豬ECR1-like免疫黏附功能對PAMs捕獲GFP-E. coli的影響
2023-10-25 10:24:00張崢凌小雅范闊海孫娜孫盼盼孫耀貴李宏全尹偉
中國農業科學 2023年19期
關鍵詞:檢測
張崢,凌小雅,范闊海,孫娜,孫盼盼,孫耀貴,李宏全,尹偉
豬ECR1-like免疫黏附功能對PAMs捕獲GFP-的影響
1中獸醫藥現代化山西省重點實驗室/山西農業大學動物醫學學院,山西太谷 030801;2中獸醫藥現代化山西省重點實驗室/山西農業大學動物實驗中心,山西太谷 030801
【目的】探討豬紅細胞類Ⅰ型補體受體(erythrocyte complement receptor type 1-like, ECR1-like)免疫黏附功能是否能夠促進豬肺泡巨噬細胞(porcine alveolar macrophages, PAMs)捕獲致敏基因工程菌(GFP-, GFP),以期闡釋豬紅細胞免疫的分子機理及紅細胞免疫在機體先天免疫中的作用。【方法】利用流式細胞術、菌落平板計數及RT-PCR技術檢測PAMs捕獲的GFP-的水平,分析豬ECR1-like免疫黏附對PAMs捕獲GFP-的影響;運用流式細胞術、細胞免疫熒光化學技術檢測豬ECR1-like免疫黏附的致敏GFP-被PAMs移除后,豬紅細胞免疫黏附功能的變化及豬ECR1-like數量的變化。【結果】流式細胞術檢測發現豬紅細胞黏附組較空白對照組中的PAMs平均熒光強度顯著提高(<0.001),且PAMs陽性細胞率也顯著提高(<0.05);菌落涂板計數發現紅細胞黏附組較空白對照組PAMs捕獲GFP-情況顯著增強(<0.05);RT-PCR檢測發現,紅細胞黏附組PAMs中GFP-的相對數量顯著高于空白對照組(<0.01);進一步阻斷豬紅細胞表面的CR1-like,流式細胞術檢測發現PAMs平均熒光強度降低至256 301.56±9 208.85(<0.001),PAMs陽性細胞率降低至(88.32±0.92)%(>0.05),菌落平板計數發現PAMs捕獲GFP-情況減弱為(136 666±8 818)CFU/mL(<0.05),RT-PCR檢測發現PAMs中GFP-的相對數量顯著減少(<0.01);利用細胞流動循環互作技術發現:豬紅細胞免疫黏附致敏GFP-的平均熒光強度由循環前2 892.18±47.76降至2 407.43±141.78(<0.05),陽性細胞率由循環前(20.58±0.36)%降至(17.39±0.23)%(<0.001),黏附水平顯著低于循環前,與此同時,間接免疫熒光試驗結果顯示:試驗組循環后豬ECR1-like平均熒光強度由循環前344.33±37.92降低至291.56±11.99(<0.05),陽性細胞率由(30.20±1.24)%減少至(28.27±0.64)%(<0.05)。【結論】豬ECR1-like通過免疫黏附功能促進了PAMs對致敏GFP-的捕獲。PAMs移除豬紅細胞表面黏附的致敏GFP-后,豬紅細胞的活性CR1-like減少,免疫黏附功能下降。
豬;紅細胞;豬肺泡巨噬細胞;類Ⅰ型補體受體;免疫黏附
0 引言
【研究意義】自1981年SIEGEL等首次提出“紅細胞免疫系統”概念,并發現紅細胞具有多種免疫功能[1]。紅細胞表面存在I型補體受體(erythrocyte complement receptor type 1, ECR1),且在ECR1的介導下,能夠黏附經補體活性成分C3b致敏的病原體或免疫復合物(immune complex, IC),通過血液循環至肝臟、脾臟,最終由吞噬細胞清除[2]。紅細胞類I型補體受體(erythrocyte complement receptor I-like,ECR1-like)是豬紅細胞膜表面的天然免疫活性分子,其被證實是C3b調理免疫復合物的免疫黏附受體[3-5]。研究表明,豬紅細胞能夠通過CR1-like介導的免疫黏附功能,從而結合經血清致敏的大腸桿菌[6-7]。ECR1-like在清除IC的過程中同樣發揮著重要作用,因此,深入探究ECR1-like介導清除IC的過程及分子機制對于維持機體內環境穩態、免疫平衡及機體健康具有重要意義。【前人研究進展】紅細胞CR1的免疫黏附功能在相關疾病發生發展及轉歸中發揮重要作用。研究表明,紅細胞免疫黏附功能與細菌[8-10]、病毒[11-13]、寄生蟲[14]等病原體的清除具有相關性,且紅細胞表面的CR1分子在機體清除內源性IC的過程中同樣發揮著重要作用[15-18]。REINAGEL等[19]將補體致敏雜聚物(heterochain polymer, HP)和噬菌體φX174作為模式IC進行研究,發現人紅細胞結合IC后與小鼠巨噬細胞孵育,在小鼠巨噬細胞內能檢測到IC的存在,表明紅細胞結合的IC可由小鼠巨噬細胞吞噬內化,同時伴隨有紅細胞CR1的損失,通過ELISA檢測到CR1出現在巨噬細胞內部,CR1可能同IC一起被巨噬細胞吞噬內化。HEPBURN等[20]建立了單層巨噬細胞的平行板流動系統用于檢測體內血流剪切力條件下IC從紅細胞表面的轉移,Fc受體(Fc receptor, FcR)和補體受體(complement receptor, CR)的阻斷抑制了IC向巨噬細胞的轉移,其中,FcγRIIa的阻斷影響最大,FcR和CR在巨噬細胞移除紅細胞表面IC的過程中發揮協同作用,共阻斷抑制作用最明顯。本課題組前期經研究發現豬紅細胞上存在CR1-like[3]。且證明其是通過EPB41分子錨合在紅細胞膜表面[21-22],在體外條件下,經酵母雜交體系,研究證明了豬紅細胞CR1-like活性片段能夠與C3b相互結合[5]。PAMs能通過CR1-like競爭性結合豬紅細胞表面黏附的GFP-,且豬紅細胞形態不受影響。但是,豬ECR1-like水平顯著降低[23-24]。【本研究切入點】本試驗通過模擬血液循環的自然生理狀態,構建豬紅細胞與PAMs動態互作循環系統,針對PAMs捕獲GFP-的量和循環前后的平均熒光強度和陽性細胞率進行分析。【擬解決的關鍵問題】探討ECR1-like介導PAMs移除表面GFP-過程中PAMs捕獲效率是否提高?豬紅細胞免疫黏附水平是否變化?等關鍵問題,深入探究紅細胞CR1清除IC的過程及分子機制,以期為調控紅細胞免疫應用于動物臨床研究奠定理論基礎。
1 材料與方法
1.1 試驗動物及菌株
試驗于2020年10月至2022年5月進行。試驗地點為山西農業大學中獸醫藥現代化山西省重點實驗室。試驗所需健康40日齡長白仔豬,體重(20±2)kg,購自太谷縣冠農農牧科技有限公司。表達綠色熒光蛋白的大腸桿菌(GFP-)由山西農業大學中獸醫藥現代化山西省重點實驗室制備保存。
1.2 試驗試劑
鼠抗豬CR1-like McAb(IgG1亞型)為山西農業大學中獸醫藥現代化山西省重點實驗室制備(專利:ZL201410308534.0);Mouse IgG1 Isotype Control購自北京博奧森生物技術有限公司;羊抗小鼠IgG- CoraLite 488購自武漢三鷹生物技術有限公司。
RPMI-1640培養基,購自美國Gibico公司;DMSO、青鏈霉素混合液、多聚-L-賴氨酸(15—30萬)、Hank’s緩沖液,均購自索萊寶科技有限公司;胎牛血清,購自以色列Biolnd公司;胰蛋白胨、酵母提取物,購自上海生工生物工程有限公司;豬外周血紅細胞分離液試劑盒購自天津市灝洋生物制品科技有限責任公司;TaKaRa PrimeScriptTM RT reagent Kit with gDNA Eraser、Trizol、SYBR Premix Ex Taq TM II均購自寶日醫生物技術(北京)有限公司。
1.3 豬ECR1-like活性影響PAMs捕獲GFP-E.coli的檢測
1.3.1 流式細胞術檢測PAMs陽性細胞率及平均熒光強度 參照文獻[25-27]的方法制備豬紅細胞懸液、豬血清和致敏GFP-懸液,構建豬紅細胞與PAMs動態互作體系按文獻[28]方法操作。分4組進行檢測,即空白對照組、紅細胞黏附組、ECR1-like阻斷組和同型對照組,分別記為A、B、C、D組。
A組:取4 mL致敏GFP-懸液與4 mL Hank’s緩沖液直接孵育,結束后加入8 mL Hank’s緩沖液,將致敏GFP-懸液置于流動相瓶中,按照文獻[27]方法使菌液循環流動,待充滿整個小室后開始計時,60 min后進行后續檢測;B組:取Hank’s緩沖液80 μL與4 mL豬紅細胞懸液混勻于37 ℃孵育2 h,孵育結束后1 500 r/min,5 min離心兩次,后續方法同上;C組:取豬CR1-like McAb 80 μL與4 mL豬紅細胞懸液混勻于37 ℃孵育2 h,孵育結束后1 500 r/min,5 min離心兩次,后續方法同上;D組:以IgG1同型抗體替代C組的豬CR1-like McAb,其余操作同C組。
取上述各組循環后的PAMs載玻片,流式細胞儀檢測各組PAMs陽性細胞率及平均熒光強度。
1.3.2 菌落平板計數法檢測PAMs裂解液中GFP-數量 取上述4組循環后的PAMs載玻片,PBS洗滌3次,加1 mL 0.3%的TritonX-100裂解10 min。收集PAMs及裂解產物,稀釋100倍后均勻涂布15 μL于LB固體培養基(含100 μg·mL-1Amp),37℃倒置培養16 h,計數各組GFP-的CFU。按式(1)分別計算各組PAMs捕獲GFP-的情況。
PAMs吞噬情況(CFU/mL)=

1.3.3 RT-PCR檢測PAMs中GFPmRNA相對表達量 取上述各組循環后的PAMs載玻片,PBS洗滌3次,加入1 mL Trizol,提取RNA,經TaKaRa SYBR PrimeScript TM RT reagent Kit with gDNA Eraser試劑盒反轉錄為cDNA,引物的設計參考NCBI數據庫進行設計(表1),以GAPDH作內參,實時熒光定量PCR檢測GFP-基因的變化。

表1 引物序列
1.4 PAMs移除致敏GFP-E.coli后豬紅細胞免疫黏附功能的檢測
1.4.1 流式細胞術檢測豬紅細胞免疫黏附GFP-水平 分3組進行檢測,即空白對照組、試驗組、陰性對照組,分別記為Ⅰ、Ⅱ、Ⅲ 3組。
(Ⅰ)空白對照組:取未復蘇PAMs的空白潔凈載玻片,蓋上平行板流動小室,以黏附有致敏GFP-的豬紅細胞懸液16 mL為流動相,建立循環流動體系。啟動蠕動泵,待流動相充滿整個平行板流動小室后開始計時,60 min進行后續檢測;(Ⅱ)試驗組:取復蘇有PAMs的載玻片,蓋上平行板流動小室,以黏附有致敏GFP-的豬紅細胞懸液16 mL為流動相,其余操作同(Ⅰ)組;(Ⅲ)陰性對照組:取復蘇有PAMs的載玻片,蓋上平行板流動小室,以豬紅細胞懸液16 mL為流動相,其余操作同(Ⅰ)組。
取各組0 min時的流動相紅細胞懸液300 μL,取各組60 min時的流動相紅細胞,按照1.3的方法再次黏附致敏GFP-,用流式細胞儀檢測豬紅細胞平均熒光強度及陽性細胞率,兩次樣的數值分別代表PAMs移除致敏GFP-前后豬紅細胞的免疫黏附功能。
1.4.2 流式細胞術檢測豬ECR1-like數量 試驗分為6組,分別記為a、b、c、d、e、f組。
a組:取循環前的豬紅細胞懸液200 μL,1 500 r/min離心5 min,棄上清,加入0.5% BSA 200 μL重懸,于37℃孵育30 min,1 500 r/min、5 min離心洗滌2次,以200 μL Hank’s重懸。按1﹕50比例加入豬CR1-like McAb,37℃孵育1 h后,用Hank’s緩沖液1 500 r/min、5 min離心洗滌2次,以200 μL Hank’s重懸。再向其中加入2 μL羊抗小鼠IgG-CoraLite 488,37 ℃避光孵育1 h,間歇振蕩。孵育完畢,用Hank’s緩沖液1 500 r/min、5 min離心洗滌2次,重懸后用流式細胞儀檢測其平均熒光強度。
b組:取1.4.1中I組60 min后的豬紅細胞懸液200 μL,其余操作同a組。
c組:取1.4.1中Ⅱ組60 min后的豬紅細胞懸液200 μL,其余操作同a組。
d組:取1.4.1中III組60 min后的豬紅細胞懸液200 μL,其余操作同a組。
e組:取循環前的豬紅細胞懸液200 μL,以小鼠同型抗體IgG1代替CR1-like McAb進行處理,其余操作同a組。
f組:取循環前的豬紅細胞懸液200 μL,Hank’s緩沖液代替抗體進行處理,其余操作同a組。
2 結果
2.1 豬ECR1-like增強PAMs捕獲GFP-E.coli后的平均熒光強度及陽性細胞率。
收集各組循環后的PAMs,用流式細胞儀分析PAMs陽性細胞率及平均熒光強度并進行統計(圖1,表2)。
各組平均熒光強度經單因素方差分析可見:與A組相比,B組PAMs平均熒光強度極顯著增高(<0.001),C組PAMs平均熒光強度較B組極顯著降低(<0.001),D組PAMs平均熒光強度與B組相比無顯著差異(>0.05);各組陽性細胞率經單因素方差分析可見:與A組相比,B組PAMs陽性細胞率顯著增高(<0.05),C、D組PAMs陽性細胞率與B組相比均無顯著差異(>0.05,圖2)。
2.2 豬ECR1-like增強PAMs捕獲GFP-E. coli的數量
收集各組循環后的PAMs載玻片,收集PAMs并裂解,涂布于LB固體培養基(含Amp),根據培養結果計算得出各組PAMs捕獲致敏GFP-的情況(圖3,表3)。將計數結果進行單因素方差分析可見:與A組相比,B組PAMs在1 h內對GFP-的捕獲數量顯著增多(<0.05),C組PAMs捕獲GFP-較B組顯著減少(<0.05),D組PAMs捕獲GFP-數與B組相比無顯著差異(>0.05,圖4)。
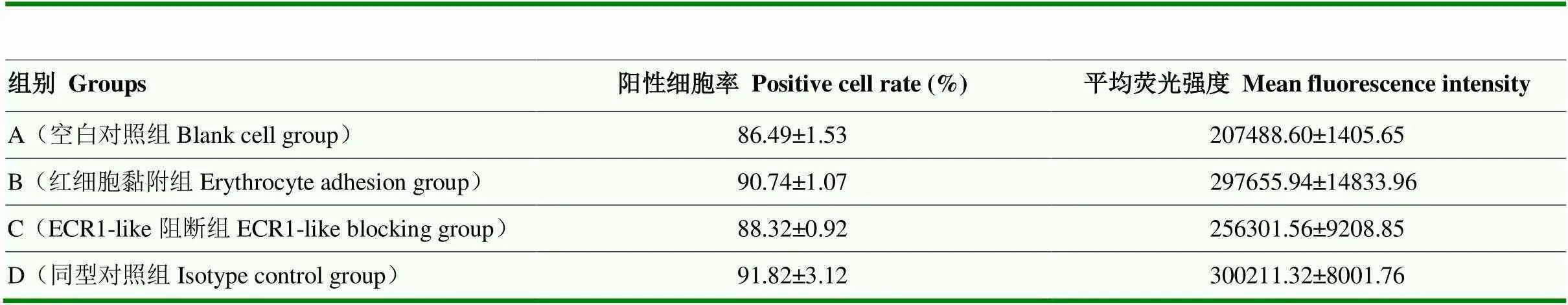
表2 阻斷豬ECR1-like活性對PAMs陽性細胞率及平均熒光強度的影響
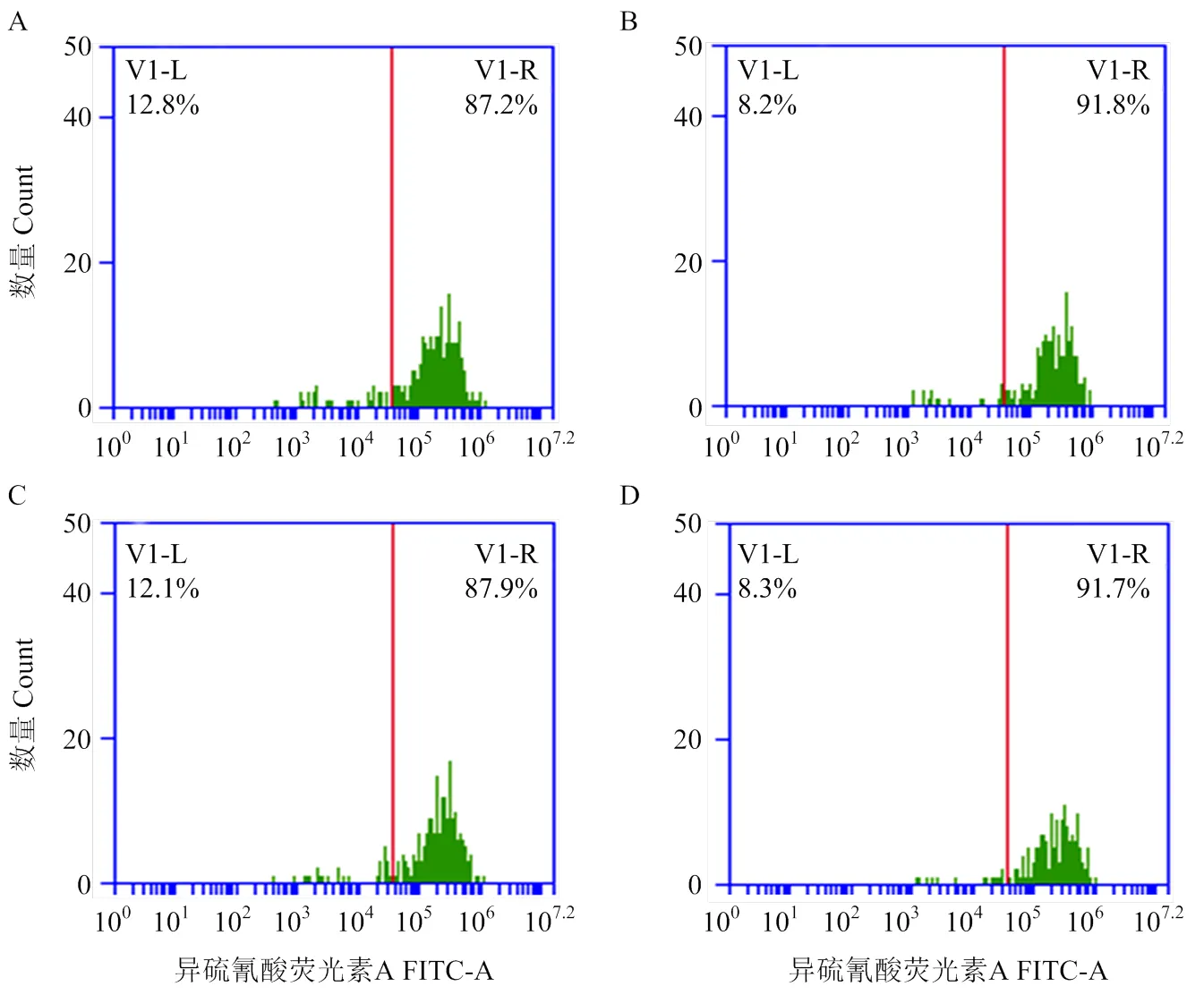
A為空白對照組;B為紅細胞黏附組;C為ECR1-like阻斷組;D為同型對照組。圖2—5同

圖2 PAMs陽性細胞率及平均熒光強度單因素方差分析

圖3 阻斷豬ECR1-like活性后PAMs捕獲GFP-E. coli的平板涂布情況

表3 阻斷豬ECR1-like活性對PAMs捕獲GFP-E. coli菌落計數結果的影響
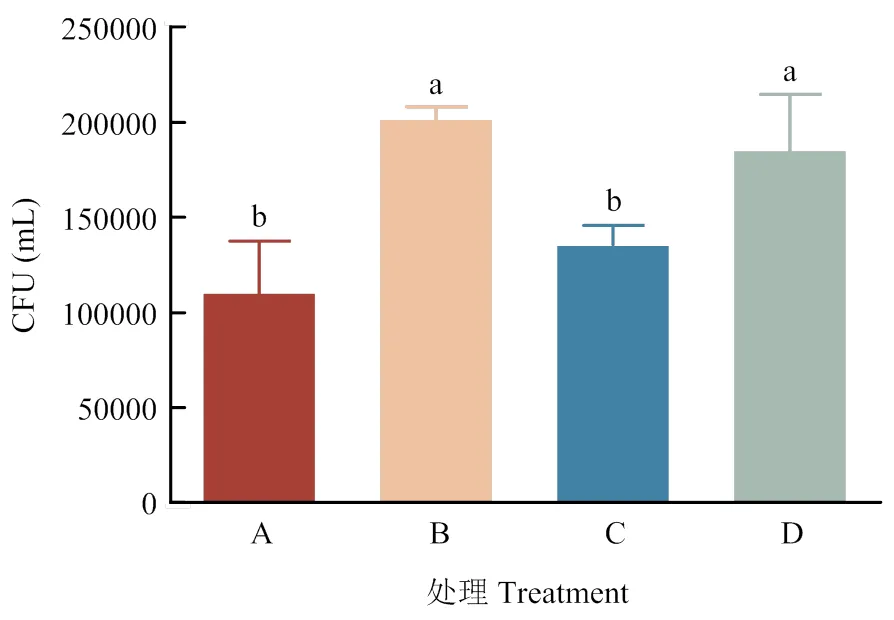
圖4 PAMs捕獲GFP-E. coli情況單因素方差分析
2.3 豬ECR1-like增強PAMs中GFP-E coliβ-D- galactosidase mRNA相對表達量
RT-PCR檢測大腸桿菌中mRNA相對表達量,從而反映GFP-的相對數量,進行單因素方差分析。與A組相比,B組PAMs中GFP-的相對數量極顯著升高(<0.01),與B組相比,C組PAMs中GFP-的相對數量極顯著降低(<0.01),D組中GFP-的相對數量無顯著變化(>0.05,圖5)。

圖5 PAMs捕獲GFP-E coli相對數量單因素方差分析
2.4 PAMs移除致敏GFP-E. coli后豬紅細胞免疫黏附功能下降
流式細胞技術檢測各組循環前后豬紅細胞的免疫黏附GFP-的功能(圖6),以豬紅細胞循環前免疫黏附及循環后重新黏附GFP-的陽性細胞率和平均熒光強度作為評判(表4)。
將平均熒光強度數據進行T檢驗分析,Ⅱ組PAMs移除豬紅細胞表面致敏GFP-后免疫黏附平均熒光強度較循環前免疫黏附平均熒光值顯著降低(<0.05);Ⅰ、Ⅲ組循環前、后豬紅細胞免疫黏附平均熒光值無顯著變化(>0.05,圖7)。
將陽性細胞率數據進行T檢驗分析,Ⅱ組PAMs移除豬紅細胞表面致敏GFP-后豬紅細胞免疫黏附陽性細胞率較循環前豬紅細胞的免疫黏附陽性細胞率極顯著降低(<0.001);Ⅰ、Ⅲ組循環前、后豬紅細胞免疫黏附陽性細胞率無顯著變化(>0.05,圖8)。

表4 PAMs移除致敏GFP-E. coli前后豬紅細胞免疫黏附陽性細胞率及平均熒光強度檢測

Ⅰ為空白對照組,Ⅱ為試驗組,Ⅲ為陰性對照組。圖7和8同
2.5 PAMs移除致敏GFP-E. coli后導致豬ECR1-like數量減少
細胞免疫熒光結合流式細胞術對Ⅰ、Ⅱ、Ⅲ組處理后的豬ECR1-like的數量變化進行檢測(圖9),以豬紅細胞陽性細胞率及平均熒光強度作為評判(表5),進行單因素方差分析(圖10)。
b組與a組平均熒光強度相比無顯著差異(>0.05),c組較a組平均熒光強度相比顯著降低(<0.05),d組與a組相比無顯著差異(>0.05);e組極顯著低于a、b、c、d組的平均熒光強度(<0.01),f組極顯著低于a、b、c、d、e組的平均熒光強度(<0.01)。
b組與a組陽性細胞率相比無顯著差異(>0.05),c組較a組陽性細胞率相比顯著降低(<0.05),d組與a組陽性細胞率相比無顯著差異(>0.05);e組極顯著低于a、b、c、d組的陽性細胞率(<0.01),f組為空白對照,無陽性細胞。

圖7 PAMs移除致敏GFP-E. coli前后豬紅細胞免疫黏附水平(平均熒光強度)的比較

圖8 PAMs移除致敏GFP-E. coli前后豬紅細胞免疫黏附水平(陽性細胞率)的比較

表5 PAMs移除致敏GFP-E. coli前后豬ECR1-like陽性細胞率及平均熒光強度檢測
3 討論
3.1 豬ECR1-like促進PAMs捕獲GFP-E. coli
本試驗以致敏GFP-作為模式IC觀察豬ECR1-like免疫黏附在PAMs清除IC過程中的作用,發現豬ECR1-like免疫黏附能夠促進PAMs對致敏GFP-的捕獲。試驗運用3種不同技術對PAMs捕獲GFP-的量進行了檢測,首先,運用流式細胞術檢測了PAMs的平均熒光強度和陽性細胞率,平均熒光強度越高,表明PAMs捕獲的GFP-越多,結果顯示,在紅細胞介入的試驗組中PAMs平均熒光強度顯著高于沒有紅細胞介入的等量的單純的GFP-空白對照組,表明紅細胞在該體系中能夠促進PAMs對GFP-的捕獲,進而預先以CR1-like單抗阻斷紅細胞表面的CR1-like位點,再將處理過的紅細胞以上述方法操作試驗,豬紅細胞對PAMs捕獲GFP-的促進作用減弱,與試驗組相比PAMs平均熒光強度顯著降低,這表明豬紅細胞能夠促進PAMs對GFP-的清除,發揮功能的分子基礎與CR1-like相關。收集各組試驗后的PAMs裂解后進行菌落涂板計數能夠直觀地評估PAMs捕獲GFP-的情況,將計數結果進行統計分析,試驗組比空白對照組細菌數顯著增多,ECR1-like阻斷組比試驗組細菌數顯著減少,與空白對照組細菌數無差異,這樣的趨勢與流式細胞術檢測結果一致,并展現了ECR1-like在PAMs清除GFP-過程中的關鍵作用。之后,運用RT-PCR技術對PAMs捕獲GFP-的情況進行檢測,統計分析得到了和前兩種技術結果相同的趨勢。而且,ECR1-like的阻斷組除了阻斷對照作用,還能夠作為無免疫黏附功能的粒子介入的陰性對照,即使豬紅細胞喪失了免疫黏附功能,又能保持和豬紅細胞相同的細胞特性,試驗對照更加完善,更加提高了試驗結果的客觀性與準確性。前期試驗結果表明,CR1-like是豬紅細胞免疫黏附重要的分子基礎,CR1-like單抗對豬紅細胞免疫黏附具有阻斷作用[29]。結合本試驗3種技術得到的結果分析得出:豬紅細胞免疫黏附能夠促進PAMs對致敏GFP-的吞噬清除,CR1-like是發揮促進功能的分子基礎。也就是說,豬ECR1-like免疫黏附在吞噬細胞清除IC的過程中起到了正面的、積極的促進作用。
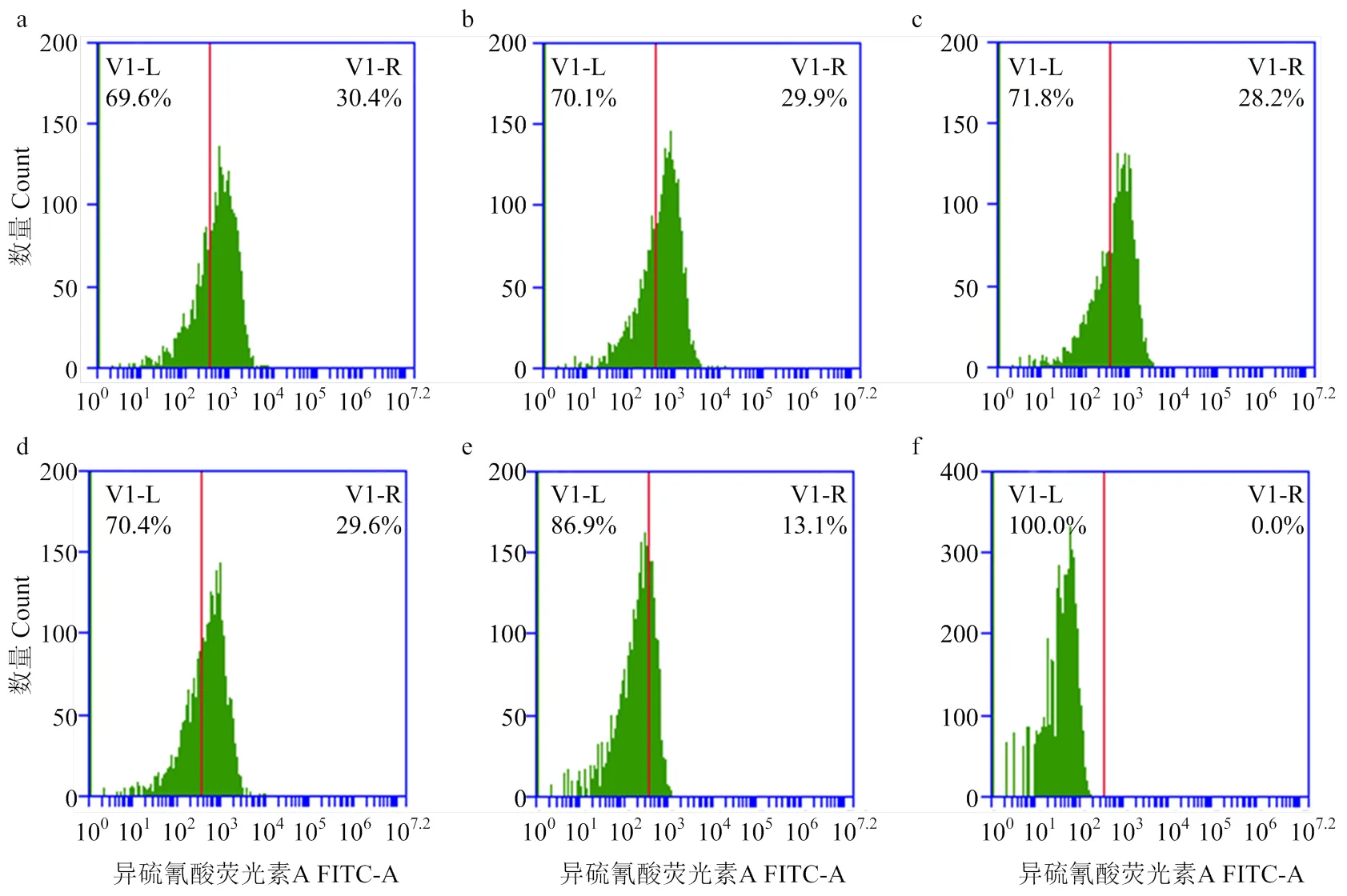
a組為正常對照組,b組為Ⅰ處理組,c組為Ⅱ處理組,d組為Ⅲ處理組,e組為同型對照組,f組為空白細胞組,圖10同

圖10 PAMs移除致敏GFP-E. coli后豬ECR1-like數量的比較
這與LI等的研究結果一致,LI等[9, 29]以CR1+轉基因小鼠紅細胞和野生型小鼠紅細胞為研究對象對小鼠清除肺炎鏈球菌的效率進行研究,將等量的FITC標記的肺炎鏈球菌與紅細胞孵育,并加入小鼠巨噬細胞系J774A.1中以評估肺炎鏈球菌向巨噬細胞的轉移,結果發現,30 min后,與野生型小鼠紅細胞相比,從表達CR1的小鼠紅細胞轉移至巨噬細胞的肺炎鏈球菌顯著增多。然而2019年BREKKE等[8]的研究發現,紅細胞CR1表達量越高,結合大腸桿菌數越多,人全血中白細胞(包括粒細胞和單核細胞)的吞噬能力下降;在將CR1阻斷后,血漿游離菌的數量增多,白細胞的吞噬能力增強,推測其原因可能是因為大腸桿菌的游離增加了白細胞對其的可得到性。本試驗結果與此結果的差異可能因不同病原體而異,也可能是由于在吞噬系統中單核細胞和巨噬細胞等不同吞噬細胞的角色和職能有所差異。
由于1 h的時間限制,觀測到的豬紅細胞能夠促進PAMs對紅細胞表面致敏的GFP-的捕獲結果可以解釋為兩種情況,一種是豬紅細胞的介入能夠加快肺泡巨噬細胞對GFP-的獲取,在單位時間內PAMs對IC的捕獲增多;經過對PAMs陽性細胞率的統計比較,也可能是紅細胞的介入增加了活性PAMs的數量,使得之前不能及時捕獲IC的PAMs能夠快速結合IC。另外,從顯微鏡記錄的過程來看,PAMs在1 h內便可將部分GFP-吞噬清除,因此,試驗條件需進一步優化以得到更加準確的結果。豬紅細胞表面CR1-like被阻斷后,對PAMs捕獲IC的促進功能未完全消失,可能在PAMs清除豬紅細胞表面IC的過程中除了CR1-like還有其他分子發揮作用。前期實驗室的研究也證明了PAMs上存在有CR1-like,且PAMs表面CR1-like相對數量是豬紅細胞的4—5倍甚至更多[26],IC從紅細胞到巨噬細胞上的轉移是PAMs上FcR和CR1-like共同作用的結果,兩個受體可產生協同作用[24]。從本試驗的結果中也不難看出,PAMs對GFP-的黏附及吞噬功能強于豬紅細胞,CR1-like相對數量越多,免疫黏附功能越強,IC的轉移是CR1-like多的細胞從CR1-like少的細胞的競爭性結合,無論是阻斷PAMs表面的CR1-like或是阻斷豬紅細胞表面的CR1-like,PAMs移除GFP-的量都會減少,這表明CR1-like在其中起到了重要的承接作用,而且,CR1-like與CR1-like獨特承接介導的IC轉移可能更快。
3.2 PAMs移除致敏GFP-E. coli后豬紅細胞免疫黏附功能下降
本試驗利用流式細胞技術結合免疫熒光細胞化學技術初步探究了PAMs清除豬紅細胞表面IC后豬紅細胞免疫黏附功能的變化,將流動小室循環后的豬紅細胞再次進行免疫黏附致敏GFP-,用流式細胞儀檢測其熒光強度,與循環前豬紅細胞免疫黏附致敏GFP-的熒光強度進行T檢驗得出,PAMs清除豬紅細胞表面GFP-后,豬紅細胞免疫黏附水平下降,而CR1-like是豬紅細胞發揮免疫黏附功能重要的分子基礎,其數量也呈下降趨勢。試驗設置了兩個對照組,空白對照組為無PAMs組,循環體系中只有免疫黏附致敏GFP-的豬紅細胞,該組的設立避免了平行板流動小室物理循環體系對試驗結果的干擾;陰性對照組為PAMs與豬紅細胞循環組,其中的紅細胞沒有黏附致敏GFP-,結果證明自然情況下正常豬紅細胞不會與PAMs互作而導致豬紅細胞免疫黏附水平及CR1-like的變化。結合以上分析說明,PAMs移除豬紅細胞表面GFP-這一生物學過程導致了豬紅細胞免疫黏附水平的下降和ECR1-like的減少。這與前人研究結果一致[30-32]。但是由于本試驗CR1-like McAb的限制,并不能完全下結論為豬紅細胞表面CR1-like在此過程中丟失,也可能由于IC清除過程中對CR1-like位點的占用或CR1-like變構所致,其具體機制還待進一步試驗探究。
4 結論
在體外流動狀態下,豬紅細胞類Ⅰ型補體受體通過其免疫黏附功能能夠促進豬肺泡巨噬細胞對致敏GFP-的捕獲。豬肺泡巨噬細胞移除豬紅細胞表面黏附的致敏GFP-后,豬紅細胞的活性CR1-like減少,免疫黏附功能下降。
[1] SIEGEL I, TIAN L L, GLEICHER N. The red-cell immune system. The Lancet, 1981, 318(8246): 556-559.
[2] JAVA A, LISZEWSKI M K, HOURCADE D E, ZHANG F, ATKINSON J P. Role of complement receptor 1 (CR1; CD35) on epithelial cells: a model for understanding complement-mediated damage in the kidney. Molecular Immunology, 2015, 67(2 Pt B): 584-595.
[3] YIN W, CUI J Y, JIANG J B, ZHAO J X, FAN K H, SUN N, WANG Z W, SUN Y G, MA H L, LI H Q. The immune adherence receptor CR1-like existed on porcine erythrocytes membrane. Scientific Reports, 2015, 5: 13290.
[4] HOU Z, YIN W, HAO Z L, FAN K H, SUN N, SUN P P, LI H Q. Molecular simulation study on the interaction between porcine CR1-like and C3b. Molecules, 2023, 28(5): 2183.
[5] 孫雨晨, 賈瑞璞, 范闊海, 孫娜, 孫耀貴, 孫盼盼, 李宏全, 尹偉. 豬Ⅰ型補體受體與C3b活性片段相互結合的體外檢測. 中國農業科學, 2021, 54(19): 4243-4254.
SUN Y C, JIA R P, FAN K H, SUN N, SUN Y G, SUN P P, LI H Q, YIN W. Detection of interaction between porcine type Ⅰ complement receptor and C3b active fragment. Scientia Agricultura Sinica, 2021, 54(19): 4243-4254. (in Chinese)
[6] SUN Y G, YIN W, FAN X F, FAN K H, JIANG J B, LI H Q. The cytological observation of immune adherence of porcine erythrocyte. Cell Communication & Adhesion, 2012, 19(5/6): 79-84.
[7] YIN W, WANG C, FAN K H, SUN N, SUN Y G, LI H Q.observation: The GFP-.adhering to porcine erythrocytes can be removed by porcine alveolar macrophages. PeerJ, 2019, 7: e6439.
[8] BREKKE O L, CHRISTIANSEN D, KISSERLI A, FURE H, DAHL J A, DONVITO B, REVEIL B, LUDVIKSEN J K, TABARY T, MOLLNES T E, COHEN J H M. Key role of the number of complement receptor 1 on erythrocytes for binding ofto erythrocytes and for leukocyte phagocytosis and oxidative burst in human whole blood. Molecular Immunology, 2019, 114: 139-148.
[9] LI J, WANG J P, GHIRAN I, CERNY A, SZALAI A J, BRILES D E, FINBERG R W. Complement receptor 1 expression on mouse erythrocytes mediates clearance ofby immune adherence. Infection and Immunity, 2010, 78(7): 3129-3135.
[10] LI J, GLOVER D T, SZALAI A J, HOLLINGSHEAD S K, BRILES D E. PspA and PspC minimize immune adherence and transfer of pneumococci from erythrocytes to macrophages through their effects on complement activation. Infection and Immunity, 2007, 75(12): 5877-5885.
[11] SALAM K A, WANG R Y, GRANDINETTI T, DE GIORGI V, ALTER H J, ALLISON R D. Binding of free and immune complex-associated hepatitis C virus to erythrocytes is mediated by the complement system. Hepatology, 2018, 68(6): 2118-2129.
[12] WANG F S, CHU F L, JIN L, LI Y G, ZHANG Z, XU D, SHI M, WU H, MOULDS J M. Acquired but reversible loss of erythrocyte complement receptor 1 (CR1, CD35) and its longitudinal alteration in patients with severe acute respiratory syndrome. Clinical and Experimental Immunology, 2005, 139(1): 112-119.
[13] PEIRIS J S M, LAI S T, POON L L M, GUAN Y, YAM L Y C, LIM W, NICHOLLS J, YEE W K S, YAN W W, CHEUNG M T, CHENG V C C, CHAN K H, TSANG D N C, YUNG R W H, NG T K, YUEN K Y. Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet, 2003, 361(9366): 1319-1325.
[14] DE OLIVEIRA R B, WANG J P, RAM S, GAZZINELLI R T, FINBERG R W, GOLENBOCK D T. Increased survival in B-cell- deficient mice during experimental cerebral malaria suggests a role for circulating immune complexes. mBio, 2014, 5(2): e00949-e00914.
[15] 陳紫晗, 孫琳琳, 張藝璇, 閆軍浩. β-淀粉樣蛋白清除障礙在阿爾茲海默癥發病中的作用. 生理科學進展, 2019, 50(2): 149-152.
CHEN Z H, SUN L L, ZHANG Y X, YAN J H. The roles of amyloid β-protein clearance impairment in the Alzheimer’s disease. Progress in Physiological Sciences, 2019, 50(2): 149-152. (in Chinese)
[16] SAWADA T, FUJIMORI D, YAMAMOTO Y. Systemic lupus erythematosus and immunodeficiency. Immunological Medicine, 2019, 42(1): 1-9.
[17] BRUBAKER W D, CRANE A, JOHANSSON J U, YEN K, GARFINKEL K, MASTROENI D, ASOK P, BRADT B, SABBAGH M, WALLACE T L, GLAVIS-BLOOM C, TENNER A J, ROGERS J. Peripheral complement interactions with amyloid β peptide: Erythrocyte clearance mechanisms. Alzheimer’s & Dementia, 2017, 13(12): 1397-1409.
[18] 谷新利, 李宏全, 王俊東, 蔣建軍, 陳韓英, 李炳奇, 張建海, 劉振中, 劉紅, 羅燕, 邵永斌. 從中藥方劑中提取的復合多糖對雛雞免疫功能的影響. 中國農業科學, 2005, 38(4): 813-820.
GU X L, LI H Q, WANG J D, JIANG J J, CHEN H Y, LI B Q, ZHANG J H, LIU Z Z, LIU H, LUO Y, SHAO Y B. Effects of compound polysaccharide extracted from traditional Chinese medical herbs on the immunity function in chickens. Scientia Agricultura Sinica, 2005, 38(4): 813-820. (in Chinese)
[19] REINAGEL M L, TAYLOR R P. Transfer of immune complexes from erythrocyte CR1 to mouse macrophages. Journal of Immunology, 2000, 164(4): 1977-1985.
[20] HEPBURN A L, MASON J C, WANG S, SHEPHERD C J, FLOREY O, HASKARD D O, DAVIES K A. Both Fcgamma and complement receptors mediate transfer of immune complexes from erythrocytes to human macrophages under physiological flow conditions. Clinical and Experimental Immunology, 2006, 146(1): 133-145.
[21] 張琪琪, 凌小雅, 孫雨晨, 尹偉, 范闊海, 孫娜, 孫耀貴, 李宏全. 豬紅細胞類補體受體Ⅰ型膜結合蛋白的篩選. 中國獸醫科學, 2020, 50(1): 42-48.
ZHANG Q Q, LING X Y, SUN Y C, YIN W, FAN K H, SUN N, SUN Y G, LI H Q. Screening of membrane-binding proteins of complement receptor 1-like on porcine erythrocytes. Chinese Veterinary Science, 2020, 50(1): 42-48. (in Chinese)
[22] 張琪琪. 豬紅細胞CR1-like膜結合蛋白的檢測及鑒定[D]. 太谷: 山西農業大學, 2020.
ZHANG Q Q. Detection and identification of membrane-binding proteins of complement receptor 1-like on porcine erythrocytes[D]. Taigu: Shanxi Agricultural University, 2020. (in Chinese)
[23] 王春, 尹偉, 范闊海, 孫娜, 孫耀貴, 李宏全. 豬肺泡巨噬細胞膜表面類補體受體分子的鑒定. 黑龍江畜牧獸醫, 2019(7): 13-17.
WANG C, YIN W, FAN K H, SUN N, SUN Y G, LI H Q. Identification of surface complement receptor molecules in porcine alveolar macrophages. Heilongjiang Animal Science and Veterinary Medicine, 2019(7): 13-17. (in Chinese)
[24] 王春. 豬肺泡巨噬細胞移除紅細胞免疫粘附致敏GFP-.的研究[D]. 太谷: 山西農業大學, 2019.
WANG C. Study on porcine alveolar macrophages removing opsonized GFP-.from erythrocytes[D]. Taigu: Shanxi Agricultural University, 2019. (in Chinese)
[25] 凌小雅, 朱樂樂, 孫加樂, 王緣, 張睿玉, 薛曉姝, 尹偉. PAMs移除豬紅細胞表面GFP-的體外觀察. 山西農業科學, 2021, 49(9): 1132-1136.
LING X Y, ZHU L L, SUN J L, WANG Y, ZHANG R Y, XUE X S, YIN W.observation of GFP-removed from porcine erythrocytes by porcine alveolar macrophages(PAMs). Journal of Shanxi Agricultural Sciences, 2021, 49(9): 1132-1136. (in Chinese)
[26] RUSSELL B L, GILDENHUYS S. Solubilisation and purification of recombinant bluetongue virus VP7 expressed in a bacterial system. Protein Expression and Purification, 2018, 147: 85-93.
[27] BANEYX F, MUJACIC M. Recombinant protein folding and misfolding in. Nature Biotechnology, 2004, 22(11): 1399-1408.
[28] 崔姣艷.豬紅細胞免疫黏附受體的鑒定與檢測[D]. 太谷:山西農業大學, 2015.
CUI J Y. Identification and detection of porcine erythrocyte immune adhesion receptors[D]. Taigu: Shanxi Agricultural University, 2015. (in Chinese)
[29] LI J, SZALAI A J, HOLLINGSHEAD S K, NAHM M H, BRILES D E. Antibody to the type 3 capsule facilitates immune adherence of pneumococci to erythrocytes and augments their transfer to macrophages. Infection and Immunity, 2009, 77(1): 464-471.
[30] KAVAI M. Immune complex clearance by complement receptor type 1 in SLE. Autoimmunity Reviews, 2008, 8(2): 160-164.
[31] ROCHOWIAK A, NIEMIR Z I. The role of CR1 complement receptor in pathology. Polski Merkuriusz Lekarski: Organ Polskiego Towarzystwa Lekarskiego, 2010, 28(163): 84-88.
[32] ARORA V, VERMA J, DUTTA R, MARWAH V, KUMAR A, DAS N. Reduced complement receptor 1 (CR1, CD35) transcription in systemic lupus erythematosus. Molecular Immunology, 2004, 41(4): 449-456.
Effect of Porcine ECR1-Like Immune Adhesion on PAMs Capturing GFP-
ZHANG Zheng1, LING XiaoYa1, FAN KuoHai2, SUN Na1, SUN PanPan1, SUN YaoGui1, LI HongQuan1, YIN Wei
1Shanxi Key Laboratory for Modernization of TCVM/College of Veterinary Medicine, Shanxi Agricultural University, Taigu 030801, Shanxi;2Shanxi Key Laboratory for Modernization of TCVM/Animal Experimental Center, Shanxi Agricultural University, Taigu 030801, Shanxi
【Objective】The aim of this study was to investigate whether the immune adherence function of porcine erythrocyte complement receptor type 1-like (ECR1-like) could promote porcine alveolar macrophages (PAMs) to capture sensitized genetic engineering bacteria GFP-, in order to explain the molecular mechanism of porcine erythrocyte immunity and its role in innate immunity. 【Method】The level of GFP-captured by PAMs was detected by flow cytometry, colony plate counting and RT-PCR, and the effect of porcine ECR1-like immune adherence on the capture of GFP-by PAMs was analyzed. Flow cytometry and cellular immunofluorescence technique were used to detect the changes of immune adherence function of porcine erythrocytes, and the number of porcine ECR1-like after the sensitized GFP-with ECR1-like immune adherence was removed by PAMs. 【Result】Flow cytometry showed that the average fluorescence intensity of PAMs in porcine erythrocyte adhesion group was significantly higher than that in blank control group (<0.001), while the positive rate of PAMs cells in porcine erythrocyte adhesion group was significantly higher than that in blank control group (<0.05). Colony smear count showed that the capture of GFP-by PAMs in erythrocyte adhesion group was significantly higher than that in blank control group (<0.05). RT-PCR showed that the relative quantity of GFP-in PAMs of erythrocyte adhesion group was significantly higher than that of blank control group (<0.01). Further blocking CR1-like on the surface of porcine erythrocyte, flow cytometry showed that the average fluorescence intensity of PAMs decreased to 256 301.56±9 208.85 (<0.001), and the positive cell rate of PAMs decreased to (88.32±0.92)% (>0.05). Colony count showed that the capture of GFP-in PAMs decreased to (136 666±8 818) CFU/ml (<0.05), and RT-PCR showed that the relative quantity of GFP-in PAMs decreased significantly (<0.01). Using cell flow and circulation interaction technique, it was found that the average fluorescence intensity of GFP-sensitized by porcine erythrocyte immune adherence decreased from 2 892.18±47.76 before circulation to 2 407.43±141.78 (<0.05), and the positive cell rate decreased from (20.58±0.36)% before circulation to (17.39±0.23)% (<0.05). The adhesion level was significantly lower than that before circulation. Meanwhile, the results of indirect immunofluorescence test showed that the average fluorescence intensity of porcine ECR1-like decreased from 344.33±37.92 before to 291.56±11.99 (<0.05), and the positive cell rate decreased from (30.20±1.24)% before to (28.27±0.64)% (<0.05). 【Conclusion】Porcine ECR1-like promoted the capture of sensitized GFP-by PAMs through its immune adhesion function. After PAMs removed sensitized GFP-adhered to the surface of porcine erythrocyte, the activity of CR1-like of porcine erythrocyte decreased, and the immune adhesion function decreased too.
porcine; erythrocyte; PAMs; CR1-like; immune adherence
10.3864/j.issn.0578-1752.2023.19.016
2022-11-26;
2023-08-04
山西省自然科學基金面上項目(20210302123407)、山西省科技創新人才團隊專項(202204051001021)、山西省高等學校科技創新項目(2019L0364)、山西農業大學博士科研啟動項目(2021BQ77)、中獸醫藥現代化重點實驗室建設項目(202104010910015)
張崢,E-mail:zhangzhengjiayou@163.com。通信作者尹偉,E-mail:dkyyinwei@126.com
(責任編輯 林鑒非)
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
