
陸地棉GhNAC1基因的克隆及抗黃萎病功能分析
2023-11-02 07:48:56張曼張進張新雨王國寧王省芬張艷
中國農業科技導報 2023年10期
張曼, 張進, 張新雨, 王國寧, 王省芬, 張艷
(河北農業大學農學院,華北作物改良與調控國家重點實驗室,河北省作物種質資源重點實驗室,河北保定 071000)
棉花作為重要的經濟作物,不僅是紡織工業的關鍵原料,還是我國近億棉農的主要經濟來源。黃萎病(Verticillium wilt)是一種土傳真菌維管束病害,主要由大麗輪枝菌(Verticillium dahliae)、黑白輪枝菌(Verticillium albo-atrum)侵染所致[1]。大麗輪枝菌依靠其微菌核結構可在土壤中存活數年,導致世界范圍內多種作物產生嚴重維管束病害,引起植物葉片黃化、失水萎蔫甚至整株死亡的癥狀[2-3]。黃萎病自1935 年由美國傳入我國后,在黃河流域、長江流域和西北內陸三大棉區不斷發展,每年造成的棉花產量損失約占世界皮棉總產量的10%~30%[4]。20 世紀90 年代后,該病在全國范圍內大暴發,年發生面積約300 萬hm2,經濟損失高達12億元[5]。利用基因工程手段進行抗病防治是最為經濟有效的措施,然而受限于栽培陸地棉遺傳基礎和抗源狹窄,生產上推廣的棉花品種很少能夠達到抗病水平。
病毒誘導的基因沉默(virus-induced gene silence,VIGS)是RNA 介導的轉錄后沉默(post transcriptional gene silencing,PTGS),據此發展出了適用于多種植物的VIGS 技術。與傳統的轉基因技術相比,VIGS 技術具有操作更加簡便、周期更短、效果明顯、適合高通量等優點,在植物功能基因組學領域表現出巨大的潛力。Gao 等[6]基于煙草脆裂病毒(tobacco rattle virus,TRV)載體體系,構建了適用范圍更廣、沉默效果更好的植物VIGS技術系統。基于TRV 載體體系的VIGS技術成功應用于棉花,使一些重要的棉花黃萎病抗性基因獲得很好的功能分析,如GbRvd[7]、GhLAC15[8]、GhGPA[9]、GhnsLTP[10]等。雖然VIGS 技術難以保持長久穩定的沉默效果[11],但是考慮到棉花苗期黃萎病抗性鑒定可以在沉默的幼苗接菌后25~30 d完成,在此過程中,VIGS 完全可以保證穩定的沉默效果,因此利用VIGS 技術可以高效、快速地鑒定棉花抗黃萎病相關基因功能。
NAC(no apical meristem)轉錄因子是一種含有NAM/ATAF/CUC 保守結構域的植物特異性轉錄因子。NAC 轉錄因子于1997 年首次在矮牽牛中被報道,擬南芥ATAF1/2和CUC2基因編碼蛋白的N 端[12]。研究表明,NAC 轉錄因子不僅在植物生長發育中發揮重要作用,而且也參與病原菌防御相關的次生壁形成、乙烯產生等過程[13-15]。如煙草NbNAC062屬于NAC 類膜結合轉錄因子,被PVY(potato virus Y, PVY)侵染后其轉移至細胞核,并可能通過調控UPR(unfolded protein response,UPR)相關基因BiP的表達促進細胞生存,抑制PVY 的早期侵染[16]。接種橡膠樹白粉菌后,擬南芥NAC 家族轉錄因子ATAF2 上調,并與EDS1 發生互作進而正調控擬南芥對白粉菌的抗病性[17]。辣椒CaNAC1能夠快速對細菌性斑點病菌、辣椒輕斑駁病毒的侵染和外源激素作出響應,表達量上調并激活抗病反應[18]。
本課題組培育的抗病高產品種農大601(ND601)2006年通過河北省品種委員會審定,多年多點田間病圃與培養室抗病性鑒定結果均顯示,該品種黃萎病抗性穩定且達到抗病級別,能夠將抗病與高產性狀有效協調,保證其高產穩產。因此,以農大601 為材料,深入挖掘其含有的黃萎病抗性基因對于棉花抗病育種及解析抗病與產量之間的協調關系意義重大。本課題組前期從黃萎病菌脅迫處理后的抗病陸地棉農大601 轉錄組數據庫中發現GhNAC1對大麗輪枝菌的侵染響應快速,推測其可能在早期防御反應中發揮作用。基于此,本研究從農大601 中克隆GhNAC1,利用生物信息學等手段分析該基因的特征,利用實時熒光PCR(real-time PCR,RT-PCR)明確GhNAC1在黃萎病菌處理后的表達模式,應用VIGS技術研究該基因的抗病功能,為進一步探索GhNAC1的抗病機制奠定基礎。
1 材料與方法
1.1 試驗材料
陸地棉抗黃萎病品種農大601 用于克隆GhNAC1,農大棉8 號因具有較高的沉默效果而用于VIGS試驗,6個抗病品種(湯棉7401、慈棉9號、蘇遠04-3、魯890)和6 個感病品種(奎屯系353、73-782、仁洞67-86、73-184、湖南麻陽洋棉花)用于分析不同種基因表達差異,均由本課題組提供。利用濃硫酸對棉花種子進行脫絨處理,清水中浸泡種子過夜,待種子露白后在濕潤的毛巾上催芽,挑選發芽一致(約0.5 cm)的種子播種于裝有蛭石的六棱缽中,于光照培養室中生長,培養條件為16 h 光照、8 h 黑暗,光強6 000~6 500 lx,晝溫28 ℃、夜溫22 ℃,相對濕度40%~50%。
供試大麗輪枝菌(Verticillium dahliae)為生產上強致病力菌系臨西2-1[19],由本課題組分離鑒定,并繼代保存于PDA(potato dextrose agar)培養基。
1.2 基因克隆
棉苗根組織用液氮研磨,使用EASYspi 植物RNA 快 速 提 取 試 劑 盒(AidLab,北 京)和EasyPure?植物DNA提取試劑盒(TransGen,北京)分別提取總RNA 和DNA,應用PrimeScript? ⅡcDNA 合成試劑盒(Takara,大連)進行反轉錄。根據Cotton FGD database(https://cottonfgd.org/)中陸地棉TM-1 的同源序列設計引物(表1),并在農大601 中進行PCR 擴增。PCR 反應體系20 μL:1 μL DNA/cDNA 模板(< 100 ng),正反向引物(表1)各1 μL(10 μmol·L-1),10 μL 10 ×Ex TaqBuffer(Takara,大連)和7 μL 滅菌雙蒸水。PCR 程序:95 ℃ 5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,35 個循環;72 ℃ 10 min。PCR 產物經1%瓊脂糖凝膠電泳進行檢測,膠回收后克隆到pMD?18-T 載體(Takara,大連),轉化大腸桿菌TOP10 后經菌落PCR篩選陽性克隆,送金為智公司進行測序。

表1 試驗所用引物序列Table 1 Primers used in this experiment
1.3 生物信息學分析
利用Cotton FGD 數據庫(https://cottonfgd.org/)進行Blast 比對,同時在NCBI 數據庫(http://www.ncbi.nlm.nih.gov/)中分析目的基因編碼蛋白的功能域。通過在線工具ProtParam(http://web.expasy.org/protparam)預測蛋白氨基酸組成(amino acid composition)、分子量(molecular weight)、理論等電點(theoretical pI)、不穩定指數(instability index)、脂肪指數(aliphatic index)和總平均親水性(grand average of hydropathicity)等,利用在線軟件SignalP 4.0(http://www. cbs. dtu. dk/services/SignalP)預測蛋白信號肽;利用TMpred 在線工具(http://www.ch.embnet.org/software/TMPRED_form.html)分析蛋白跨膜域;利用在線工具CELLO(http://cello.life.nctu.edu.tw/)進行亞細胞定位;利用NovoPro(https://www.novopro.cn/tools/)預測蛋白二級結構。
1.4 亞細胞定位
利用Premier 5.0 設計引物NAC1-G-F/R(表1),以抗病品種農大601 的cDNA 為模板,克隆基因并使用賽默飛世爾Gateway? BP Clonase?Ⅱ Enzyme mix 試劑盒連接到pDONR207 入門載體,測序正確后,使用賽默飛世爾Gateway? LR Clonase? Ⅱ Enzyme mix 試劑盒,連接到pMDC43(含GFP)用于亞細胞定位。將構建好的亞細胞定位載體轉化農桿菌GV3101,28 ℃培養2~3 d,將單克隆搖勻后加入到50 mL LB 液體培養基中,28 ℃搖 床150 r·min-1振 蕩 培 養 至OD600為0.8~1.2。菌液以5 000 r·min-1離心10 min,棄上清并用無菌水重懸后混勻,離心10 min,參照Sparkes等[20]方法配置緩沖液重懸菌液,將沉淀用緩沖液進行重懸,靜止15 min 后備用。使用1 mL 注射器在5~6 周的本氏煙草葉片下表皮上進行注射,23 ℃過夜避光培養,采用激光共聚焦顯微鏡(Fluo View FV1000, Olympus)觀察熒光信號,GFP 激發光和發射光分別為488和550 nm。
1.5 黃萎病菌脅迫處理
將大麗輪枝菌LX2-1 接種到PDA 固體培養基上,25 ℃培養7 d,然后將菌塊接種到PDB(potato dextrose broth)液體培養基[21]中,于25 ℃、150 r·min-1搖床中培養5~7 d。培養好的懸浮孢子液用雙層紗布過濾后,再用滅菌蒸餾水將懸浮孢子液濃度調至1 × 107cfu·mL-1,備用。采用底部注射菌液定量接種法[22]接菌,每缽接種8 mL 菌液,接菌后的棉苗置于晝溫25 ℃、夜溫22 ℃,相對濕度65%~70%,光照不變的條件下繼續培養[23],其間定期澆灌Hoagland 營養液[24]以補充養分。分別在接菌后6、12、24、36 和48 h 取棉苗根組織,保存在-80 ℃備用。每個樣本取3 棵整齊一致的棉苗混樣,每個時間點取3個生物學重復。以蒸餾水代替孢子懸浮液處理的棉苗作為對陰性照組。
1.6 RT-PCR
采用液氮研磨和EASYspi 植物RNA 快速提取試劑盒提取棉苗RNA。通過瓊脂糖電泳和NanoDrop2 000 超微量分光光度計(Thermo Scientific)對RNA 的質量和含量進行檢測。利用PrimeScript ? RT reagent Kit with gDNA Eraser(TaKaRa)并參照說明書完成cDNA 的合成,并用于RT-PCR。RT-PCR 反應體系(20 μL):1 μL cDNA 模板(< 100 ng),10 μL SYBR?Premix Ex TaqTM(TaKaRa),正、反向引物(表1)各0.8 μL(10 μmol·L-1),7.4 μL 滅菌雙蒸水。反應在CFX96 Real-Time PCR Detection System(Bio-Rad)中 進行,反應程序:95 ℃ 15 s;95 ℃ 10 s,58 ℃ 10 s,72 ℃ 15 s,40 個循環。以陸地棉UBQ14(Ghir_D1 0G001850)作為內參,每個反應設置3次生物學重復。采用2-ΔΔCt法[25]進行相對定量分析,并利用GraphPad Prism?6軟件進行統計分析和作圖。
1.7 病級統計
接種后25 d,采用5級分類法[20]對棉苗進行病級統計:0級,植株無病癥;1級,子葉發病,真葉無病癥;2 級,子葉以上的第一片真葉發病;3 級,兩片及以上真葉發病,新葉無病癥;4 級,植株生長點或整株枯死。病情指數(disease index, DI)的計算公式[26]如下。
1.8 棉花VIGS試驗
設計引物NAC1-V-F/R(表1),在GhNAC1序列兩端引入酶切位點EcoRⅠ和KpnⅠ,以含有GhNAC1ORF 的質粒為模板,進行PCR 擴增。PCR 產物經膠回收后克隆至pMD?18-T 載體,再經雙酶切,將回收目的片段克隆至VIGS 載體(pTRV2)。采用凍融法將pTRV2 重組質粒轉入農桿菌GV3101 感受態細胞[27]。參照Gao 等[28]的方法進行遺傳轉化,GhNAC1沉默效果通過RT-PCR檢測,方法同1.4。
2 結果與分析
2.1 GhNAC1的克隆與生物信息學分析
從黃萎病菌脅迫處理后的陸地棉農大601 轉錄組數據庫中,篩選到與棉花抗黃萎病相關的基因Ghir_A01G000170(GhM_A01G0037),該基因含有NAM 結構域,命名為GhNAC1。根據陸地棉TM-1 基因組中GhNAC1的序列信息,設計特異引物進行PCR 擴增,獲得約800 bp 的目的片段。經測序,獲得GhNAC1的ORF 全長序列840 bp,該基因編碼279 個氨基酸殘基。在NCBI 上BLAST 結果顯示,GhNAC1屬于NAM 超級家族,NAM 結構域在10~130 aa位置。
GhNAC1蛋白分子量32.4 kD,理論等電點pI為8.91,包括4 542 個原子,分子式C1456H2264N388O421S13。在組成GhNAC1 蛋白的20 種氨基酸中,賴氨酸(Lys)所占比例最高(9.3%),而半胱氨酸(Cys)所占的比例最低(1.1%)。GhNAC1 的不穩定指數47.78,屬于不穩定蛋白;脂肪指數72.33;總平均親水性-0.592。GhNAC1 沒有信號肽和跨膜結構(圖1A 和B),亞細胞定位預測結果顯示GhNAC1定位在細胞核。蛋白二級結構分析顯示,GhNAC1含有10個α螺旋和10個β折疊(圖1C)。
2.2 GhNAC1的亞細胞定位分析
亞細胞定位預測結果顯示GhNAC1定位于細胞核。為進一步確定GhNAC1 在細胞中的位置,將其與GFP 蛋白進行融合表達。在CaMV 35S 啟動子的控制下,構建了GhNAC1 與GFP 的融合蛋白,并在煙草表皮細胞中瞬時表達。GhNAC1-GFP 融合蛋白侵染煙草表皮后,熒光成像顯示定位在細胞核上(圖2),與預測結果一致。

圖2 GhNAC1亞細胞定位Fig. 2 Subcellular localization of GhNAC1

圖2 GhNAC1蛋白結構分析Fig. 2 Structure analysis of GhNAC1 protein
2.3 GhNAC1的表達模式分析
在四葉期取棉苗不同組織進行表達檢測,結果顯示GhNAC1在葉中的表達顯著高于根和莖(圖3A)。對于抗黃萎病品種ND601,在檢測的5個時間點中,GhNAC1的表達在6、12 和24 phi(hours post infection)均顯著上調(圖3B)。GhNAC1在6 個抗病和6 個感病品種的表達水平檢測結果顯示,GhNAC1在抗病品種的表達水平顯著高于感病品種(圖3C),進一步證明GhNAC1參與了棉花對黃萎病的抗病反應,且可能是參與棉花抗病反應的正調控因子。
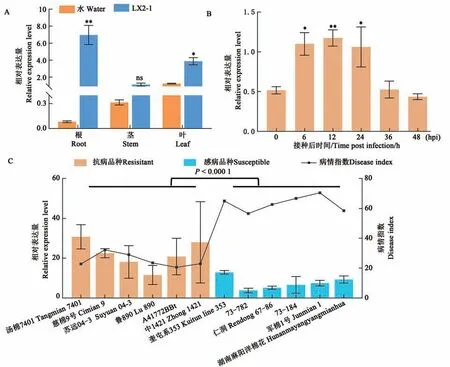
圖3 GhNAC1的表達模式Fig. 3 Expression pattern of GhNAC1
2.4 GhNAC1正調控棉花黃萎病抗性
利用VIGS 處理農大棉8 號7 d 后,沉默CLA1基因的棉苗呈現出新生真葉白化現象(圖4A),表明本研究成功建立了VIGS技術體系。此時,通過RT-PCR 檢測GhNAC1的沉默效果,如圖4B 所示,與對照(CK)棉苗相比,GhNAC1基因在沉默棉苗中的表達量顯著下調,表明棉花內源GhNAC1得到有效沉默,這些沉默植株繼續用于抗黃萎病功能分析。沉默植株與對照植株分別接種臨西2-1,接菌20 d 后發現GhNAC1沉默植株較對照呈現出更加嚴重的葉片黃化、萎蔫等典型的黃萎病癥狀(圖4C)。病情指數統計結果顯示,沉默植株病指為44.1(感病),顯著高于對照植株27.9(耐病)(圖4D)。植株莖桿縱切觀察顯示,沉默植株的維管束褐化程度較對照植株更嚴重(圖4E)。由此可見,抑制GhNAC1基因的表達可顯著降低棉花對黃萎病的抗性,表明GhNAC1正調控棉花抗黃萎病反應。
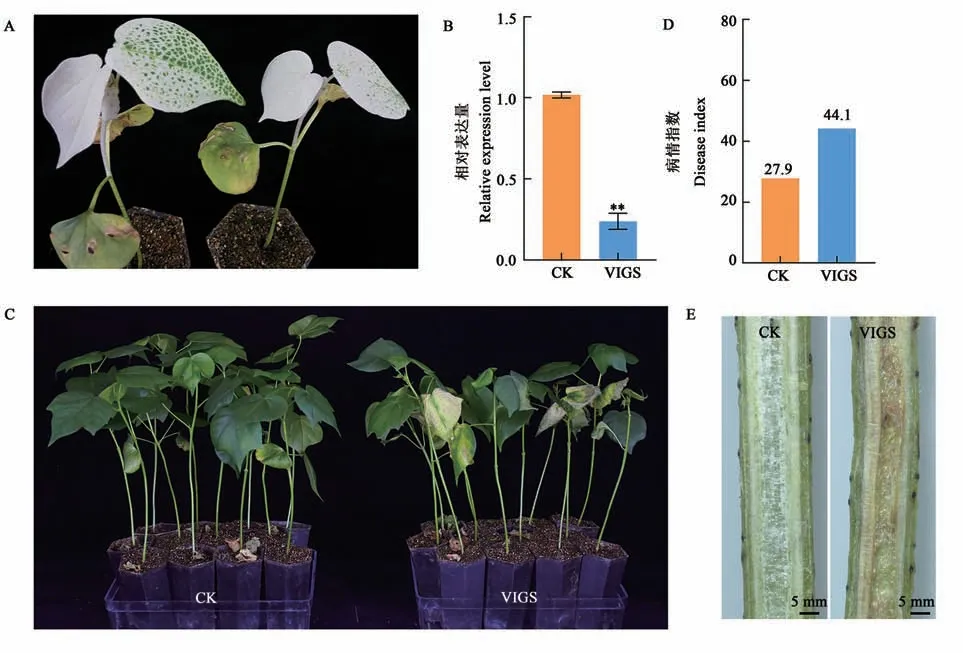
圖4 沉默GhNAC1降低了棉花對大麗輪枝菌的抗性Fig. 4 Silencing GhNAC1 reduced the resistance to V. dahliae in cotton
2.5 沉默GhNAC1 后降低了水楊酸通路相關基因表達
水楊酸(salicylic acid,SA)能夠誘導植物被病原菌侵染后相關蛋白的合成,從而提高植物的抗病性,SA 相關信號途徑被認為是參與生物脅迫的重要抗病路徑[29]。對GhNAC1沉默植株中SA 通路相關的基因進行了檢測,發現沉默植株中SA通路主要標志基因(PAD4、NDR1、NPR1、和PR1)的表達水平顯著下降(圖5),推測GhNAC1 通過激活SA 信號通路增強棉花黃萎病抗性。
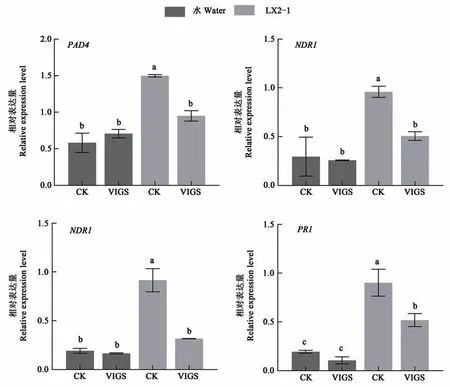
圖5 GhNAC1沉默棉花接菌后水楊酸途徑基因表達分析Fig. 5 Analysis of gene expression of salicylic acid pathway in GhNAC1 silenced cotton after inoculation
3 討論
黃萎病是土傳真菌病害,嚴重影響植物的生長發育,降低作物的品質和產量。黃萎病每年對棉花造成嚴重的產量損失與品質下降[4]。培育抗病品種是防治黃萎病最有效的途徑,挖掘黃萎病抗性基因,闡明基因的抗病功能一直是棉花抗病育種的重要研究內容。近年來研究表明,NAC 轉錄因子在生長、發育、衰老和形態發生等多種生物過程中發揮重要作用,并廣泛參與不同植物激素、多種非生物和生物脅迫響應的信號通路[30]。在模式植物和大田作物中,均有報道NAC基因響應病原菌的感染而誘導表達[31]。本研究在黃萎病菌脅迫處理的cDNA 文庫中鑒定了GhNAC1基因,該基因定位在細胞核,能夠在根部快速響應黃萎病菌誘導表達量顯著升高,并在抗病品種的表達水平顯著高于感病品種,表明GhNAC1基因參與了棉花對黃萎病的抗性,為棉花抗性育種提供新的候選基因。過表達或沉默NAC基因導致植株對病原菌的抗性增強或減弱[32-33],表明NAC 轉錄因子可以正向或負向地調節植物的抗病/防御反應。OsNAC6是參與水稻抗病的NAC 轉錄因子,過表達OsNAC6的轉基因水稻對稻瘟病、缺水和高鹽脅迫的耐受性都有提高[34]。在水稻中過表達OsNAC111、OsNAC58和OsONAC066可以提高水稻對稻瘟病的抗性[19,35];在小麥中,沉默TaNAC2可在條銹菌(Puccinia striiformis f.sp.Tritici,Pst)與小麥互作早期顯著增加H2O2的合成,降低菌絲生長,從而增強植株對條銹病的抗性[36];沉默番茄SlSRN1(solanum lycopersicum stress-related NAC1)降低了番茄對灰葡萄球菌(Botrytis cinerea)的抗性,表明SlSRN1是番茄對灰葡萄球菌防御反應的正調控因子[37]。馬鈴薯中的StNACb4被證明能夠增強對青枯病的抗性[38]。上述研究表明,NAC 轉錄因子參與了植株對多種病原菌的抗病反應。本研究對陸地棉GhNAC1基因進行沉默后發現,相較于TRV∶00植株,TRV∶GhNAC1沉默植株葉片褪綠更明顯,整體表現為萎蔫,大多數沉默植株表現出較高的病害等級(2、3、4),莖稈被真菌侵染力度更大。因此,沉默GhNAC1顯著降低了棉花對黃萎病的抗性,表明GhNAC1正向調控棉花黃萎病抗性,進一步擴充了NAC 轉錄因子調節植物防御反應的內容。
SA 是重要的信號分子,在許多植物病原體相互作用中激活防御反應,特別是針對生物營養體和半生物營養體[39-40]。SA的生物合成途徑有2種,分別是異分支酸途徑(isochorismate synthase,ICS)和苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)途徑,它們都是從分支酸鹽開始的[41],SA合成主要通過ICS途徑,發生在葉綠體中,約占SA合成的90%[42]。此外,染色質免疫沉淀(ChIP)試驗表明,在ICS1和SAGT1啟動子中均含有NAC 核心結合位點,且被ANAC019沉淀富集。 因此,ANAC019、ANAC055、ANAC072可能分別通過抑制ICS1和誘導SAGT降低擬南芥SA 的合成,增強SA的代謝,成為SA 積累的負轉錄調控因子[43]。本研究對沉默植株SA 信號通路相關基因(PAD4、NDR1、NPR1和PR1)的表達進行了檢測,發現它們在沉默植株中明顯受到抑制,表明GhNAC1參與了SA 介導的抗病信號通路。上述結果為深入研究GhNAC1的抗病機制及分子調控網絡奠定了基礎。
本研究克隆了GhNAC1,并驗證了其在黃萎病抗性中的作用。GhNAC1是棉花黃萎病抗性的正調控基因,可能通過參與SA信號途徑增強棉花對黃萎病抗性,為棉花抗病育種提供新的候選基因。
