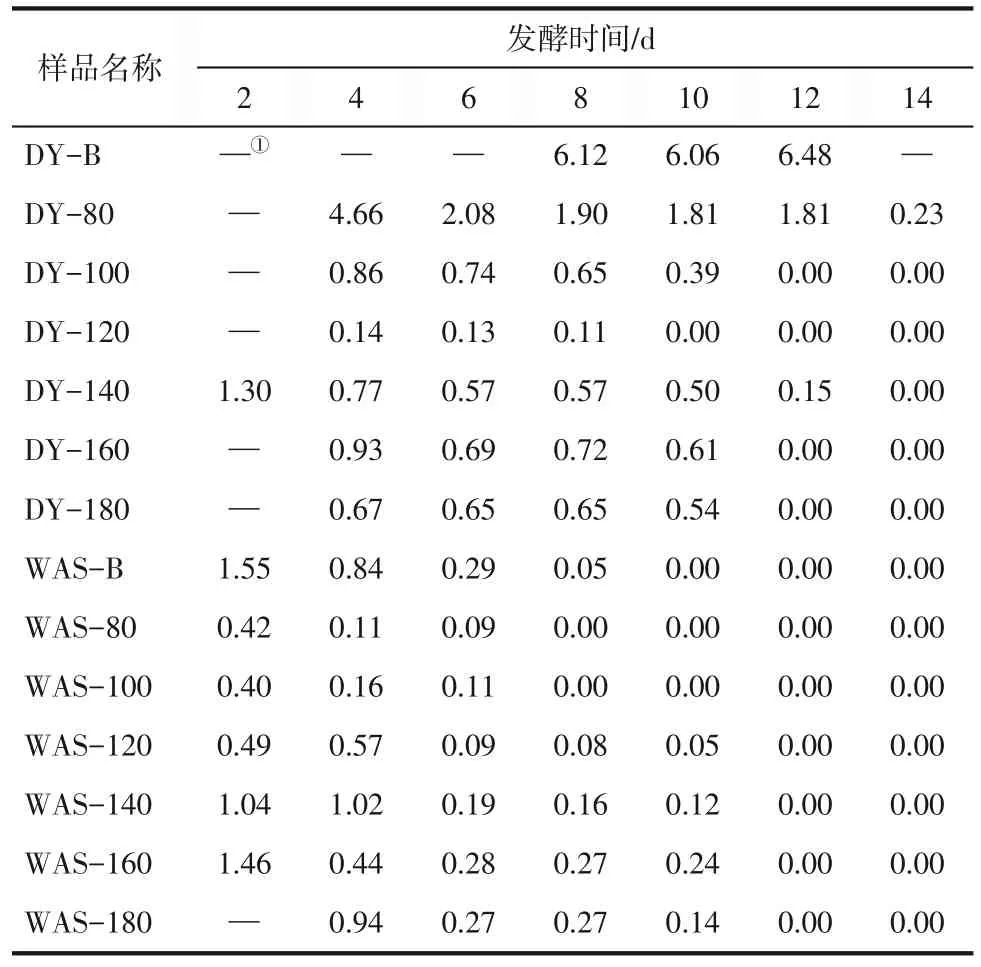
表1 不同發(fā)酵時間下各樣品的乙醇/丙酸值
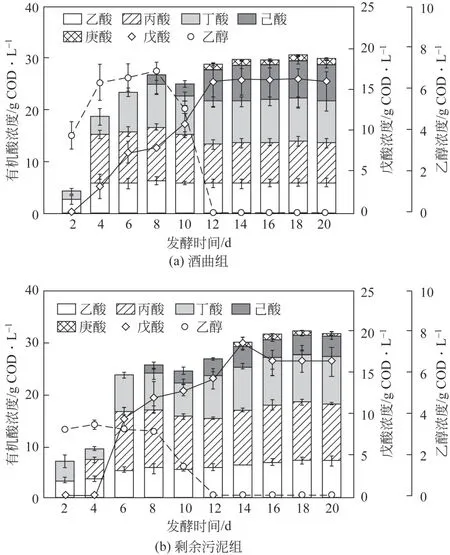
圖5 180℃時不同菌源發(fā)酵體系下物質的產量
2.3 微生物群落結構分析
戊酸的產生與電子供體乙醇密切相關,在發(fā)酵的第12 天,接種酒曲和剩余污泥的體系中乙醇的濃度大部分均下降至0,因此選取了發(fā)酵第12天時的樣品進行Alpha 多樣性分析和微生物群落結構分析。
2.3.1 Alpha多樣性分析
各發(fā)酵罐的Chao1、Shannon 和Simpson 指數如表2所示,在酒曲體系中,原始酒曲的微生物群落單一[12],水熱預處理后細菌群落Chao1、Shannon和Simpson 指數與空白組相比有明顯的提高,說明水熱預處理后微生物群落豐富度和多樣性更高,群落結構趨于復雜,豐富了參與溶出、水解和產戊酸的功能菌群,水熱預處理組戊酸產量的增加可能與產戊酸功能菌群的豐度提高有關。在剩余污泥體系中,原始剩余污泥微生物種群豐富度高[27],群落結構復雜,水熱預處理降低了細菌群落Chao1、Shannon 和Simpson 指數,這說明水熱預處理降低了微生物群落豐富度和多樣性,降低了競爭代謝微生物菌群。
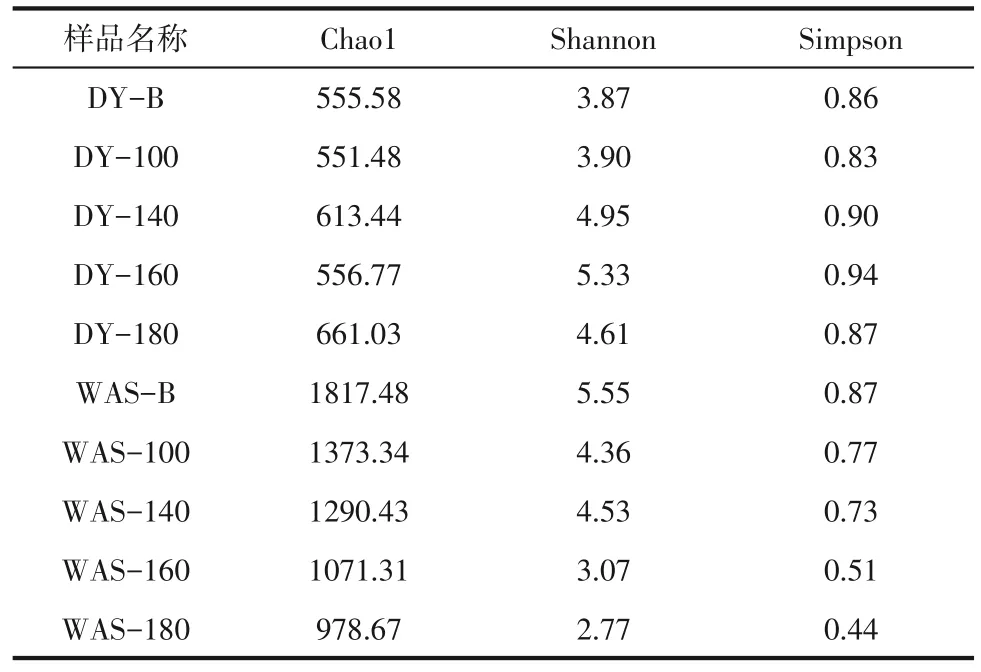
表2 不同樣品的Chaol、Shannon和Simpson指數
2.3.2 微生物群落結構分析
微生物門水平群落結構組成如圖6所示,各發(fā)酵罐主要由厚壁菌門(Firmicutes)、放線菌門(Actinobacteria)、變形菌門(Proteobacteria)和擬桿菌門(Bacteroidetes)組成,其與厭氧發(fā)酵過程密切相關,能產生大量的酶,促進底物的水解酸化產乳酸和揮發(fā)性脂肪酸[28-29]。其中優(yōu)勢菌門Firmicutes在所有發(fā)酵罐中的相對豐度均超過40%,隨著水熱溫度的升高,Firmicutes的相對豐度逐漸提高。在酒曲體系中,Firmicutes的相對豐度從44.81%(DY-B)提高到66.03%(DY-180),剩余污泥體系中Firmicutes的相對豐度從40.40%(WAS-B)提高到88.41%(WAS-180),研究表明Firmicutes對極端環(huán)境的耐受性較強,能適應不斷酸化的環(huán)境,并且目前已經被證實有碳鏈延長功能的細菌大部分都屬于Firmicutes[30],因此水熱預處理組戊酸產量的提高可能與這類微生物有關。
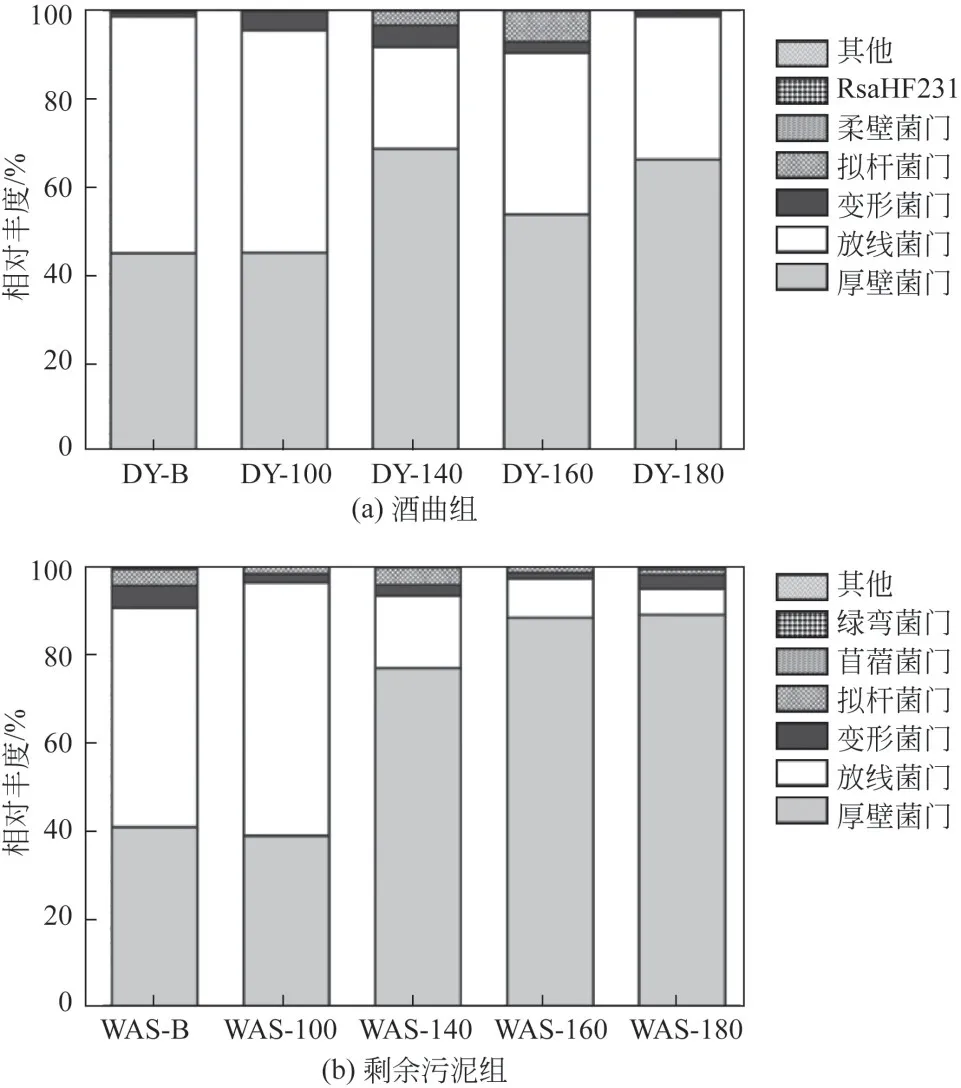
圖6 不同水熱預處理發(fā)酵體系下門水平群落結構組成
微生物屬水平群落結構組成如圖7所示,在酒曲體系中,DY-B組主要由雙歧桿菌屬(Bifidobacterium)(27.56%)和放線菌屬(Actinomyces)(25.80%)組成,這兩類微生物均與復雜有機物質的水解及乙酸和丁酸的生成有關[31],因此DY-B 組丁酸產量高而戊酸產量低可能與這兩類菌屬有關。水熱預處理后, 瘤 胃 球 菌 屬(UBA1819)、 腸 球 菌 屬(Enterococcus)、巨球菌屬(Megasphaera)和小類桿菌屬(Dialister)被選擇性富集。其中,UBA1819和Enterococcus均與產乙酸和丙酸有關[32],這可能是造成水熱預處理組乙酸和丙酸積累的原因。研究發(fā)現Megasphaera和Dialister均是產戊酸功能菌群[33],在DY-B 中,無Megasphaera和Dialister富集,而DY-100、DY-140、DY-160 和DY-180 中Megasphaera的相對豐度分別提高到2.13%、16.02%、4.52%和13.77%;Dialister的相對豐度為0%、2.86%、6.65%和2.26%。結果表明,水熱預處理實現了產戊酸相關功能菌群的富集,這可能是水熱預處理組戊酸產量高的原因。
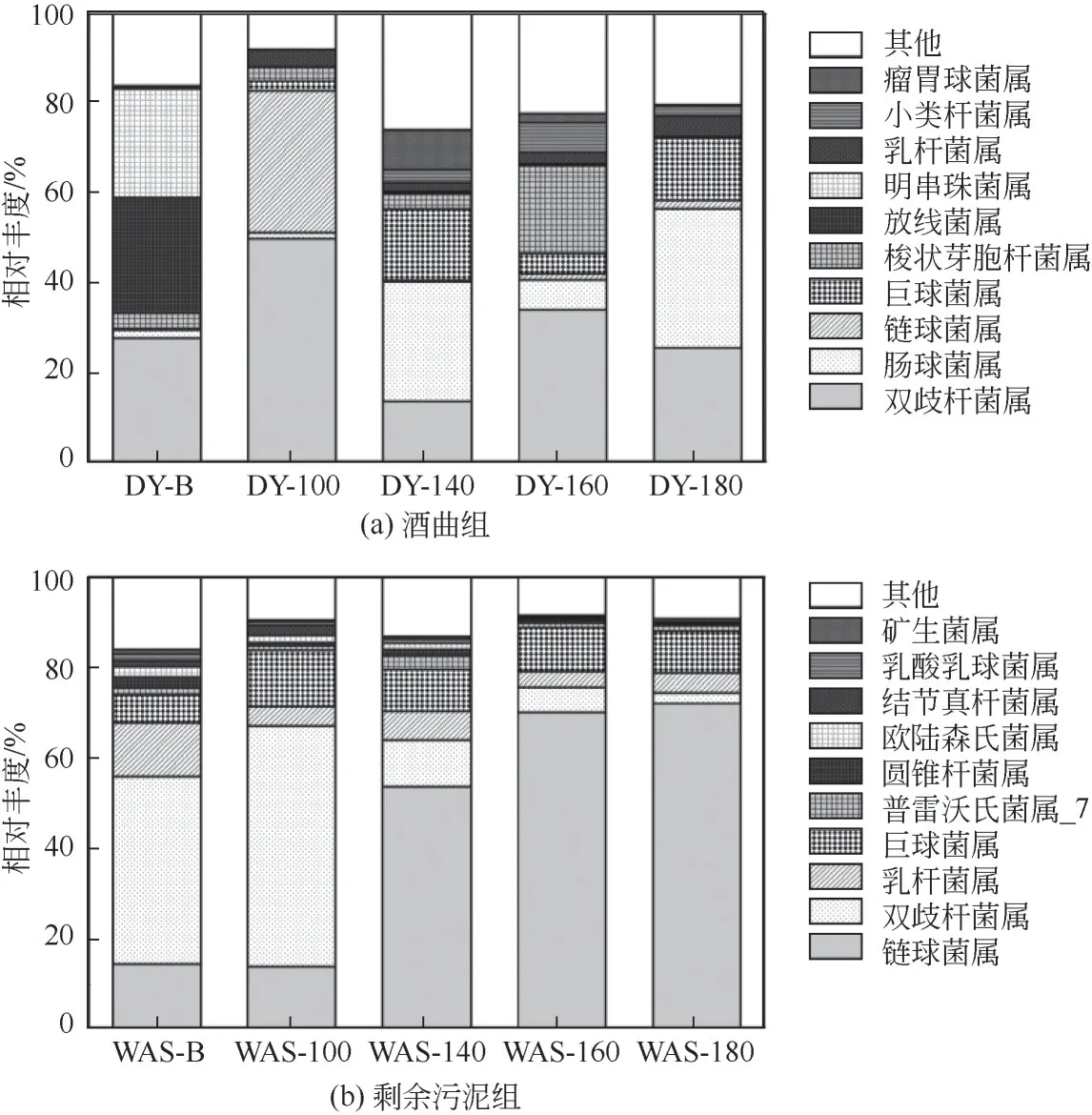
圖7 不同水熱預處理發(fā)酵體系下屬水平群落結構組成
在剩余污泥體系中,發(fā)酵罐中均含有可以將底物水解、 酸化以及產乳酸的鏈球菌屬(Streptococcus)[34]。在WAS-B、WAS-100、WAS-140、WAS-160 和WAS-180 中的相對豐度分別是14.09%、13.60%、53.52%、69.92%和72.92%,隨著水熱溫度的提高,Streptococcus的相對豐度逐漸提高。除此之外,所有的反應器中均選擇性富集了巨球菌屬(Megasphaera) 和普雷沃氏菌屬_7(Prevotella_7),Megasphaera在WAS-B、 WAS-100、WAS-140、WAS-160 和WAS-180 中的相對豐度分別是6.21%、12.38%、9.22%、9.77% 和9.35%,水熱預處理提高了Megasphaera的相對豐度;Prevotella_7 在WAS-B、 WAS-100、 WAS-140、WAS-160 和WAS-180 中的相對豐度分別是1.68%、1.04%、2.99%、1.04%和1.19%。已有研究證明Megasphaera和Prevotella_7 可以產生高濃度丙酸和戊酸[35],水熱預處理組產戊酸功能菌群的總相對豐度高于空白組,這可能是水熱預處理組戊酸產量升高的原因之一。另外,WAS-B 中產戊酸功能菌群的相對豐度是7.89%,而DY-B 無產戊酸功能菌群的富集,因此WAS-B 中戊酸的產量高于DY-B。
3 結論
(1)水熱預處理可以顯著提高戊酸的產量,在水熱預處理溫度為180℃時,戊酸的產量達到最大值。在接種酒曲體系中,戊酸的產量從3.47g COD/L 提高至16.19g COD/L,與空白組相比提高了3.66倍;在接種剩余污泥體系中,戊酸的產量從10.64g COD/L 提高至18.55g COD/L,與空白組相比提高了0.74倍。
(2)水熱預處理提高了發(fā)酵體系中丙酸的產量,降低了庚酸和己酸的產量,調控了發(fā)酵體系的醇酸比,促進乙醇和丙酸轉化為戊酸。
(3)水熱預處理促進了產戊酸功能菌群的高效富集。在接種酒曲體系中,空白組無與產戊酸相關功菌群的富集,水熱預處理溫度為180℃時與產戊酸相關功菌群Megasphaera和Dialister的相對豐度分別增加至13.77%和2.26%;在接種剩余污泥體系中,空白組Megasphaera的相對豐度為6.21%,水熱預處理溫度為180℃時Megasphaera的相對豐度最高加至9.35%。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(yè)(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
今日農業(yè)(2020年20期)2020-11-26 06:09:10
中國外匯(2019年17期)2019-11-16 09:31:14
中國果業(yè)信息(2019年10期)2019-11-13 01:21:34
聚氯乙烯(2018年9期)2018-02-18 01:11:34
現代企業(yè)(2015年1期)2015-02-28 18:43:18
新高考·高一物理(2014年1期)2014-09-18 01:26:07
