日照沿海養殖三疣梭子蟹血卵渦鞭蟲流行病發生過程研究*
2023-10-17 07:12:18劉煒鑫陳曉玲王君霞張緒濤黃李才文
海洋與湖沼 2023年5期
劉煒鑫 陳曉玲 李 蒙 王君霞 張緒濤黃 騫 李才文,4,5①
(1.中國科學院海洋研究所海洋生態與環境科學重點實驗室 山東青島 266071; 2.日照市海洋與漁業研究院 山東日照276826; 3.嶗山實驗室海洋生態與環境科學功能實驗室 山東青島 266237; 4.中國科學院大學 北京 100049;5.中國科學院海洋大科學研究中心 山東青島 266071)
三疣梭子蟹(Portunus trituberculatus)隸屬于甲殼綱(Crustacea)、十足目(Decapoda)、梭子蟹科(Portunidae)、梭子蟹屬(Portunus), 是我國沿海的重要養殖經濟蟹類之一, 養殖區域主要涵蓋江蘇、山東半島、浙江、福建、廣東等沿海地區, 其2021年產量超過10萬噸(農業農村部漁業漁政管理局等, 2022)。日照是山東省沿海地市三疣梭子蟹規模化養殖的重要區域, 已有20 多年的養殖歷史。2007 年以來, 日照地區養殖三疣梭子蟹頻繁發病發生大規模死亡事件; 病蟹個體主要表現為行動遲緩、溜邊, 停止攝食, 逐漸消瘦, 體重下降; 病蟹肌肉呈現白濁、乳黃色病變, 打開病蟹殼后通常可見大量黃白色、乳白色液體, 伴隨有空腸、空胃等情況, 據此當地養殖戶將該病害稱為“牙膏病”或“牛奶病”。夏季暴發期間, 養殖池塘中梭子蟹死亡率達到60%~80%, 給當地養殖戶造成了嚴重經濟損失。
血卵渦鞭蟲(Hematodiniumspp.)是一類主要感染海水甲殼類的致病性寄生甲藻(parasitic dinoflagellates),在世界范圍內感染40 多種蟹、蝦類, 其中包括美國蘭蟹、挪威龍蝦、帝王蟹、三疣梭子蟹、鋸緣青蟹等多種重要經濟物種(Small, 2012; Stentifordet al, 2015;Liet al, 2021)。該寄生甲藻主要寄生于海水甲殼類動物的血淋巴和組織血腔中; 感染后期, 寄生甲藻在宿主體內大量增殖, 導致宿主的心臟、肝胰腺、鰓等重要器官發生功能失調乃至喪失, 最終造成宿主死亡(Fieldet al, 1995; Tayloret al, 1996; Sheppardet al,2003)。近年來, 在我國浙江舟山、廣東汕頭、山東青島、河北黃驊等沿海地區的主要經濟蟹類養殖區均報道發現了血卵渦鞭蟲流行病發生, 且三疣梭子蟹(P.trituberculatus)、擬穴青蟹(Scylla paramamosain)、鋸緣青蟹(Scylla serrata)和脊尾白對蝦(Exopalaemon carinicauda)等我國主要經濟甲殼動物均可被感染(許文軍等, 2007a, 2007b; Liet al, 2008; Xuet al, 2010;劉順等, 2014; Wanget al, 2017)。近年來, 血卵渦鞭蟲的流行區域和宿主種類持續增加, 已成為影響海水經濟甲殼動物漁業養殖業可持續發展的新型流行性病原(Stentifordet al, 2005; Small, 2012; Liet al, 2021)。
課題組前期在日照地區采集的養殖梭子蟹樣品中發現存在血卵渦鞭蟲感染, 為進一步探究該區域夏季養殖三疣梭子蟹大規模死亡的原因和流行病發生過程, 本研究對日照沿海地區的主要梭子蟹養殖區域開展了系統的流行病學調查, 并對該區域內代表性養殖池塘進行了連續現場監測, 以解析該地區血卵渦鞭蟲流行病發生的生態學過程, 為防控三疣梭子蟹血卵渦鞭蟲流行病提供理論依據。
1 材料與方法
1.1 樣品采集
2020 年8 月, 在日照市主要三疣梭子蟹養殖區進行了血卵渦鞭蟲感染情況調查, 自東港區兩城鎮和經濟技術開發區的發病池塘分別采集了23和26 只養殖三疣梭子蟹成蟹, 于濕潤、低溫、充氧條件下運回實驗室,后根據檢測方法要求分別采集血淋巴、組織樣品進行血卵渦鞭蟲形態學觀察及組織病理檢測。
2021 年5~9 月, 在日照市兩城鎮一梭子蟹養殖池塘進行了血卵渦鞭蟲流行病發生過程連續監測調查(每月1~2 次), 分別采集60~100 只養殖梭子蟹個體, 置于低溫、潮濕泡沫盒中, 充氧后即刻運至實驗室, 進行類似處理、采集相應樣品; 同時在養殖池塘(設4 個采樣點)進行環境樣品采集, 自水面以下0.5 m 處用采水器采集2 L 水樣裝于樣品瓶中, 運至實驗室進行下一步處理。
1.2 養殖池塘環境因子測定
養殖池塘水體溫度和鹽度通過YSI 水質分析儀(EC300, EcoSense, 中國)現場測定。
葉綠素a樣品的處理與測定: 將現場采集的水樣通過真空泵負壓抽濾到Whatman GF/F 濾膜上, 濾膜用鋁箔包裹后于-80 °C 保存; 濾膜用90%丙酮萃取24 h 后, 使用Trilogy 熒光計(7200-000, Turner, 美國)測定上清液酸化前后的熒光值, 計算葉綠素a的質量濃度(中華人民共和國國家質量監督檢驗檢疫總局等, 2008a)。
營養鹽樣品的處理與測定: 將現場采集的水樣經 0.45 μm 孔徑的濾膜過濾, 加入氯仿固定, 于-80 °C 保存; 使用營養鹽連續流動分析儀(San Plus,Skalar, 荷蘭)測定硝酸鹽、亞硝酸鹽、銨鹽、硅酸鹽和磷酸鹽的質量濃度(中華人民共和國國家質量監督檢驗檢疫總局等, 2008b)。
1.3 血卵渦鞭蟲形態學觀察及組織病理檢測
采用中性紅染色血涂片法(Stentifordet al, 2005)對三疣梭子蟹中的血卵渦鞭蟲感染情況進行形態學檢測, 具體步驟如下: 采用一次性1 mL 無菌注射器從梭子蟹第五步足的關節膜處抽取約200 μL 血淋巴,將1~2 滴血淋巴液滴在載玻片上并與等體積的中性紅溶液(質量體積比為0.04%, 過濾海水)混勻, 蓋上蓋玻片于顯微鏡(BX53, Olympus)下觀察; 將剩余血淋巴加入到含有800 μL 無水乙醇的1.5 mL 離心管中固定, 于-20 °C 下保存, 用于DNA 提取。參照Wang等(2017), 基于200×顯微鏡視野下觀察到的血卵渦鞭蟲數量, 將梭子蟹感染程度認定為輕度(<10)、中度(10~100)、重度(>100)感染狀態。
將經血涂片法確認存在血卵渦鞭蟲感染的三疣梭子蟹解剖, 采集肝胰腺、心臟、鰓、肌肉等組織, 置于波恩氏液進行固定, 48 h 后轉移到70%酒精中保存。參考Wheeler 等(2007)的步驟處理固定的組織樣品, 經系列脫水后進行H&E(hematoxylin & eosin)染色, 切片經蓋片、封片后于顯微鏡下觀察梭子蟹感染血卵渦鞭蟲后的組織病理變化。
1.4 血卵渦鞭蟲的分子生物學鑒定及遺傳分析
使用海洋動物組織基因組 DNA 提取試劑盒(TIANGEN, 天津)提取1.3 采集的梭子蟹血淋巴樣品的基因組DNA, 于-20 °C 保存, 以待后續檢測使用。
采用血卵渦鞭蟲 ITS1 序列的特異性引物(F:5′CATTCACCGTGAACCTTAGCC3′; R: 5′CTAGTCAT ACGTTTGAAGAAAGCC3′)(Smallet al, 2007)對梭子蟹中的血卵渦鞭蟲感染情況進行分子PCR 檢測。PCR擴增反應體系為: 2×ApexHFFS PCR Master Mix(艾科瑞生物, 海南)10.0 μL, DNA 模板1.0 μL, 上、下游引物(10 μM)各0.8 μL, 無菌水7.4 μL。擴增程序為: 94 °C 預變性5 min; 94 °C 變性30 s, 56 °C 退火30 s, 72 °C 延伸30 s, 進行30 個循環; 72 °C 延伸10 min, 4 °C 保存。采用1.5%的瓊脂糖凝膠對PCR 擴增產物進行電泳檢測分析。挑選陽性PCR 產物進行瓊脂糖凝膠電泳并切膠回收, 送上海生工生物工程股份有限公司純化后進行TA克隆和Sanger 測序。將獲得的血卵渦鞭蟲ITS1 序列在NCBI 網站進行BLAST 分析, 并與GenBank 中對應的代表性ITS1序列進行比對, 運用Mega 5.05軟件基于最大似然法構建系統發育樹。
1.5 環境水體中血卵渦鞭蟲豐度的qRT-PCR 檢測
將現場采集的環境水樣(300 mL)通過真空泵負壓抽濾到孔徑為0.2 μm 的聚碳酸酯膜上, 濾膜置于2 mL凍存管中, 經液氮速凍后, 置于-80 °C 保存, 以待后續檢測使用。
使用Biomarker Soil Genomic Kit 試劑盒(百邁客生物科技, 北京)提取100 mL 環境水樣的環境基因組DNA, 參照Li 等(2010)的方法對水體中的血卵渦鞭蟲進行qRT-PCR 檢測。將含有ITS1 擴增子的質粒經梯度稀釋為102~109copies/μL, 進行qRT-PCR 檢測, 反應體系為: TB Green? Premix Ex Taq? Ⅱ(TaKaRa, 日本)10.0 μL, DNA 模板1.0 μL, 上、下游引物(10 μmol/L)各0.8 μL, 無菌水7.4 μL, 反應條件為: 95 °C, 30 s; 95 °C,5 s, 60 °C, 30 s, 進行40 個循環。所得擴增循環數(threshold cycle,CT)與每反應拷貝數的對數值通過線性回歸擬合為標準曲線。將樣品DNA 進行qRT-PCR 檢測,根據標準曲線計算得到每反應拷貝數的對數值, 根據單個血卵渦鞭蟲細胞中有300 個ITS1 序列拷貝數(Liet al, 2010), 將拷貝數換算為血卵渦鞭蟲細胞數, 估算環境水體中的血卵渦鞭蟲豐度。
1.6 數據分析
使用SPSS 23.0 軟件, 對不同時期水體中血卵渦鞭蟲的豐度是否存在差異進行單因素方差分析(oneway ANOVA,P<0.05), 并對梭子蟹感染率、水體中血卵渦鞭蟲豐度和環境因子進行Pearson 相關性分析。
2 結果與分析
2.1 日照地區血卵渦鞭蟲株系的鑒定分析
2.1.1 血卵渦鞭蟲形態學觀察及組織病理檢測 對三疣梭子蟹進行中性紅染色血淋巴涂片顯微觀察,健康梭子蟹個體血淋巴涂片中的細胞均為未被中性紅染色的宿主血細胞, 未見被中性紅染色的血卵渦鞭蟲細胞(圖1a)。染病梭子蟹個體的鏡檢結果顯示,除了未被中性紅染色的血細胞外, 血淋巴中充斥著大量近似球形的類變形蟲狀滋養體, 其直徑約為10~15 μm, 細胞內多個溶酶體吸收中性紅染液而呈亮紅色(圖1b)。
組織病理學觀察顯示, 染病梭子蟹個體的肝胰腺、心臟、肌肉、鰓等器官和組織中均發現存在血卵渦鞭蟲感染(圖2)。大多數血卵渦鞭蟲處于類變形蟲滋養體階段, 充斥在肝胰腺的肝小管間隙以及心肌和步足肌的肌束間隙中(圖2a, 2b, 2d), 鰓組織中也存在少量的血卵渦鞭蟲細胞(圖2c)。染病梭子蟹個體肌肉組織中的肌束間隙明顯增大, 肌絲斷裂溶解, 相鄰肌絲纖維互相分離; 肝胰腺的肝小管間隙和管腔增大, 結締組織嚴重缺失, 肝小管纖毛柱狀上皮結構發生溶解變性呈空泡狀; 鰓組織中除鰓絲末端發生膨大外, 未見其他明顯的組織病理變化。
感染梭子蟹的心臟、肌肉、肝胰腺、鰓等多個重要組織器官中能夠觀察到血卵渦鞭蟲的多種生活史階段, 包括絲狀滋養體(圖3a)、類變形蟲滋養體(圖2a)、蛛網狀滋養體(圖3b)、團塊狀聚合體(圖3c)、孢子前細胞和孢子(圖3d)。其中, 絲狀滋養體主要出現在處于感染早期的病蟹血淋巴或組織中; 類變形蟲滋養體是檢測中常見的生活史階段, 在感染早期和中期的病蟹血淋巴和組織器官中常被發現; 蛛網狀滋養體、團塊狀聚合體是血卵渦鞭蟲的主要增殖階段,主要出現于感染中、后期的病蟹組織中; 孢子前細胞、孢子是血卵渦鞭蟲在宿主體內生活史發展的末尾階段, 出現在感染晚期的病蟹組織中。
2.1.2 血卵渦鞭蟲分子生物學鑒定分析 通過分子克隆和測序方法, 分別從日照市2 個梭子蟹養殖區(東港區兩城鎮和經濟技術開發區)的感染梭子蟹樣品中獲得了長度約為~299 bp 的特異性HematodiniumITS1 rDNA 序列, 并提交至GenBank 數據庫(序列號分別為ON668054、ON668055 和ON668056)。NCBI數據庫BLAST 比對分析結果顯示, 該序列與數據庫中感染我國沿海甲殼類的H.perezi株系的ITS1 序列相似性為98%~100%, 系統發育分析結果也表明本研究獲得的ITS1 序列與感染我國海水蟹類的血卵渦鞭蟲ITS1序列聚為一枝(圖4), 同屬于H.perezi基因Ⅱ型。
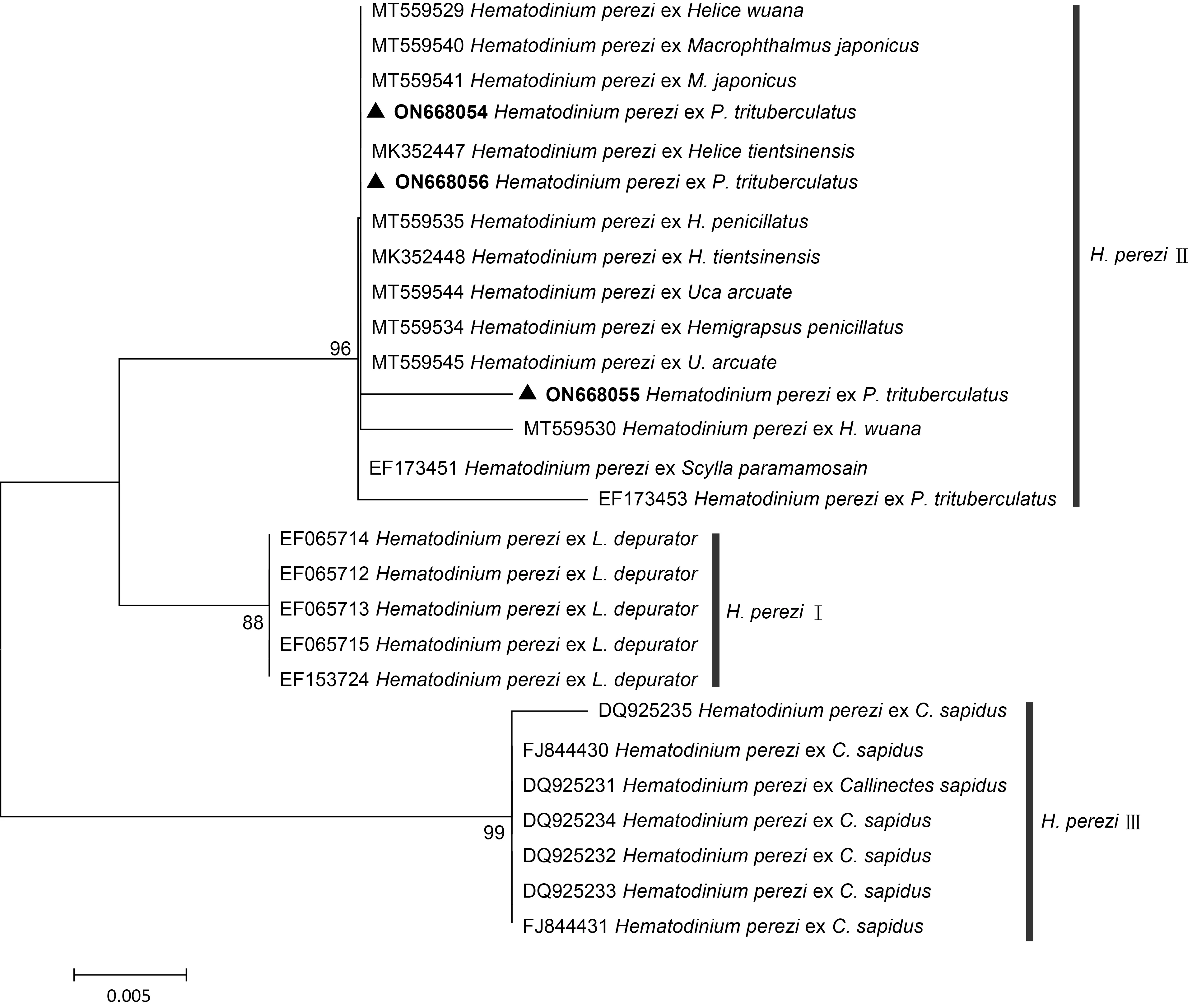
圖4 基于ITS1 序列構建的日照地區養殖池塘中感染三疣梭子蟹的血卵渦鞭蟲系統發育樹Fig.4 Molecular phylogenetic analysis of H. perezi infected P. trituberculatus in Rizhaoarea based on Hematodinium ITS1
2.2 三疣梭子蟹血卵渦鞭蟲感染狀況及感染率變化
2020 年8 月自日照市2 個養殖區域(東港區兩城鎮和經濟技術開發區)采集的三疣梭子蟹成體中的血卵渦鞭蟲感染率分別為52.2%和42.3%, 其中多數為中、重度感染。
2021 年5 至9 月, 我們對日照兩城鎮養殖池塘中梭子蟹的血卵渦鞭蟲感染情況進行了連續定期采樣檢測, 統計分析養殖期間梭子蟹的感染率和感染程度(圖5)。結果顯示, 5 月在梭子蟹幼苗中未發現血卵渦鞭蟲感染。6 月梭子蟹血卵渦鞭蟲感染率為24.5%,其中輕度感染個體占比較高, 占感染宿主的60.9%。7月中旬, 梭子蟹感染率上升至50.7%, 感染宿主中輕度感染個體比例下降, 中、重度感染個體比例升高,分別為47.2%、22.2%和30.6%。7 月底, 梭子蟹感染率為47.0%, 輕度感染個體比例升高, 占感染宿主的59.0%。8 月下旬, 梭子蟹感染率達到峰值, 為52.5%。感染宿主中, 輕度感染個體的比例出現下降, 由8 月中旬的55.6%下降至18.8%; 而中度和重度感染個體的比例升高, 分別由25.9%和18.5%升至43.7%和37.5%, 中、重度感染個體比例高達81.2%。9 月梭子蟹感染率下降至23.8%, 感染宿主中33.3%為輕度感染、6.7%為中度感染, 60.0%為重度感染。該養殖池塘中血卵渦鞭蟲流行病高發期為7 至8 月, 其中8 月下旬中、重度感染宿主占比最高。
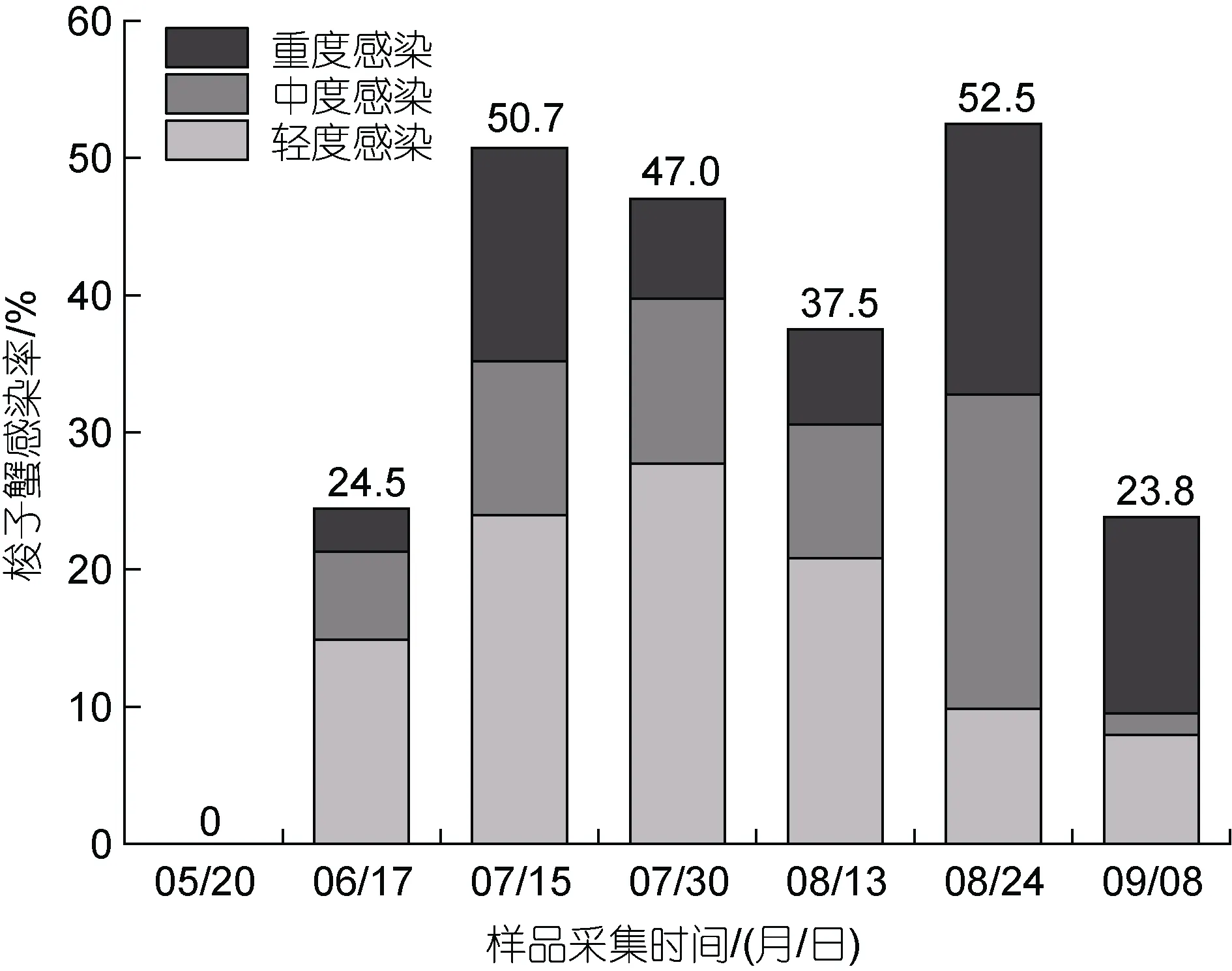
圖5 三疣梭子蟹血卵渦鞭蟲感染的流行情況Fig.5 Prevalence of H. perezi infection in P. trituberculatus
2.3 養殖池塘水體中血卵渦鞭蟲豐度變化
2021 年5~11 月, 在日照兩城鎮養殖池塘環境水體中檢測到血卵渦鞭蟲存在, 其豐度不同時間點有明顯變化(圖6)。養殖池塘水體中血卵渦鞭蟲的豐度在5 至6 月較高(血卵渦鞭蟲細胞數量1 194~1 271 cells/L)與7月中旬、8 月至11 月有顯著差異(P<0.05); 后在7 月中旬出現呈下降趨勢, 到下旬再次出現峰值(676 cells/L),隨后8 月中旬至11 月中旬, 血卵渦鞭蟲豐度在193~556 cells/L 之間波動, 但無顯著差異(P>0.05)。進一步分析梭子蟹血卵渦鞭蟲感染率變化與養殖池塘水體中血卵渦鞭蟲豐度變化的關聯性發現, 梭子蟹感染率的峰值出現在水體血卵渦鞭蟲豐度高值期之后(圖6)。
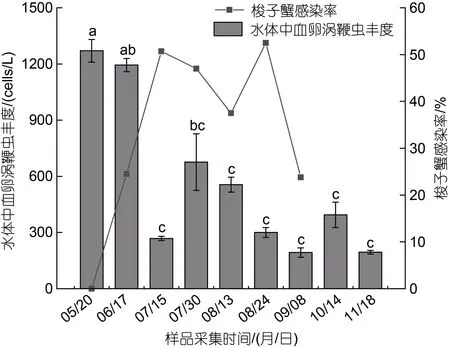
圖6 三疣梭子蟹血卵渦鞭蟲感染率與水體中血卵渦鞭蟲豐度的關聯性Fig.6 Correlation between the prevalence of H. perezi and the abundance of H. perezi
2.4 血卵渦鞭蟲流行病發生與環境因子關聯性
2021 年5 至11 月調查期間, 日照市兩城鎮養殖池塘水體主要環境指標變化概述如下: 水溫從5~6 月開始升高, 到7~8 月達到26.2~28.6 °C, 隨后開始降低。養殖水體鹽度變化范圍在20.7~29.6, 最大值為6月的29.6, 最小值為9 月的20.7。葉綠素a濃度變化范圍在2.2~13.3 μg/L, 在5 月、8~10 月維持在較高水平(8.0~13.3 μg/L), 峰值在8 月。硝酸鹽濃度變化范圍在0.5~694.3 μg/L, 峰值出現在9 月為694.3 μg/L。亞硝酸鹽濃度變化范圍在0~72.9 μg/L, 峰值出現在9月為72.9 μg/L。銨鹽濃度變化范圍在31.2~616.4 μg/L,峰值出現在8 月為616.4 μg/L。磷酸鹽濃度變化范圍在6.5~48.2 μg/L, 峰值出現在8 月為48.2 μg/L。硅酸鹽濃度變化范圍在521.4~2 735.0 μg/L, 峰值出現在9月為2 735.0 μg/L。
養殖池塘梭子蟹感染率和各因子之間進行Pearson 相關性分析, 結果顯示梭子蟹血卵渦鞭蟲感染率與溫度極顯著正相關(P<0.01), 梭子蟹感染率與溫度的變化趨勢基本一致, 7 月至8 月, 養殖池塘水溫較高時(>25 °C)發病率最高(圖7); 梭子蟹血卵渦鞭蟲感染率與其他生物及環境因子無顯著相關性(表1)。養殖池塘水體中血卵渦鞭蟲豐度和各因子之間的Pearson 相關性分析結果顯示, 水體中血卵渦鞭蟲豐度與鹽度顯著正相關(P<0.05), 兩者變化趨勢基本一致(圖8); 與亞硝酸鹽、銨鹽、磷酸鹽呈顯著負相關(P<0.05); 而葉綠素a、硝酸鹽、硅酸鹽與環境水體中的血卵渦鞭蟲豐度無顯著相關性(表1)。

表1 梭子蟹血卵渦鞭蟲感染率、水體中血卵渦鞭蟲豐度與環境因子相關性Tab.1 Correlation between the prevalence of H. perezi in P. trituberculatus and environmental factors, and correlation between the abundance of H. perezi in water and environmental factors
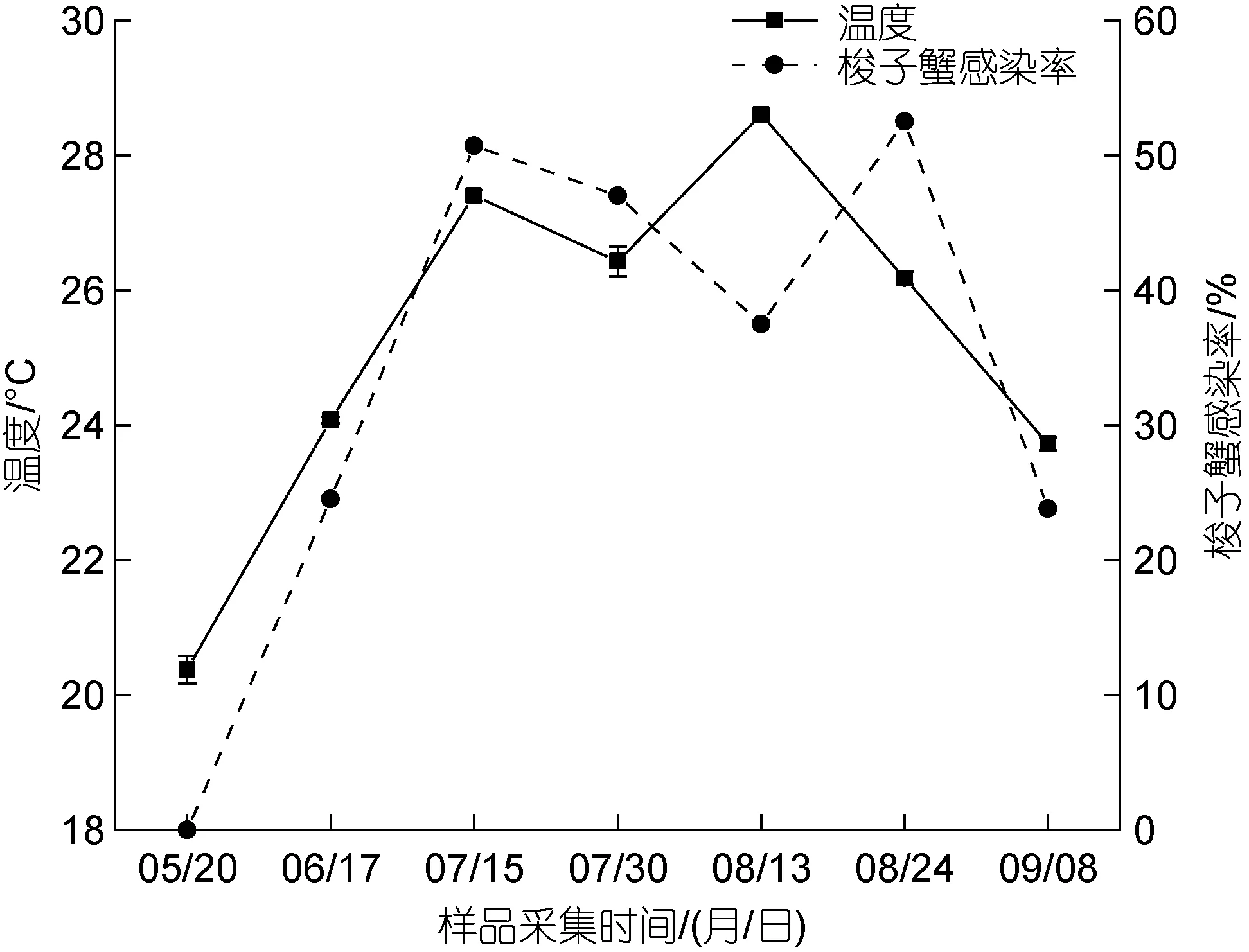
圖7 三疣梭子蟹血卵渦鞭蟲感染率與水體溫度的關聯性Fig.7 Correlation between the prevalence of H. perezi and the temperature
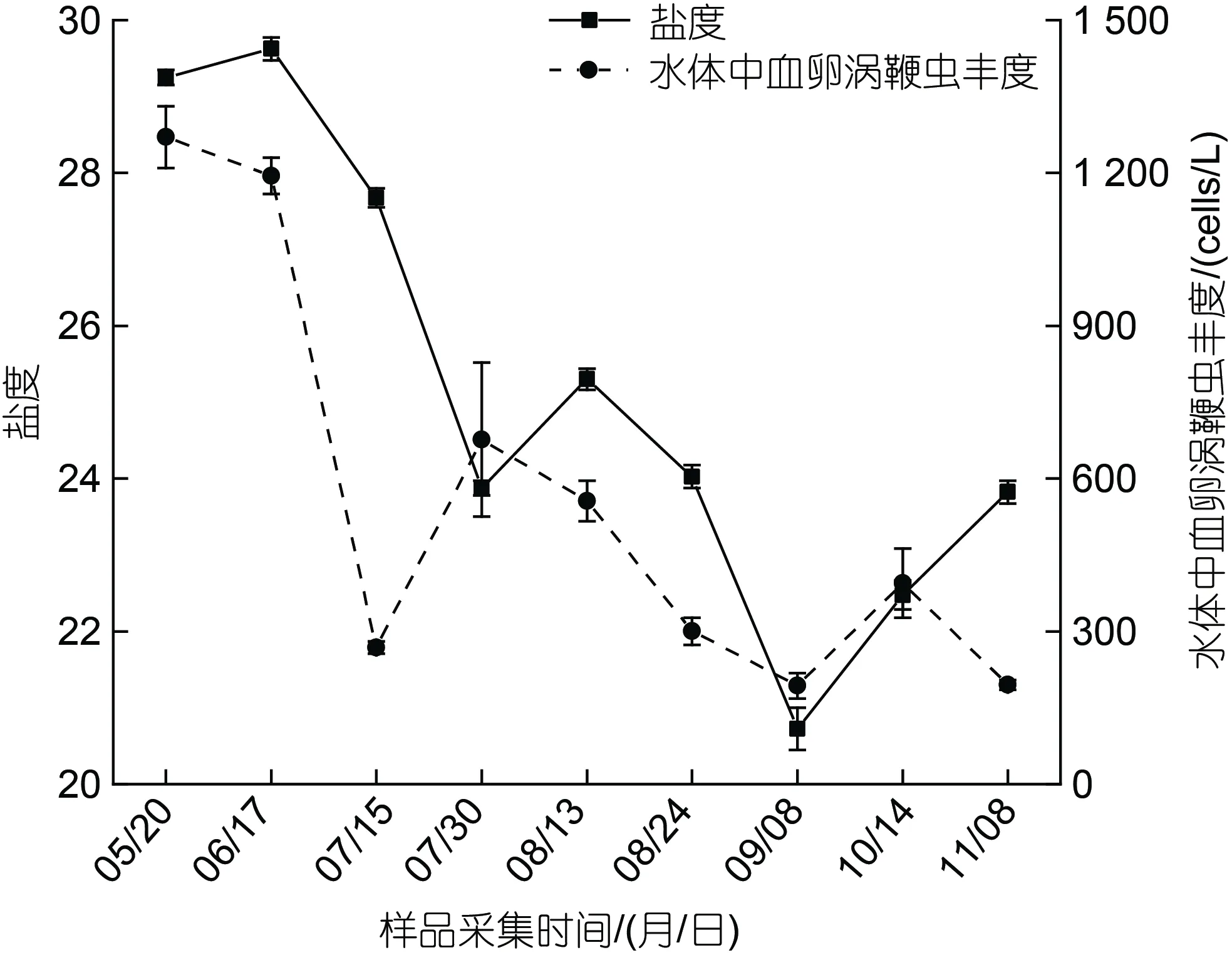
圖8 水體鹽度與水體中血卵渦鞭蟲豐度的關聯性Fig.8 Correlation between the salinity and the abundance of H.perezi in water
3 討論
本研究首次在日照市沿海地區的養殖三疣梭子蟹中報道發現了海洋寄生甲藻血卵渦鞭蟲感染。2020~2021 年在日照沿海地區的主要養殖區采集的“牛奶病”癥狀梭子蟹的血淋巴和器官、組織中均發現了包括絲狀滋養體、類變形蟲滋養體、蛛網狀滋養體、團塊狀聚合體、孢子前細胞和孢子等血卵渦鞭蟲的多個生活史階段。染病梭子蟹血淋巴中存在大量的血卵渦鞭蟲細胞, 尚存在少量的血淋巴細胞; 病蟹肝胰腺的肝小管間隙和管腔增大, 充斥著大量的血卵渦鞭蟲, 結締組織嚴重缺失; 心臟肌肉組織中的肌束間隙明顯增大, 肌絲斷裂溶解, 血竇中存在著大量的血卵渦鞭蟲。這些染病梭子蟹的主要器官和組織病理變化與我國沿海其他地區報道的血卵渦鞭蟲感染導致宿主的組織病理變化類似(許文軍等, 2007a, 2007b;王金鳳等, 2015; 王印庚等, 2017)。進一步的分子遺傳學分析結果顯示, 日照地區發現的血卵渦鞭蟲株系與感染我國沿海蟹類的血卵渦鞭蟲株系的序列相似性較高、親緣關系較近, 屬于我國沿海廣泛分布的主要致病株系Hematodinium perezi基因Ⅱ型(Smallet al,2012; Xiaoet al, 2016)。
該養殖區域血卵渦鞭蟲流行病主要發生在6~9月, 其中7~8 月為高峰期。養殖池塘中梭子蟹血卵渦鞭蟲感染率峰值與水體中血卵渦鞭蟲豐度變化呈現錯峰交替現象, 間隔時間(約3~4 周)基本與血卵渦鞭蟲生活史發展周期一致(Messicket al, 2000; Liet al,2011; Shieldset al, 2015)。5~6 月, 水體中血卵渦鞭蟲的豐度處于高值; 6 月, 在梭子蟹中檢測到血卵渦鞭蟲感染, 且輕度感染比例較高; 7 月中旬, 梭子蟹感染率升高至50.7%, 半數以上為中、重度感染。7 月中旬重度感染的個體數量較多, 而水體中血卵渦鞭蟲的豐度在7 月中旬至7 月底期間升高; 表明血卵渦鞭蟲已經在宿主體內完成生活史過程, 重度感染宿主將大量的血卵渦鞭蟲孢子釋放到環境水體中(Appletonet al, 1998; Liet al, 2010)。水體中的大量血卵渦鞭蟲孢子又引發了新一輪感染, 7 月底至8 月中旬又出現大量輕、中度感染個體, 至8 月下旬感染率再次達到峰值, 且多為重度感染個體。一般宿主從感染血卵渦鞭蟲到重度感染或死亡的高峰期需經過一個月左右時間(Messicket al, 2000); 據此認為6 月和7 月中下旬為該區域血卵渦鞭蟲傳播擴散和梭子蟹大量感染的關鍵時期。
血卵渦鞭蟲流行病的發生與水體溫度和鹽度變化密切相關。本研究發現, 梭子蟹血卵渦鞭蟲感染率與溫度極顯著正相關(P<0.01), 梭子蟹血卵渦鞭蟲感染率在7~8 月養殖池塘水溫較高(26.2~28.6 °C)時達到峰值。血卵渦鞭蟲流行病發生具有季節性特征, 其發生受溫度影響(Briggset al, 2002; Shieldset al, 2015;Huanget al, 2021)。在美國蘭蟹中, 血卵渦鞭蟲能夠在20~24 °C 下迅速發展(Messicket al, 2000), 其流行的最適溫度是25 °C(Huchin-Mianet al, 2018)。分離自山東沿海的血卵渦鞭蟲可以在15~25 °C 下在宿主體內正常發育, 且溫度越高, 發展越快, 25 °C 時能迅速增殖發育(呂曉陽等, 2022)。因此, 認為是7~8 月適宜的溫度促進了養殖池塘血卵渦鞭蟲流行病的爆發。本研究中, 水體中血卵渦鞭蟲的豐度與鹽度顯著正相關(P<0.05)。7 月底至9 月, 水體鹽度較低(20.7~25.3), 大致呈降低趨勢, 與水體中血卵渦鞭蟲豐度變化趨勢基本一致。血卵渦鞭蟲孢子可以在20~35 鹽度的環境中存活3~7 天(Liet al, 2011), 其生存最適鹽度是30(Huchin-Mianet al, 2018), 其活性在10~20 鹽度中有明顯降低(Coffeyet al, 2012)。因此, 認為鹽度降低抑制了水體中血卵渦鞭蟲存活, 進而限制了血卵渦鞭蟲傳播擴散。
本研究在放苗期(5 月)的三疣梭子蟹蟹苗(C2~C4期)中未發現陽性感染個體, 而在后續養殖過程中采集的梭子蟹成蟹中陸續發現了血卵渦鞭蟲感染, 表明該區域養殖環境中存在血卵渦鞭蟲的病原來源。此前研究也發現, 養殖池塘及周邊環境中的野生宿主的是養殖梭子蟹血卵渦鞭蟲流行病的重要病原來源(Huanget al, 2021), 孢子水體傳播是養殖池塘內血卵渦鞭蟲流行病傳播擴散的主要途徑(Shieldset al,2017; Huanget al, 2019, 2021)。本研究環境因子相關性分析顯示, 水體中血卵渦鞭蟲豐度與亞硝酸鹽、銨鹽、磷酸鹽也呈顯著負相關關系, 氮、磷營養鹽的變化會影響環境微生物群落結構(Weiet al, 2016)。然而,血卵渦鞭蟲屬于海洋寄生甲藻類群, 其生活史過程主要在甲殼動物宿主內完成, 在感染晚期釋放游動孢子到環境水體中(Appletonet al, 1998; Liet al,2010), 微生物群落結構的改變有可能對孢子的存活產生一定的影響。因此, 進一步明確養殖區域血卵渦鞭蟲的病原來源, 揭示關鍵環境因子對于其流行病發生的影響機理是后續研究的重點, 可為養殖生產中有效防控該流行性提供理論依據和參考。
4 結論
本研究首次報道發現了日照地區養殖三疣梭子蟹感染寄生甲藻血卵渦鞭蟲及其流行病發生情況。該區域血卵渦鞭蟲流行病主要發生在6~9 月間, 7~8 月為暴發期, 發病池塘中梭子蟹感染率超過50%。梭子蟹血卵渦鞭蟲感染率變化與水體中血卵渦鞭蟲豐度變化密切相關, 梭子蟹感染率峰值出現在水體血卵渦鞭蟲豐度高值的3 至4 周之后。養殖池塘中血卵渦鞭蟲流行病的發生與水體溫度和鹽度密切相關, 7~8月較高的水溫(26.2~28.6 °C)促進了血卵渦鞭蟲在宿主體內快速增殖及流行病暴發, 而較低的鹽度(20.7~25.3)可能抑制了水環境中血卵渦鞭蟲的存活及其傳播擴散。
致謝 感謝日照市春祥水產合作社及黃海水產研究所謝國駟博士在現場調查及實驗過程中給予的幫助和支持。
