棉花纖維優勢表達基因GhSLD1啟動子的克隆和功能分析
2023-10-25 10:23:48劉芳徐夢貝王巧玲孟倩李桂名張宏菊田惠丹徐凡羅明
中國農業科學 2023年19期
關鍵詞:煙草
劉芳,徐夢貝,王巧玲,孟倩,李桂名,張宏菊,田惠丹,徐凡,羅明
棉花纖維優勢表達基因啟動子的克隆和功能分析
劉芳,徐夢貝,王巧玲,孟倩,李桂名,張宏菊,田惠丹,徐凡,羅明
西南大學生物技術中心/農業農村部生物技術與作物品質改良重點開放實驗室,重慶 400716
【目的】棉花纖維是棉花的主要經濟產品,是由胚珠外珠被表皮細胞經極性伸長和次生壁加厚而成的單細胞。棉花纖維細胞是最長的植物細胞之一,是研究植物細胞生長發育的理想材料。鑒定纖維細胞特異或優勢表達啟動子可為纖維發育的基礎研究提供控制目標基因表達的調控序列,為改良纖維性狀的分子育種提供依據。【方法】克隆纖維細胞優勢表達基因的啟動子,通過啟動子序列分析網站PlantCARE分析克隆序列中包含的重要順式調控元件。根據部分重要順式調控元件的分布,對克隆啟動子片段進行5′-端刪除,共獲得4個啟動子片段,并構建了相應的植物表達載體。利用構建的植物表達載體進行煙草和棉花的遺傳轉化,通過轉基因煙草和棉花的分子鑒定明確轉基因植株。并檢測轉基因植株不同組織器官、纖維細胞不同發育時期的GUS活性。【結果】克隆獲得最長啟動子片段為2 900 bp,除了包含多個啟動子必備轉錄調控元件外,還包含多個脫落酸響應元件、厭氧誘導元件、茉莉酸甲酯響應元件、油菜素內酯響應元件、種子特異調控元件、脅迫響應元件和MYB轉錄因子結合位點。通過5′-端刪除獲得長度分別為2 900()、2 178()、1 657()和1 232 bp()4個啟動子片段,經分子鑒定,獲得4個片段的轉基因煙草,在轉基因煙草中,、和不表達,而廣泛表達,表達強度與CaMV 35S啟動子相似。和差異序列中包含4個脫落酸響應元件、2個油菜素內酯響應元件和3個MYB結合位點,這些順式調控元件可能與、和在轉基因煙草中不表達有一定關系。經分子鑒定,獲得轉基因棉花。在轉基因棉花纖維中優勢表達,在轉基因花粉中有較低的表達,在其他組織器官中幾乎不表達。在纖維細胞的生長發育過程中,在纖維細胞早期生長階段(5 DPA)表達較低,在纖維細胞伸長期(10—15 DPA)表達水平相對較高,在纖維細胞次生壁合成期(20—30 DPA)表達水平有所降低。【結論】啟動子是一個廣泛表達啟動子,啟動子是一個纖維細胞優勢表達啟動子,在纖維細胞伸長期表達量相對較高。可應用于棉花纖維發育相關基因的功能研究和改良纖維性狀的分子育種。
棉花;啟動子;功能分析;;鞘脂delta8-去飽和酶
0 引言
【研究意義】棉花是世界上最重要的天然纖維作物,中國是棉花生產和消費大國,棉花生產在中國國民經濟中占有重要的地位。傳統的育種方法曾在棉花品種改良上取得較大的成功,但近年來,改良棉花產量和品質的育種進入平臺期。利用現有的遺傳資源和傳統育種手段難以再大幅度提高棉花產量。基因工程方法具有后代易于穩定,育種周期短等優點,可以打破物種間的遺傳障礙,實現優良目的基因的定向轉移,是改良棉花產量和纖維品質的有效途徑。但基因工程在作物改良中的作用發揮取決于3個方面的進展程度:目標基因的功能、調控目標性狀形成的分子機理和控制目標基因在特定部位和特定時間表達的特異啟動子。在棉花纖維品質和產量的基因工程改良中,常常需要在纖維細胞中超量或抑制一些基因的表達,來達到提高產量或改良品質的目的。在棉花纖維發育的分子機理研究中,也需要纖維細胞特異表達啟動子來上調或下調目標基因的表達,進而分析其在纖維細胞中的功能。最大限度地減少目標基因對其他組織和器官的不良影響。因此,棉花纖維特異或優勢表達啟動子的克隆和功能分析,對棉花纖維發育相關基因功能研究和棉花纖維改良的基因工程具有重要的理論意義和應用價值。【前人研究進展】啟動子是一段能被RNA聚合酶識別并結合的,位于結構基因5′上游的DNA序列,能活化RNA聚合酶并指導相應類型的RNA聚合酶與模板的正確結合,決定轉錄的方向和效率,控制基因表達(轉錄)的起始時間、空間和表達的強度。最早在棉花中克隆和功能分析的啟動子是的啟動子,的2.5 kb啟動子序列能夠指導報告基因在5—28 DPA(day post anthesis)的纖維中表達,在15—22 DPA表達量最高,而在其他組織中不表達[1-2]。說明啟動子是纖維發育早期特異表達的啟動子。隨后,于曉紅等[3]從亞洲棉中克隆了纖維特異表達基因的5′端約1.5 kb的上游序列,通過轉基因煙草證明該啟動子能夠驅動報告基因在煙草的根、莖、葉、表皮毛及維管組織中表達,說明該啟動子不具備纖維細胞特異表達特性。吳靄民等[4]進一步驗證了614 bp啟動子的功能,發現只在煙草葉片的表皮毛中檢測到報告基因表達,推測614 bp的啟動子序列已足夠驅動基因在棉花纖維中特異表達。Rinehart等[5]克隆了纖維發育中后期特異表達基因的啟動子并進行了5′刪除試驗。瞬時表達結果表明,的5′端2.3 kb片段已經具備啟動子活性,其表達活性約為組成型強啟動子CaMV 35S啟動子的1/3,是早期特異表達的啟動子的3倍。3個脂轉移蛋白基因(、和)在纖維伸長期特異表達。通過轉基因煙草分析其啟動子活性,結果顯示,報告基因僅在煙草表皮毛中表達。啟動子的啟動活性較弱,僅為CaMV 35S啟動子的1/1000左右。啟動子的活性強于啟動子[6-10]。棉花纖維素合成酶4亞基基因()在纖維次生壁沉積起始期和沉積期表達。該基因2.6 kb啟動子能夠指導報告基因在纖維細胞次生壁合成期表達[11-12]。棉花肌動蛋白基因()0.8 kb啟動子序列能夠指導報告基因在纖維中高量表達[13]。2個的啟動子(1 679 bp)和(1 285 bp)能驅動在纖維細胞中高量表達[14]。啟動子(1 000 bp)僅在轉基因棉花苞葉和花藥中表達[15]。(seed coat and fiber protease)啟動子(1 006 bp)指導報告基因在纖維細胞中特異表達,是一個纖維細胞發育早期的特異啟動子[16]。啟動子在棉花纖維發育過程中優勢表達,且表達活性較高,該啟動子活性受赤霉素誘導和脫落酸抑制[17]。是一個纖維伸長期優勢表達基因,其啟動子(961 bp)指導在0—20 DPA纖維以及柱頭、雄蕊、萼片、胚根、子葉和根中表達[18]。這些研究結果為解析目標基因的功能和調控機制提供了重要數據,也為改良纖維產量和品質的分子設計提供更多可選擇的調控序列。但迄今為止,可選擇的纖維細胞特異或優勢表達的啟動子還十分有限。【本研究切入點】棉花纖維細胞中特異/優勢表達啟動子的研究雖然取得了一定進展,但由于棉花遺傳轉化比較困難,多數在纖維細胞中特異/優勢表達基因的啟動子還沒有通過轉基因棉花進行功能鑒定。部分啟動子通過轉基因擬南芥或煙草進行了功能鑒定,但僅是間接地揭示了目標啟動子的功能。同時,由于纖維細胞的生長發育包含纖維細胞起始、伸長和次生壁沉積等不同發育過程,不同時空和不同表達效率的啟動子還十分缺乏。鞘脂(sphingolipid)在棉花纖維細胞生長發育中具有重要作用,但不同的鞘脂分子在棉花不同組織、器官和細胞中的積累差異較大[19-21]。與此相對應,參與鞘脂分子修飾的鞘脂合成酶基因的表達也具有明顯的組織器官特異性。鞘脂delta8-去飽和酶(GhSLD1)介導LCB(long chain base)鏈C-8的去飽和反應,在棉花纖維細胞中優勢表達。【擬解決的關鍵問題】本研究通過克隆啟動子序列和構建不同長度啟動子(5′-刪除)植物表達載體,利用轉基因煙草和棉花,分析啟動子的功能,證明具有纖維細胞表達特異性,為棉花纖維發育基礎研究和現代育種分子設計提供新的纖維特異表達啟動子。
1 材料與方法
1.1 試驗材料
棉花遺傳轉化受體材料為棉花品種冀棉14(J14),由河北農業大學馬峙英教授所贈,西南大學生物技術中心保存。煙草遺傳轉化受體為cv.xanthi,由西南大學生物技術中心保存。
1.2 克隆GhSLD1的啟動子序列
位于陸地棉D亞基因組(),利用的cDNA序列,搜索已公布的棉花D-亞基因組序列(http://www.phytozome.net)。獲得的5′-上游調控序列,進而在ATG上游約3.0 kb處設計特異引物-up(5′-ATTCTCCACC ACATGCAAACC-3′)和在近ATG位點處設計特異引物-down(5′-GGCTTGATCATGTCTAG ACTC-3′),以冀棉14基因組DNA為模板進行擴增。擴增產物回收,與克隆載體pMD19(TaKaRa,中國大連)連接,經轉化、驗證和測序,獲得擴增片段長度為2 900 bp,并命名為。采用PlantCare數據庫(http://bioinformatics.psb.ugent.be/webtools/ plantcare/html/)分析啟動子順式調控元件。為了進行啟動子5′-端刪除試驗,根據啟動子順式調控元件分析結果,分別設計特異引物(-up:5′- cctaaagcaaccacatgcttc-3′,-up:5′-catcacgtgacatccttctct-3′,-up:5′-ttaggtgatgaatggcacgac-3′)。以為模板進行擴增,分別獲得2178 bp、1 657 bp和1 232 bp。
1.3 植物表達載體構建
用dⅢ和Ⅰ內切酶從pMD19--—載體上酶切不同長度的啟動子片段,與經dⅢ和Ⅰ雙酶切的pBI121載體片段連接,從而使啟動子片段置換了pBI121載體中的CaMV35S啟動子。通過酶切驗證,構建植物表達載體pBI121--—::GUS。
1.4 棉花的遺傳轉化
參照LUO等[22]棉花遺傳轉化方法進行棉花遺傳轉化。取50顆剝去外殼的冀棉14種子,放入一個三角瓶中,先用75%乙醇滅菌1 min,再用3%過氧化氫滅菌10 min,滅菌期間用手不斷搖晃三角瓶;用無菌水沖洗8—10次,直至將泡沫洗凈;在三角瓶中加入約50 mL無菌水,置于搖床,早晚各換一次無菌水;1 d之后就可以選擇長出根的種子,并將其插入種子萌發培養基上,于26 ℃暗培養1—2 d,選擇適宜的下胚軸進行遺傳轉化。將棉花下胚軸切成小段,放入制備好的農桿菌菌液,于26 ℃ 90 r/min浸染40—60 min;侵染后的下胚軸段轉移到棉花共培養基上,于26 ℃暗培養2 d之后,將其轉移到下胚段篩選培養基中,15 d后轉移到繼代培養基直到大量長出愈傷組織;將其轉移到棉花胚性愈傷誘導培養基中;當大量出現胚性愈傷時,可進行胚性愈傷懸浮培養,于26 ℃ 120 r/min搖床培養15 d左右;將體胚鋪于體胚伸長培養基上,體胚轉入體胚伸長培養基;待體胚伸長到1—2 cm后,轉入生根培養基上(繼代培養基、胚性愈傷誘導培養基、生根培養基每隔15 d繼代一次)。抗性幼苗在培養基中長大后,栽種于溫室培養缽。
1.5 煙草的遺傳轉化
采用農桿菌介導法進行煙草的遺傳轉化。取煙草種子用1%次氯酸鈉滅菌,再用無菌水沖洗5—8次,三角瓶中加入約50 mL無菌水,置于搖床培養2 d后種子至露白,然后轉移至萌發培養基,約1個月可取葉片進行遺傳轉化。在無菌條件下將葉片除去主葉脈,切成0.5 cm2大小的葉片,轉移到準備好的菌液中,于26 ℃ 90 r/min搖床浸染30—50 min,將浸染后的葉片轉移到煙草共培養基上,于26 ℃暗培養2 d后,將其轉移到煙草篩選培養基中,10 d后轉移到繼代培養基直到大量長出不定芽,將其切取轉移到生根培養基中。幼苗長大后栽種于溫室培養缽。
1.6 植物gDNA的提取
依據北京艾德來生物科技公司的新型植物基因組DNA(gDNA)快速提取試劑盒操作說明書提取植物基因組DNA,并進行瓊脂糖凝膠電泳檢測。
1.7 棉花RNA的提取
選取新鮮棉花材料(根、莖、葉、花、開花后0 d的胚珠和纖維至開花后6 d的胚珠和纖維、開花后6 d的纖維至開花后20 d的纖維、開花后6 d的胚珠至開花后20 d的胚珠),利用北京艾德來(Aidlab,中國)生物科技公司的EASYspin植物RNA快速提取試劑盒(貨號:RN09)提取各個樣品的總RNA,具體操作按說明書進行。
1.8 基因表達分析
將上述RNA合成cDNA(TakaRa),-20 ℃保存。運用實時熒光定量PCR擴增分析的表達水平。反應體系包括10 μL擴增混合緩沖液(試劑盒提供)、5′-端和3′-端引物各1 μL(5 μmol·L-1)、1 μL cDNA,去離子水補至20 μL。反應程序為94 ℃ 3 min;94 ℃ 30 s,54 ℃ 30 s,72 ℃ 30 s,40個循環。用棉花作內參基因,引物為(5′-ATGCCCAAGGACATCCAGTTG-3′)和(5′-CCTACCACTACCATCATGGCT-3′)。的引物為-1(5′-GATACAGAGTGG TTGGATAGG-3′)和-2(5′-GATCCTAGCAA AGCACATGAC-3′)。
1.9 轉基因煙草和棉花的分子鑒定
提取轉基因煙草和棉花抗性苗基因組DNA,以特異引物-up、-up、-up和-up分別和結合5′-端的3′引物-down(5′-ATCGAAACGCAGCACGATACG-3′)擴增抗性煙草或棉花基因組DNA,電泳檢測。
1.10 轉基因棉花中GUS活性的檢測
隨機選取5株PCR陽性轉基因棉花植株進行全面的β-葡萄糖苷酸酶(GUS)活性檢測。樣品中加適量植物組織GUS染液,37 ℃避光處理1 h,每隔一定時間更換一次75%酒精進行脫色。脫色完全后,在體視鏡下拍照記錄。
2 結果
2.1 GhSLD1的表達分析
為了分析鞘脂在棉花纖維細胞生長發育中的功能,從陸地棉中克隆了擬南芥鞘脂delta8-去飽和酶(SLD)基因的同源基因。首先檢測了該基因在棉花不同組織器官和纖維胚珠不同發育時期的表達特性(圖1)。結果顯示,主要在纖維細胞中表達,在胚珠早期發育階段(6—12 DPA)有較低的表達,在根、莖、葉、花以及開花當天胚珠和14—20 DPA胚珠中幾乎檢測不到表達信號。在不同發育時期的纖維細胞中,該基因的表達峰值出現8 DPA和10 DPA,隨后,表達水平快速降低。在14—20 DPA纖維細胞中維持較低的表達水平。說明是一個纖維細胞優勢表達基因,在纖維細胞不同發育時期,該基因的表達水平有明顯差異,在纖維細胞快速伸長期的表達水平最高。

圖1 GhSLD1在棉花不同器官和不同發育時期纖維胚珠中的表達特征
2.2 GhSLD1啟動子的克隆和序列分析
基因的啟動子是調控基因表達的主要元件,棉花纖維細胞特異或優勢表達調控序列是棉花纖維發育研究和現代分子設計育種的重要序列。是一個在纖維細胞快速伸長期優勢表達的基因,為了深入研究的表達特性和調控機制以及獲得纖維細胞特異或優勢表達的調控序列,首先克隆的啟動子并對克隆序列進行了分析。以(3)的5′-上游序列為參考,設計特異引物,從陸地棉(冀棉14)基因組中克隆獲得約3 000 bp的片段(圖2),通過克隆測序顯示擴增片段長2 900 bp。將該序列導入PlantCARE數據庫(http://bioinformatics.psb.ugent.be/webtools/plantcare/ html/)進行順式調控元件分析(表1),結果顯示,序列中除具有一般啟動子所具有的大量TATA框和CAAT框,以及較多的光反應元件外,還具有許多特異性響應元件,如脫落酸響應元件(CACGTG/ ACGTG)、厭氧響應元件(AAACCA)、油菜素內酯響應元件(CANNTG)、茉莉酸響應元件(CGTCA)、MYB結合位點(AACCTAA/TAACCA/CAACCA/ TAACTG),防御與脅迫響應元件(ATTCTCTAAC)等(表1)。說明的表達受到多種因子的調控。

M:DNA marker;GhSLD1-P1:GhSLD1的啟動子P1

表1 GhSLD1-P1啟動子序列中的順式調控元件
2.3 啟動子功能分析及植物表達載體的構建
為了尋找啟動子的核心區段,根據重要調控元件的分布,進行5′-刪除試驗。共獲得4個片段,長度分別為2 900()、2 178()、1 657()和1 232 bp(),分別構建啟動子植物表達載體(圖3)。由于許多研究者認為棉花纖維細胞的生長發育及其調控機制與植物表皮毛相似[23-26],為了探究啟動子各區段的功能,將構建的表達載體先進行煙草的遺傳轉化。

LB:T-DNA區段左邊界;RB:T-DNA區段右邊界;NPTII:卡那霉素抗性基因;GUS:b-葡萄糖酸苷酶基因(報告基因);Ter:終止子;CaMV 35S:花椰菜花葉病毒(CaMV)35S啟動子
2.4 啟動子轉基因煙草的鑒定
利用構建的植物表達載體進行煙草的遺傳轉化。待抗性再生苗移栽成活后,提取再生苗基因組DNA,利用特異引物-up、-up、-up和-up分別和結合5′-端的3′引物GUS-down擴增煙草基因組DNA(圖4)。結果表明,各個植物表達載體的轉基因煙草都擴增出特異條帶,該擴增帶與各自載體質粒的擴增帶大小一致,而以非轉基因煙草基因組DNA為模板的陰性對照和以水為模板的空白對照沒有任何特異擴增產物。表明包含不同啟動子長度的T-DNA區段整合到煙草基因組DNA中,獲得轉基因煙草植株。
2.5 啟動子轉基因煙草GUS活性分析
為了明確不同長度的啟動子在煙草中的表達模式,對不同啟動子轉基因煙草的根、莖、葉、花等不同部位進行GUS組織化學染色(圖5)。結果表明,在、和轉基因煙草的各部位均未檢測到GUS活性。在轉基因煙草各部位均檢測到GUS活性,包括葉片和莖的表皮毛中也具有較強的GUS信號。說明1 232 bp的啟動子序列具有啟動子的必要元件,可以控制基因的表達,但不具有表達特異性。-2 900— -1 232 bp序列中可能具有調控特異性表達的元件。由于也沒有GUS染色信號,進一步分析了-1 657—-1 232 bp間的序列特征,該序列包含4個脫落酸響應元件(G-box,ACGTG),同時與2個油菜素內酯響應元件(E-box,CANNTG)重疊;包含3個MYB結合位點(AACCTAA、CAACCA和TAACCA),這些順式調控元件可能與、和在轉基因煙草中沒有明顯GUS信號有一定關系。特別是全長啟動子中共有4個脫落酸響應元件(表1),都集中在這段序列中,而中沒有脫落酸響應元件,推測該元件與表達特性有密切關系。

A:GhSLD1-P1轉基因煙草;B:GhSLD1-P2轉基因煙草;C:GhSLD1-P3轉基因煙草;D:GhSLD1-P4轉基因煙草。M:DNA Marker;+:陽性對照,以對應片段的啟動子表達載體質粒為擴增模板;-:陰性對照,以野生型煙草gDNA為擴增模板;H2O:空白對照,以水為PCR擴增模板;1—4:對應片段的啟動子轉基因煙草植株

CaMV 35S:組成型強啟動子CaMV35S轉基因煙草;GhSLD1-P1— GhSLD1-P4:分別為GhSLD1-P1、GhSLD1-P2、GhSLD1-P3和GhSLD1-P4轉基因煙草。A:柱頭,B:雄蕊,C:葉片,D:葉柄橫切,E:莖橫切,F:莖縱切,G:花瓣,H:開花當天的子房,I:根
2.6 轉基因棉花的鑒定
結合前面的轉基因煙草GUS活性的結果,將對進行進一步探索。利用不同長度啟動子控制的植物表達載體和根癌農桿菌介導的棉花遺傳轉化,獲得的卡那(kan)抗性再生苗。進一步提取再生苗基因組DNA,以此為模板進行PCR擴增驗證,擴增引物分別結合于的5′序列和5′-端序列(圖6)。結果表明,轉基因棉花植株中能夠擴增出一條與陽性對照一致的帶,而陰性對照和空白對照均無任何擴增帶。說明植物表達載體的T-DNA區段整合到了棉花基因組中。
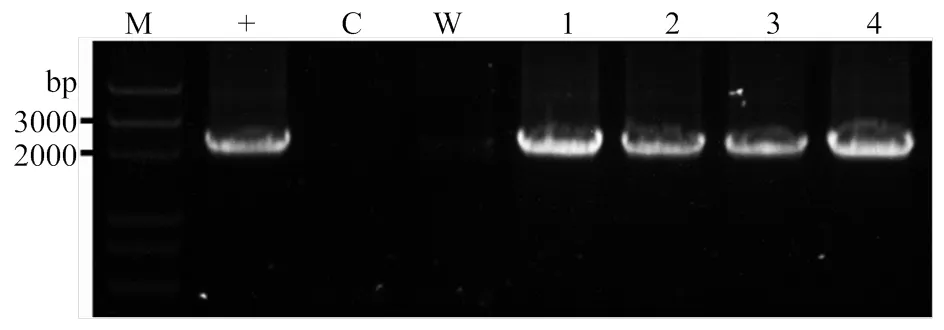
M:DNA Marker;+:陽性對照(以GhSLD1-P2植物表達載體質粒為模板);C:陰性對照(以野生型基因組DNA為模板);W:空白對照(以水為模板);1—4:轉基因棉花株系1—4
2.7 轉基因棉花中GUS活性分析
為了分析轉基因棉花中GUS活性,對不同組織器官和纖維細胞發育不同時期進行組織化學分析,并以組成型強啟動子CaMV 35S控制的轉基因棉花為陽性對照,以非轉基因棉花為陰性對照(圖7—圖9)。結果顯示,在轉基因棉花植株的根、莖、葉中均未檢測到GUS信號,在開花當天(0 DPA)也未檢測到GUS信號,僅在花粉中有較弱GUS信號(圖7和圖8)。為了明確啟動子在不同發育時期的纖維細胞中的表達特性,檢測5、10、15、20、25和30 DPA纖維細胞中GUS信號(圖9)。結果顯示,纖維細胞中有較強的GUS信號,相對而言,5 DPA中的信號較弱,10和15 DPA中的GUS信號最強,隨后GUS信號有所降低。說明主要在纖維細胞中表達,其次在花粉中有較低水平表達,是一個纖維細胞優勢表達啟動子,而且在纖維細胞伸長期表達水平最高。
3 討論
3.1 GhSLD1-P2啟動子在棉花纖維基礎研究和分子育種中有重要應用
啟動子是基因組中控制基因表達的主要序列。分析目標基因啟動子的功能不僅有助于解析目標基因的表達特性和表達調控機制,而且能夠獲得具有特殊時空表達特性的啟動子。在基因功能研究和改良作物性狀的分子設計中具有重要作用。傳統基因工程中常使用組成型強啟動子CaMV 35S,但過度的表達水平和不適的表達時空常常帶來副作用,即使改良了目標性狀,也嚴重干擾了其他生產性狀。因此,啟動子的功能鑒定是作物分子生物學的重要方面。

Non-T:陰性對照,非轉基因棉花;8DP2-GUS:pBI121-GhSLD1-P2::GUS載體轉基因棉花;35S-GUS:陽性對照,pBI121-CaMV35S::GUS載體轉基因棉花。下同

圖8 轉基因棉花花器官中的GUS活性檢測

5—30 DPA:開花后5—30 d的胚珠和纖維細胞 5-30 DPA: The ovules and fiber cells at 5-30 day post anthesis
棉花纖維是棉花的主要經濟產品,是由胚珠外珠被表皮細胞經極性伸長和次生壁加厚而成的單細胞纖維。纖維細胞的生長發育包含起始、伸長、次生壁沉積和脫水成熟4個階段,具有一個植物細胞生長和衰老的全過程。纖維細胞中的基因表達和物質代謝具有相對獨立的體系。因此,研究纖維發育相關基因的功能和調控機制迫切需要纖維細胞特異或優勢表達啟動子,以控制目標基因在纖維細胞中表達而不影響或少影響棉花植株的生長發育;同時,也使轉基因材料更具有可用性。如利用種皮特異啟動子控制生長素合成酶基因iaaM能夠促進纖維細胞起始[27]。利用CaMV 35S控制棉花類固醇5α-還原酶基因()難以獲得正常生長的轉基因棉花,而利用FBP7控制能夠獲得生長正常的轉基因棉花,且能夠促進纖維細胞的起始和伸長[22]。本研究中,啟動子在轉基因棉花纖維細胞中優勢表達,且在纖維細胞伸長期(10—15 DPA)的表達水平相對較高;同時,與CaMV 35S相比,該啟動子的總體表達水平較低。這些表達特性更有利于在纖維細胞中適時適量地調控目標基因的表達變化。
3.2 GhSLD1啟動子中脫落酸響應元件可能對表達具有重要影響
由于許多研究者認為纖維細胞的生長發育及其調控機制與植物表皮毛相似[28-29]。因此,多個棉花纖維細胞特異或優勢表達基因啟動子的功能分析都在轉基因煙草和擬南芥中進行。(棉花可逆糖基化多肽基因)啟動子能在轉基因煙草根中表達[30],(棉花葡糖醛基因)可在轉基因煙草種皮、花粉、表皮毛和根冠中表達[31],啟動子能夠指導在轉基因煙草表皮毛中優勢表達[18]。(棉花纖維素合成酶基因)啟動子在煙草表皮毛和葉脈以及下胚軸和莖的維管組織中表達[11]。啟動子在轉基因煙草和擬南芥的表皮毛中特異表達[32]。這些研究說明模式植物煙草和擬南芥分析棉花基因啟動子的功能具有一定的可行性。本研究中,4個不同長度的的啟動子載體都進行了煙草的遺傳轉化并獲得轉基因植株。其中3個較長的啟動子在轉基因煙草中不表達,而最短的啟動子()在轉基因煙草的各個檢測樣品中均表達,而且表達強度與CaMV 35S相似。說明具備啟動子表達的基本元件,但不含有組織器官特異表達元件。在-2 900—-1 232 bp序列中可能存在控制在纖維細胞特異或優勢表達的元件。由于(1 657 bp)無表達,而(1 232 bp)能夠表達,說明-1 657—-1 232 bp的425 bp序列可能具有重要的作用。為此分析了這段序列中的順式調控元件,發現這段序列中包含4個脫落酸響應元件,其中2個元件與油菜素內酯響應元件重疊,以及3個MYB結合位點。有趣的是:全長啟動子序列(2 900 bp)中也只有4個脫落酸響應元件,說明這個啟動子的脫落酸響應元件就集中在這段序列中。3個有這段序列的啟動子在轉基因煙草中無表達,缺失這段序列的啟動子在轉基因煙草中有表達,推測這段序列可能與該啟動子的表達特性有密切關系。下一步可能針對這段序列進行深入研究。(2 178 bp)在轉基因棉花的非纖維部分與煙草相似,也無表達信號;但在纖維細胞中有明顯的表達信號。由此推測:抑制啟動子在非纖維的組織器官中表達的元件可能存在于-2 178—-1 232 bp。下一步研究將對這段啟動子序列進行更加密集的5′-端刪除,可望找到控制基因在纖維細胞中優勢表達的核心序列。
4 結論
(1 232 bp)啟動子在轉基因煙草中是一個廣泛表達啟動子,(2 900 bp)、(2 178 bp)和(1 657 bp)在轉基因煙草中不表達。和之間的差異序列(425 bp)可能對表達特異性有重要影響。是一個棉花纖維細胞優勢表達啟動子,在纖維細胞伸長期(10—15 DPA)表達水平相對較高,總體表達強度低于組成型強啟動子CaMV 35S。該啟動子可應用于棉花纖維發育相關基因的功能研究和改良纖維性狀的分子育種。
[1] JOHN M E, CROW L J. Gene expression in cotton (L.) fiber: cloning of the mRNAs. Proceedings of the National Academy of Sciences of the United States of America, 1992, 89(13): 5769-5773.
[2] JOHN M E, KELLER G. Metabolic pathway engineering in cotton: Biosynthesis of polyhydroxybutyrate in fiber cells. Proceedings of the National Academy of Sciences of the United States of America, 1996, 93(23): 12768-12773.
[3] 于曉紅, 朱勇清, 林芝萍, 陳曉亞, 許智宏. 亞洲棉GAE6-3A上游序列的分離及其在煙草中的表達. 植物生理學報, 2000, 26(2): 143-147, 180.
YU X H, ZHU Y Q, LIN Z P, CHEN X Y, XU Z H. Isolation of GAE6-3A upstream fragment fromand its expression in tobacco. Acta Photophysiologica Sinica, 2000, 26(2): 143-147, 180. (in Chinese)
[4] 吳藹民, 劉進元. 棉花半乳糖苷酶基因啟動子的分離及其在轉基因煙草中的表達. 中國科學C輯: 生命科學, 2005, 35(5): 389-397.
WU A M, LIU J Y. Isolation of cotton galactosidase gene promoter and its expression in transgenic tobacco. Science in China Ser.C Life Sciences, 2005, 35(5): 389-397. (in Chinese)
[5] RINEHART J A, PETERSEN M W, JOHN M E. Tissue-specific and developmental regulation of cotton gene FbL2A (demonstration of promoter activity in transgenic plants). Plant Physiology, 1996, 112(3): 1331-1341.
[6] DELANEY S K, ORFORD S J, MARTIN-HARRIS M, TIMMIS J N. The fiber specificity of the cottongene promoter is regulated by an AT-Rich promoter region and the AT-Hook transcription factor GhAT1. Plant and Cell Physiology, 2007, 48(10): 1426-1437.
[7] MA D P, TAN H, SI Y, CREECH R G, JENKINS J N. Differential expression of a lipid transfer protein gene in cotton fiber. Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism, 1995, 1257(1): 81-84.
[8] MA D P, LIU H C, TAN H, CREECH R G, JENKINS J N, CHANG Y F. Cloning and characterization of a cotton lipid transfer protein gene specifically expressed in fiber cells. Biochimica et Biophysica Acta(BBA)-Lipids and Lipid Metabolism, 1997, 1344(2): 111-114.
[9] LIU H C, CREECH R G, JENKINS J N, MA D P. Cloning and promoter analysis of the cotton lipid transfer protein gene Ltp3(1). Biochimica Et Biophysica Acta(BBA)-Lipids and Lipid Metabolism, 2000, 1487(1): 106-111.
[10] HSU C Y, CREECH R G, JENKINS J N, MA D P. Analysis of promoter activity of cotton lipid transfer protein gene LTP6 in transgenic tobacco plants. Plant Science, 1999, 143(1): 63-70.
[11] WU A M, HU J S, LIU J Y. Functional analysis of a cotton cellulose synthase A4 gene promoter in transgenic tobacco plants. Plant Cell Reports, 2009, 28(10): 1539-1548.
[12] KIM H J, MURAI N, FANG D D, TRIPLETT B A. Functional analysis ofcellulose synthase catalytic subunit 4 promoter in transgenicand cotton tissues. Plant Science, 2011, 180(2): 323-332.
[13] LI X B, FAN X P, WANG X L, CAI L, YANG W C. The cotton ACTIN1 gene is functionally expressed in fibers and participates in fiber elongation. The Plant Cell, 2005, 17(3): 859-875.
[14] DENG F L, TU L L, TAN J F, LI Y, NIE Y C, ZHANG X L. GbPDF1 is involved in cotton fiber initiation via the core cis-element HDZIP2ATATHB2. Plant Physiology, 2011, 158(2): 890-904.
[15] HO M H, SAHA S, JENKINS J N, MA D P. Characterization and promoter analysis of a cotton RING-type ubiquitin ligase (E3) gene. Molecular Biotechnology, 2010, 46(2): 140-148.
[16] HOU L, LIU H, LI J B, YANG X, XIAO Y H, LUO M, SONG S Q, YANG G W, PEI Y. SCFP, a novel fiber-specific promoter in cotton. Chinese Science Bulletin, 2008, 53(17): 2639-2645.
[17] LI Y, TU L L, YE Z Z, WANG M J, GAO W H, ZHANG X L. A cotton fiber-preferential promoter, PGbEXPA2, is regulated by GA and ABA in. Plant Cell Reports, 2015, 34(9): 1539-1549.
[18] 胡海燕, 劉迪秋, 李允靜, 李陽, 涂禮莉. 一個棉花纖維伸長期優勢表達啟動子pGhFLA1的克隆與鑒定. 作物學報, 2017, 43(6): 849-854.
HU H Y, LIU D Q, LI Y J, LI Y, TU L L. identification of promoter GhFLA1 preferentially expressed during cotton fiber elongation. Acta Agronomica Sinica, 2017, 43(6): 849-854. (in Chinese)
[19] WANG L, LIU C, LIU Y J, LUO M. Fumonisin B1-induced changes in cotton fiber elongation revealed by sphingolipidomics and proteomics. Biomolecules, 2020, 10(9): 1258.
[20] WANG Q L, MENG Q, XU F, CHEN Q, MA C X, HUANG L, LI G M, LUO M. Comparative metabolomics analysis reveals sterols and sphingolipids play a role in cotton fiber cell initiation. International Journal of Molecular Sciences, 2021, 22(21): 11438.
[21] CHEN Q, XU F, WANG L, SUO X D, WANG Q L, MENG Q, HUANG L, MA C X, LI G M, LUO M. Sphingolipid profile during cotton fiber growth revealed that a phytoceramide containing hydroxylated and saturated VLCFA is important for fiber cell elongation. Biomolecules, 2021, 11(9): 1352.
[22] LUO M, XIAO Y, LI X, LU X, DENG W, LI D, HOU L, HU M, LI Y, PEI Y. GhDET2, a steroid 5α-reductase, plays an important role in cotton fiber cell initiation and elongation. The Plant Journal, 2007, 51(3): 419-430.
[23] WANG M J, TU L L, LIN M, LIN Z X, WANG P C, YANG Q Y, YE Z X, SHEN C, LI J Y, ZHANG L, ZHOU X L, NIE X H, LI Z H, GUO K, MA Y Z, HUANG C, JIN S X, ZHU L F, YANG X Y, MIN L, YUAN D J, ZHANG Q H, LINDSEY K, ZHANG X. Asymmetric subgenome selection and cis-regulatory divergence during cotton domestication. Nature Genetics, 2017, 49(4): 579-587.
[24] ZHANG J X, WANG F R, ZHANG C Y, ZHANG J H, CHEN Y, LIU G D, ZHAO Y X, HAO F S, ZHANG J. A novel VIGS method by agroinoculation of cotton seeds and application for elucidating functions of GhBI-1 in salt-stress response. Plant Cell Reports, 2018, 37(8): 1091-1100.
[25] ZHANG J, HUANG G Q, ZOU D, YAN J Q, LI Y, HU S, LI X B. The cotton () NAC transcription factor (FSN1) as a positive regulator participates in controlling secondary cell wall biosynthesis and modification of fibers. The New Phytologist, 2018, 217(2): 625-640.
[26] JIANG Y J, GUO W Z, ZHU H Y, RUAN Y L, ZHANG T Z. Overexpression of GhSusA1 increases plant biomass and improves cotton fiber yield and quality. Plant Biotechnology Journal, 2012, 10(3): 301-312.
[27] ZHANG M, ZHENG X L, SONG S Q, ZENG Q W, HOU L, LI D M, ZHAO J, WEI Y, LI X B, LUO M, XIAO Y H, LUO X Y, ZHANG J F, XIANG C B, PEI Y. Spatiotemporal manipulation of auxin biosynthesis in cotton ovule epidermal cells enhances fiber yield and quality. Natture Biotechnology, 2011, 29(5): 453-458.
[28] GUAN X Y, YU N, SHANGGUAN X X, WANG S, LU S, WANG L J, CHEN X Y.trichome research sheds light on cotton fiber development mechanisms. Chinese Science Bulletin, 2007, 52(13): 1734-1741.
[29] WANG Z, YANG Z R, LI F G. Updates on molecular mechanisms in the development of branched trichome inand nonbranched in cotton. Plant Biotechnology Journal, 2019, 17(9): 1706-1722.
[30] WU A M, LING C, LIU J Y. Isolation of a cotton reversibly glycosylated polypeptide (GhRGP1) promoter and its expression activity in transgenic tobacco. Journal of Plant Physiology, 2006, 163(4): 426-435.
[31] WU A M, LV S Y, LIU J Y. Functional analysis of a cotton glucuronosyltransferase promoter in transgenic tobaccos. Cell Research, 2007, 17(2): 174-183.
[32] SHANGGUAN X X, XU B, YU Z X, WANG L J, CHEN X Y. Promoter of a cotton fibre MYB gene functional in trichomes ofand glandular trichomes of tobacco. Journal of Experimental Botany, 2008, 59(13): 3533-3542.
Cloning and functional characterization of the promoter ofgene That predominantlyexpressed in cotton fiber
LIU Fang, XU Mengbei, WANG Qiaoling, MENG Qian, LI Guiming, ZHANG Hongju, TIAN Huidan, XU Fan, LUO Ming
Biotechnology Research Center, Southwest University/Key Laboratory of Biotechnology and Crop Quality Improvement, Ministry of Agriculture and Rural Affairs, Chongqing 400716
【Objective】Cotton fiber is the main economic product of cotton. It is the epidermal cells of the ovule outer integument through polar elongation and secondary wall thickening. As one of the longest plant cells, the cotton fiber cells are regarded as an ideal material in the study of plant cell growth and development. Identification of promoters specifically or preferentially expressed in fiber cells is of great significance for basic research on fiber development and molecular breeding for improving fiber traits. 【Method】In this study, we cloned the promoter ofgene, which is predominantly expressed in fiber cells. Through the PlantCARE website for promoter sequence analysis, we identified the important-regulatory elements contained in the cloned sequence. According to the distribution of some important-regulatory elements, the cloned promoter fragments were deleted at 5′- end. A total of 4 promoter fragments were obtained and the corresponding plant expression vector was constructed. The constructed plant expression vectors were used for genetic transformation of tobacco and cotton. The transgenic plants were identified through molecular identification of transgenic tobacco and cotton. GUS activity in different tissues, organs and fiber cells of transgenic plants at different development stages was also investigated. 【Result】The longest promoter cloned was 2 900 bp in length. In addition to a lot of transcription regulatory elements in the promoter, the sequence also contained multiple abscisic acid response elements, the elements essential for the anaerobic induction, methyl jasmonate response elements, brassinolide response elements, the elements involved in seed-specific regulation, the elements involved in defense and stress responsiveness, and MYB transcription factor binding sites. Four promoter fragments with a length of 2 900 bp (), 2 178 bp (), 1 657 bp () and 1 232 bp () were obtained by the 5′-terminal deletion, respectively. The transgenic tobacco plants were generated after confirmed by molecular identification.,anddid not express in transgenic tobacco, whileis widely expressed, and the expression level ofwas similar to that of CaMV 35S promoter. The different sequence betweenandcontained four abscisic acid response elements, two brassinolide response elements, and three MYB binding sites. These-regulatory elements may be associated with the non-expression of,, andpromoters in transgenic tobacco. The transgenic cotton plants ofwere obtained after confirmed by molecular identification.predominantly expressed in transgenic cotton fibers, and its expression level was higher at the elongation stage (10-15 DPA) of fiber cells while lower in the early developmental stage (5 DPA) of fiber cells and the stage of secondary cell wall deposition (20-30 DPA). 【Conclusion】Thepromoter was a widely expressed promoter, and thepromoter was a fiber predominant expression promoter, which was highly expressed during the elongation of fibers. It could be applied to the study on the gene function involved in cotton fiber development and molecular breeding for improving fiber traits.
cotton; promoter; functional characterization;; sphingolipid delta8-desaturase
2022-11-07;
2022-12-13
國家自然科學基金(31971984,31571722)
劉芳,Tel:15330430736;E-mail:lf530805@163.com。通信作者羅明,Tel:13996007163;E-mail:luo0424@126.com
10.3864/j.issn.0578-1752.2023.19.002
(責任編輯 李莉)
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
