LncRNA CDKN2BAS通過調控miR-627-3p影響肺癌細胞增殖、凋亡和放射敏感性的機制
2023-11-07 00:41:46秦玉春蔡林再謝景臣鄺向東
中國老年學雜志 2023年21期
秦玉春 蔡林再 謝景臣 鄺向東
(海南西部中心醫院呼吸與危重癥醫學科,海南 儋州 571700)
非小細胞肺癌(NSCLC)發病率、死亡率較高,且具有惡性程度高、侵襲性強的特點,目前主要治療方法是放療、手術和化療,但患者的預后仍不理想〔1,2〕。因此,亟待深入研究NSCLC病理機制以指導臨床治療。研究表明,長非編碼RNA(LncRNA)為人類多種癌癥的潛在治療靶點,其中LncRNA CDKN2BAS與多種惡性腫瘤相關。研究表明,LncRNA CDKN2BAS可促進卵巢癌細胞的增殖和遷移,也可通過miR-153-5p/Rho GTP酶激活蛋白(ARHGAP)18信號軸促進肝細胞癌轉移,與肝細胞癌患者預后不良相關,亦能靶向微小RNA-98-5p抑制骨肉瘤細胞的惡性生物學行為〔3~5〕。但是LncRNA CDKN2BAS對肺癌細胞是否具有類似作用及作用機制目前尚未見報道。miR-627-3p在肺癌組織和細胞中表達下調,LncRNA RP11-284F21.9通過調節miR-627-3p促進肺癌細胞增殖和侵襲〔6〕。盡管已有相關研究報道miR-627-3p在肺癌中的表達,但LncRNA CDKN2BAS是否與其相關目前仍不明確。本文采用LncBase數據庫預測LncRNA CDKN2BAS與miR-627-3p間的關系,體外實驗研究LncRNA CDKN2BAS對肺癌細胞增殖、凋亡和放射敏感性的作用及其機制。
1 材料與方法
1.1臨床資料 選取2017年1月至2019年1月于海南西部中心醫院肺癌患者42例,手術收集原發腫瘤組織及癌旁組織,患者均知情且同意,手術切除后立即將樣本保存在-80 ℃。其中男25例,女17例;年齡30~60歲,平均(43.14±6.78)歲;病理分型:腺癌30例,鱗癌12例;淋巴結轉移23例,未轉移19例。本研究經醫院倫理委員會批準。
1.2材料 人正常肺支氣管上皮細胞BEAS-2B和肺癌細胞系NCI-H1299、NCI-H2170、PNCI-H1975(中科院上海細胞庫),DMEM培養基、胎牛血清(美國Hyclone公司),Trziol、反轉錄、聚合酶鏈反應(PCR)試劑盒(寶日醫生物),引物合成(上海生工),噻唑藍(MTT)、膜聯蛋白Ⅴ-異硫氰酸熒光素/碘化丙啶(Annexin Ⅴ-FITC/PI)凋亡檢測試劑盒(上海晶抗生物),RIPA蛋白裂解液、二喹啉甲酸(BCA)試劑盒(上海研謹生物),si-NC、si-LncRNA CDKN2BAS、miR-NC、miR-627-3p、anti-miR-NC、anti-miR-627-3p(上海吉瑪制藥),pcDNA、pcDNA-LncRNA CDKN2BAS(上海索寶生物),Lipofectamine2000(美國Invitrogen)。
1.3細胞轉染與分組 將NCI-H1299細胞接種于6孔板(1×105/ml),分為si-NC組、si-LncRNA CDKN2BAS組、miR-627-3p組和miR-NC組,分別轉染si-NC、si-LncRNA CDKN2BAS、miR-627-3p,miR-NC組的NCI-H1299細胞不做任何處理,另設si-LncRNA CDKN2BAS+anti-miR-NC組和si-LncRNA CDKN2BAS+miR-627-3p組,分別轉染si-LncRNA CDKN2BAS+anti-miR-NC和si-LncRNA CDKN2BAS+miR-627-3p,轉染步驟嚴格按照試劑盒要求進行。
1.4實時熒光定量(RT-q)PCR實驗 Trizol法分別提取臨床收集組織及各組細胞總RNA,檢測純度和濃度并定量,并進行反轉錄,配制PCR擴增體系,在PCR儀上進行擴增,獲取各組細胞Ct值,LncRNA CDKN2BAS、miR-627-3p的內參分別為GAPDH、U6,各組細胞中LncRNA CDKN2BAS和miR-627-3p的mRNA相對表達水平采用2-ΔΔCt法計算。
1.5克隆形成實驗 將各組細胞接種6孔板,每孔約為1×103,按照源靶距15 cm給予用不同劑量X線照射(0、2、4、6、8 Gy),繼續培養14 d左右至肉眼可見明顯集落形成,磷酸鹽緩沖液(PBS)沖洗細胞,經多聚甲醛固定和結晶紫染色后,顯微鏡下計數集落數(細胞數>50個),計算細胞存活分數和放射生物學參數。
1.6MTT實驗 將各組細胞接種于96孔板(2.5×103/ml),繼續培養48 h,加MTT 20 μl,繼續培養4 h,去除上清,加二甲基亞砜150 μl,震蕩孵育5 min,檢測450 nm波長處的各組細胞吸光度A值(OD450)。
1.7流式細胞術 收集各組細胞,PBS沖洗2次,按試劑盒說明書加入相應試劑后,37 ℃反應20 min,流式細胞儀檢測細胞凋亡。
1.8Western印跡實驗 提取細胞總蛋白并進行蛋白定量,95 ℃水浴變性,制備電泳分離膠、濃縮膠,每個電泳槽上樣60 μg,常規電泳、電轉后,蛋白條帶用5%脫脂牛奶封閉2 h,孵育一抗稀釋液(1∶800),4 ℃過夜,室溫孵育二抗稀釋液2 h(1∶2 000),以β-actin為內參,化學發光顯色,凝膠系統成像后分析條帶灰度值,計算蛋白表達水平。
1.9雙熒光素酶報告實驗 根據LncRNA CDKN2BAS與miR-627-3p的結合位點構建熒光素酶表達載體,將NCI-H1299細胞分為4組,分別轉染LncRNA CDKN2BAS野生型(WT)熒光素酶表達載體+miR-NC,LncRNA CDKN2BASWT熒光素酶表達載體+miR-627-3p和LncRNA CDKN2BAS突變型(MUT)熒光素酶表達載體+miR-NC,LncRNA CDKN2BASMUT熒光素酶表達載體+miR-627-3p,繼續培養48 h后,檢測熒光素酶活性。
1.10統計學處理 采用SPSS24.0軟件進行獨立樣本t檢驗、單因素方差分析。
2 結 果
2.1臨床收集組織中LncRNACDKN2BAS、miR-627-3p表達情況 與癌旁組織(1.00±0.16)比較,肺癌組織中LncRNA CDKN2BAS mRNA相對表達量(3.28±0.37)顯著上調(t=36.655,P<0.001);肺癌組織miR-627-3p mRNA相對表達量(0.45±0.06)較癌旁組織(1.00±0.13)顯著下調(t=24.895,P<0.001)。
2.2在肺癌細胞系中LncRNA CDKN2BAS和miR-627-3p表達 與BEAS-2B細胞比較,其余各組細胞中LncRNA CDKN2BAS mRNA相對表達量顯著上調,miR-627-3p顯著下調(P<0.05),見表1。
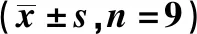
表1 在肺癌細胞系中LncRNA CDKN2BAS和miR-627-3pmRNA相對表達量
2.3低表達LncRNA CDKN2BAS對NCI-H1299細胞的影響 與si-NC組相比,si-LncRNA CDKN2BAS組LncRNA CDKN2BAS mRNA相對表達量顯著下調,CyclinD1蛋白表達水平明顯降低,Cleaved-caspase-3和γ-H2AX蛋白表達水平和細胞凋亡率明顯升高,細胞活力明顯降低(P<0.05);以不同劑量(0、2、4、6、8 Gy)照射NC組、si-NC組、si-LncRNA CDKN2BAS組NCI-H1299細胞,細胞存活分數呈劑量依賴性降低,且si-LncRNA CDKN2BAS組低于si-NC組(P<0.05),見圖1、圖2、表2。單擊多靶模型參數見表3。

1~7:NC組、si-NC組、si-LncRNA CDKN2BAS組、miR-NC組、miR-627-3p組、si-LncRNA CDKN2BAS+anti-miR-NC組、si-LncRNA CDKN2BAS+anti-miR-627-3p組圖1 各組細胞CyclinD1、Cleaved-caspase-3、γ-H2AX蛋白表達

圖2 低表達LncRNA CDKN2BAS及高表達miR-627-3p對NCI-H1299細胞凋亡的影響
表2 低表達LncRNA CDKN2BAS對肺癌細胞NCI-H1299增殖、凋亡和放射敏感性的影響

表3 低表達LncRNA CDKN2BAS對NCI-H1299細胞放射敏感性的影響
2.4高表達miR-627-3p對肺癌細胞NCI-H1299的影響 與miR-NC組相比,miR-627-3p組miR-627-3p mRNA相對表達量明顯上調,CyclinD1蛋白表達明顯降低,Cleaved-caspase-3、γ-H2AX蛋白表達和細胞凋亡率明顯升高,細胞活力明顯降低(P<0.05);細胞存活分數呈劑量依賴性降低,miR-627-3p組低于miR-NC組(P<0.05),見圖1、圖2、表4。放射敏感性相關參數見表5。
表4 高表達miR-627-3p對NCI-H1299細胞的影響

表5 高表達miR-627-3p對NCI-H1299細胞放射敏感性的影響
2.5LncRNA CDKN2BAS 靶向miR-627-3p,調控miR-627-3p的表達 LncRNA CDKN2BAS與miR-627-3p互補的核苷酸序列見圖3。與轉染miR-NC的LncRNA CDKN2BAS-WT比較,轉染miR-627-3p的熒光素酶活性顯著降低(P<0.05),見表6。比較各組miR-627-3p mRNA相對表達量,pcDNA-LncRNA CDKN2BAS組明顯低于pcDNA組,si-LncRNA CDKN2BAS組則高于si-NC組(P<0.05),見表7。
表6 miR-NC或miR-627-3p與LncRNA CDKN2BAS-WT及MUT報告質粒共轉染NCI-H1299細胞后雙熒光素酶活性檢測
表7 RT-qPCR檢測各組miR-627-3p 的表達
2.6下調miR-627-3p能夠逆轉LncRNA CDKN2BAS低表達對NCI-H1299細胞增殖、凋亡和放射敏感性的影響 與si-LncRNA CDKN2BAS+anti-miR-NC組比較,si-LncRNA CDKN2BAS+anti-miR-627-3p組miR-627-3p mRNA相對表達量明顯下調,CyclinD1蛋白水平和細胞活力明顯升高,Cleaved-caspase-3和γ-H2AX蛋白水平和細胞凋亡率明顯降低(P<0.05);細胞存活分數呈照射劑量依賴性降低,且si-LncRNA CDKN2BAS+anti-miR-627-3p組顯著高于si-LncRNA CDKN2BAS+anti-miR-NC組(P<0.05),見圖1、圖4、表8。單擊多靶模型參數見表9。

1,2:si-LncRNA CDKN2BAS+anti-miR-NC組、si-LncRNA CDKN2BAS+anti-miR-627-3p組圖4 下調miR-627-3p 能夠逆轉LncRNA CDKN2BAS低表達對NCI-H1299細胞凋亡的影響
表8 下調miR-627-3p 能夠逆轉LncRNA CDKN2BAS低表達對NCI-H1299細胞增殖、凋亡和放射敏感性的影響

表9 下調miR-627-3p 能夠逆轉LncRNA CDKN2BAS低表達對NCI-H1299細胞放射敏感性的影響
3 討 論
眾多研究表明,LncRNA異常表達與肺癌細胞的惡性表型有關。如過表達LncRNA鋅指蛋白674反義RNA(ZNF674-AS)1抑制NSCLC細胞的增殖、集落形成和腫瘤發生,并伴隨細胞周期阻滯〔7〕。LncRNA叉頭框轉錄因子D3反義RNA(FOXD3-AS)1低表達與NSCLC患者淋巴結轉移陽性和腫瘤分級較高密切相關,上調FOXD3-AS1可抑制NSCLC細胞的增殖和侵襲,下調其表達則效果相反〔8〕。LINC01089可抑制NSCLC細胞的增殖、遷移和侵襲〔9〕。高表達LncRNA TPTEP1對NSCLC表現出顯著的抗腫瘤作用〔10〕。Lnc-THOR與胰島素樣生長因子(IGF)2 mRNA結合蛋白結合后可促進NSCLC細胞增殖〔11〕。LncRNA H19在抗輻射NSCLC細胞中高表達,敲除LncRNA H19可提高NSCLC細胞對X射線和碳離子照射的敏感性〔12〕。敲除LncRNA PKMYT1AR可抑制NSCLC細胞的增殖、遷移和異種移植瘤形成能力〔13〕。體內敲除LINC01296可抑制NSCLC細胞增殖、遷移并促進其凋亡〔14〕。本實驗結果與上述研究結果相似,提示低表達LncRNA CDKN2BAS可降低NCI-H1299細胞的增殖能力,促進細胞凋亡和提高放射敏感性。
研究表明,LncRNA CDKN2BAS可作為ceRNA負調控腫瘤相關miR表達來促進腫瘤發生〔4,5〕。研究表明,miR-627-3p可在腫瘤進展中發揮抑癌基因作用。下調miR-627-3p表達可促進骨肉瘤細胞的增殖和轉移,而其過表達則具有相反效果〔15〕。miR-627-3p在食管鱗狀細胞癌中表達下降,低表達miR-627-3p與食管鱗狀細胞癌患者淋巴結轉移相關,不利于預后,通過細胞實驗證實miR-627-3p能調控食管鱗狀癌細胞遷移、侵襲〔16〕。本研究與上述研究結果一致。本實驗結果提示,miR-627-3p調控NSCLC細胞增殖、凋亡和放射敏感性,且靶向干擾miR-627-3p表達能夠逆轉低表達LncRNA CDKN2BAS對NSCLC細胞的影響,表明LncRNA CDKN2BAS可靶向miR-627-3p調控NSCLC細胞的增殖、凋亡和放射敏感性。
綜上,LncRNA CDKN2BAS在肺癌組織中表達上調,靶向miR-627-3p調控NCI-H1299細胞的增殖、凋亡和放射敏感性,具有成為治療肺癌的新靶點的潛力,本研究的局限性在于僅進行了體外實驗,未來將開展體內實驗進行深入研究。
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
中國塑料(2016年3期)2016-06-15 20:30:00
醫學研究雜志(2015年12期)2015-06-10 06:57:46
