墨蘭CsAP1-A 基因克隆及表達分析
2023-11-13 07:16:10劉嘉超楊鳳璽
廣東農業科學 2023年9期
周 榮,劉嘉超,,楊鳳璽
(1.佛山科學技術學院,廣東 佛山 528000;2.廣東省農業科學院環境園藝研究所/廣東省園林花卉種質創新綜合利用重點實驗室,廣東 廣州 510640)
【研究意義】墨蘭(Cymbidiumsinense)是我國國蘭的典型代表之一,在福建、廣東、臺灣廣泛分布。墨蘭葉型獨特,花姿優美,花香馥郁,花期長(9 月至翌年3 月),具有很高的觀賞價值和經濟價值[1-3],產品遠銷韓國、日本等地。開花性狀是墨蘭最重要的育種目標,而APETALA1(AP1)基因在植物成花轉變和花器官發育過程中具有非常關鍵的作用[4]。開展墨蘭AP1基因的克隆和表達分析研究,對探究墨蘭開花過程中的分子調控機制具有重要意義。【前人研究進展】MADS-box 基因家族是一類在植物中廣泛存在的轉錄因子家族,可與其他轉錄因子和調控因子相互作用,共同在植物花器官分化、果實發育和開花調控等方面起作用。MADSbox 基因家族通常分為兩個亞家族(Type Ⅰ和Type Ⅱ),其中Type Ⅱ亞家族基因編碼的蛋白質均有1 個特征性結構域K,能夠折疊形成3 個α螺旋結構,介導MADS-box 蛋白之間形成二聚體,通過蛋白復合體共同調控下游靶基因。以Type Ⅱ類MADS-box 基因為核心構成的花發育ABCDE模型決定植物花器官的發育[5-6]。A 類基因主要影響花朵的外輪花器官發育,同時與E 類基因共同參與萼片的發育,并與B 類基因和E 類基因共同調控花瓣的形成和特化。AP1屬于A 類基因,其突變或過表達都會對植物的開花時間和花器官產生顯著影響[7-8]。在擬南芥中,過表達AP1可使擬南芥提前1 周左右開花[9]。AP1同時可以抑制最外輪花器官萼片葉腋處形成更多花原基。另外,AP1可以分別通過抑制細胞分裂素合成基因和激活細胞分裂素降解酶,減少AP1基因表達區域的細胞分裂素含量,從而維持正常花分生組織的有限生長。目前,建蘭(C.ensifolium)[10]、石 斛(DendrobiumChao Praya Smile)[11]、萼脊蘭(Sedireajaponica)[8]、牡丹(Paeonia suffruticosa)[12]、荔枝(Litchichinensis)[13]、大豆(Glycinemax)[14]等多種植物的AP1基因已被成功克隆,研究發現,它們在花器官的形成和花期調控中起著非常重要的作用。【本研究切入點】目前關于墨蘭AP1基因的研究報道甚少,該基因在墨蘭上的功能表現也尚未清楚。因此,本研究以墨蘭為研究對象,克隆并鑒定到1 個墨蘭AP1基因(命名為CsAP1-A),并進行基因表達模式分析[15]。【擬解決的關鍵問題】本研究克隆墨蘭CsAP1-A基因,通過生物信息學分析其基因結構、蛋白結構域和進化關系,利用RTqPCR 方法分別檢測CsAP1-A在墨蘭不同器官、不同花發育階段和不同花組織部位的表達情況,通過轉錄組測序分析CsAP1-A在5 個不同花型墨蘭品種花組織部位的表達情況,并采用蛋白互作預測軟件分析CsAP1-A 與其他蛋白的互作關系,為進一步研究CsAP1-A 在墨蘭花發育過程的功能奠定基礎。
1 材料與方法
1.1 試驗材料
基因克隆、表達分析供試的墨蘭品種為‘小香’,栽培于廣東省農業科學院環境園藝研究所國家墨蘭種質資源圃內。挑選3 株長勢良好的墨蘭,采集其花芽、成熟花器官及根、莖、葉和果各3 g 左右,用液氮速凍,-80 ℃保存備用。
不同花型轉錄組測序分析供試的5 個墨蘭品種為‘瑞金’(WT 普通花型)、‘安康梅’(花瓣蕊柱化的梅瓣花型,Gynostegium-like petal variety,GPV)、‘大圣’(重瓣花型,Multi perianth variety,MPV)、‘金鳳蝶’(花瓣唇瓣化花型,Labellum -like petal variety,LaPV)和‘六瓣花’(唇瓣萼片化花型,Null-labellum variety,NLV),均來源于廣東省農業科學院環境園藝研究所國家墨蘭種質資源圃。采集的花朵立即用液氮速凍處理。
DH5α 化學感受態細胞、熒光定量試劑盒(Taq Pro Universal SYBR qPCR Master Mix)、高保真酶試劑盒(2×Phanta Flash Master Mix)、產物純化試劑盒(FastPure Gel DNA Extraction Mini Kit)、克隆載體試劑盒(5 min TA/Blunt Cloning Kit)和RNA 提取試劑盒(FastPure Universal Plant Total RNA Isolation Kit),均購自南京諾唯贊生物科技股份有限公司。
1.2 總RNA 提取及反轉錄
用多糖多酚植物RNA 提取試劑盒提取墨蘭花芽的RNA,操作過程嚴格按照試劑盒說明書進行。使用蛋白核酸分析儀檢測RNA 濃度,使用1%瓊脂糖凝膠電泳檢測RNA 質量,將檢測合格的RNA 進行反轉錄反應得到cDNA。
1.3 目的基因克隆
以前期基因組測序得到的CsAP1-ACDS 為參考序列,使用NCBI 設計基因克隆的PCR 引物:CsAP1-A-F 5'ATGGGAAGAGGGAGGGTT3'、CsAP1-A-R 5'TTATCCATTCATATGAGTGAGC3'。以墨蘭花芽cDNA 為模板,按照高保真酶試劑盒說明書操作進行CsAP1-A基因克隆,PCR 體系為50 μL,反應程序:98 ℃預變性30 s;98 ℃預變性10 s、58 ℃退火5 s、72 ℃延伸5 s,34 個循環;72 ℃延伸1 min。擴增產物回收純化后與TA 克隆載體連接,轉化到DH5α 化學感受態細胞中培養,次日挑選陽性菌液送至生工生物工程(上海)股份有限公司測序,得到CsAP1-A基因完整序列。
1.4 CsAP1-A 生物信息學分析
使用Expasy-ProtParam(https://www.expasy.org/)分析CsAP1-A 蛋白的分子量大小、親水性等理化性質;使用NCBI(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析CsAP1-A 的保守結構域;使用DNAMAN 軟件將CsAP1-A 與6 個同源蛋白進行多序列比對;使用Mega6.0 軟件的鄰接法構建CsAP1-A 系統進化樹。
1.5 CsAP1-A 基因表達分析
使用RT-qPCR 方法對CsAP1-A在墨蘭不同器官(根、莖、葉、花、果)、不同花發育階段(花芽分化初期S1、花芽分化發育期S2、花梗伸長期S3、排鈴期S4 和開花期S5)和不同花組織部位(萼片、花瓣、唇瓣、合蕊柱)的表達進行分析。以CsActin作為內參基因(表1),PCR 反應程序為:95 ℃ 2 min;95 ℃ 15 s、60 ℃ 30 s、72 ℃ 30 s,40 個循環。基因表達試驗重復3 次,使用2-ΔΔCt公式計算CsAP1-A相對表達量[16]。
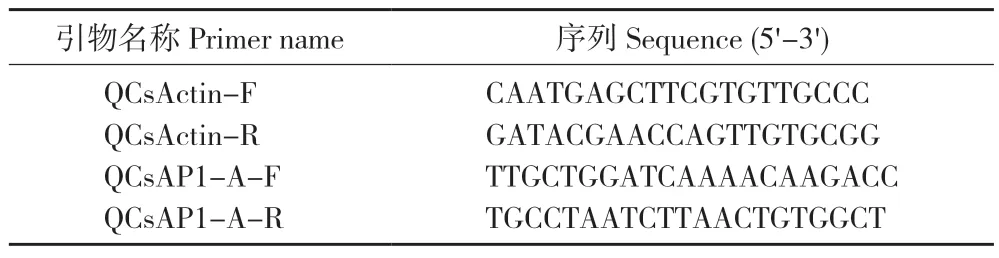
表1 CsAP1-A 基因熒光定量PCR 引物Table 1 Primers for fluorescence quantitative PCR of CsAP1-A gene
1.6 不同花型轉錄組測序分析
采集5 個不同花型(WT、GPV、MPV、LaPV、NLV)墨蘭品種(‘瑞金’‘安康梅’‘大圣’‘金鳳蝶’‘六瓣花’)的花朵,立即液氮速凍處理,送至生工生物工程(上海)股份有限公司進行轉錄組測序。對CsAP1-A的FPKM 值進行log2轉換,并利用R 語言中的Pheatmap 工具包生成熱圖。
1.7 CsAP1-A 蛋白互作關系
通過STRING 數據庫(https://cn.string-db.org/)預測可能與CsAP1-A 發生互作的蛋白質,并以水稻作為參考,構建CsAP1-A 與其他蛋白的互作關系圖。
2 結果與分析
2.1 CsAP1-A 基因全長克隆
以墨蘭花芽為材料提取RNA,并將RNA 反轉錄為cDNA。以cDNA 為模板,以墨蘭基因組參考序列設計引物,擴增得到約750 bp 的單一片段(圖1)。將回收的DNA 片段與TA 克隆載體進行連接并轉化感受態細胞,最后通過測序得到CsAP1-A基因序列,全長744 bp,編碼248 個氨基酸(圖2)。
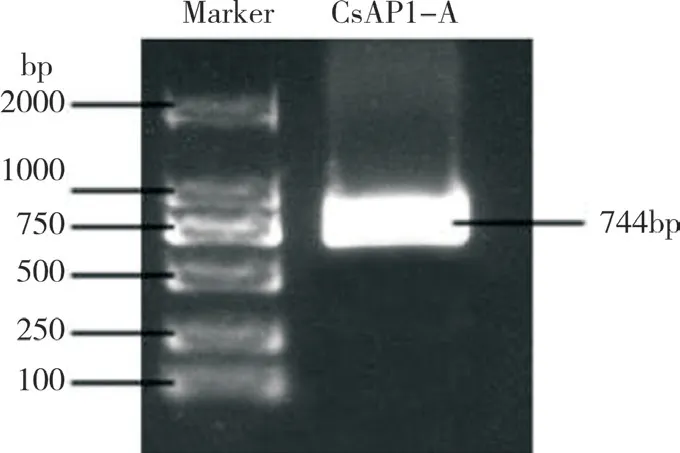
圖1 CsAP1-A 基因克隆PCR 產物Fig.1 PCR product of CsAP1-A gene cloning

圖2 CsAP1-A 基因的核苷酸序列和推導的氨基酸序列Fig.2 Nucleotide acid sequence of CsAP1-A gene and deduced amino sequence
2.2 CsAP1-A 生物信息學分析
2.2.1 CsAP1-A 蛋白理化性質分析根據Expasy-ProtParam 預測分析結果,CsAP1-A 蛋白的分子式為C1242H2038N370O376S9,蛋白分子量為28.4 kD,總原子數為4 035,脂肪系數為83.40,平均親水性為-0.762,說明CsAP1-A 是一個親水蛋白。選擇模型一致性高達98.38%的同源蛋白AOA1P8SCC5.1A 作為模板,通過SWISS-MODEL在線軟件構建CsAP1-A 蛋白的三級結構,發現該結構包含4 個保守的α-螺旋和2 個β 折疊結構(圖3)。
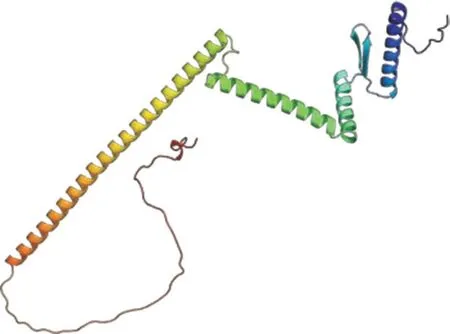
圖3 CsAP1-A 蛋白三級結構預測Fig.3 Tertiary structure prediction of CsAP1-A protein
2.2.2 CsAP1-A 蛋白保守域分析與同源序列比對 通過NCBI 數據庫分析蛋白序列可知,CsAP1-A 擁有保守的MADS-box 結構域和K-box結構域(圖4),屬于MADS-box轉錄因子家族成員。此外,通過NCBI 數據庫得到多個與CsAP1-A 同源性很高的蛋白序列,其中,CsAP1-A 與春蘭(C.goeringii)AP1/FUL(APY18447.1)和蕙蘭(C.faberi)MADS1(AGE15496.1)的同源性均高達98.38%,與金釵石斛(D.nobile)MADSbox protein 1(ABQ08573.1)、蝴蝶蘭(Phalaenopsis amabilis)AP1-related protein(AAZ76263.1)、華山姜(Alpiniaoblongifolia)APETALA1-like protein(ABS83558.1)、銀合歡(Nelumbonucifera)APETALA1(AGY54940.1)的同源性分別為85.83%、82.19%、66.27%和60.32%(圖5)。

圖4 CsAP1-A 蛋白保守結構域分析Fig.4 Analysis of conserved domains of CsAP1-A protein

圖5 CsAP1-A 蛋白同源序列比對Fig.5 Homologous sequence alignment of CsAP1-A protein
2.2.3 CsAP1-A 氨基酸序列的系統進化 為進一步了解CsAP1-A 蛋白的系統進化情況,從NCBI數據庫中挑選20 條與CsAP1-A 氨基酸序列具有一定同源性的氨基酸序列,利用MEGA6.0 軟件的NJ 法構建進化樹。結果(圖6)表明,CsAP1-A的進化樹可以分為單子葉植物和雙子葉植物兩類[17],其中在單子葉植物中,CsAP1-A 與其他蘭科植物親緣關系較近,并聚類一起。

圖6 CsAP1-A 蛋白NJ 系統進化樹Fig.6 Neighbor-joining phylogenetic tree of CsAP1-A protein
2.3 CsAP1-A 基因時空表達分析
為研究CsAP1-A的表達模式,用RT-qPCR方法分別檢測其在墨蘭根、莖、葉、花、果不同器官,不同花發育階段,以及萼片、花瓣、唇瓣、合蕊柱不同花朵組織部位的表達情況。結果(圖7)表明,CsAP1-A在墨蘭不同器官中均有表達,在花中表達量最高,其次分別為莖、果、根和葉。
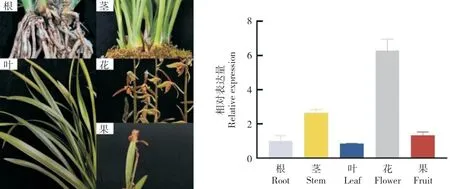
圖7 CsAP1-A 在墨蘭不同器官中的相對表達量Fig.7 Relative expression of CsAP1-A in different organs of Cymbidium sinense
在墨蘭花芽分化發育的不同階段,CsAP1-A集中在花發育早期階段表達,在S1階段的表達量最高,之后逐漸下降,在S3 階段的表達量降至60%,在S5 階段的表達量降至最低。此外,對墨蘭不同花組織部位的表達分析發現,CsAP1-A在合蕊柱中的表達量最高,其次分別是唇瓣、花瓣和萼片(圖8)。
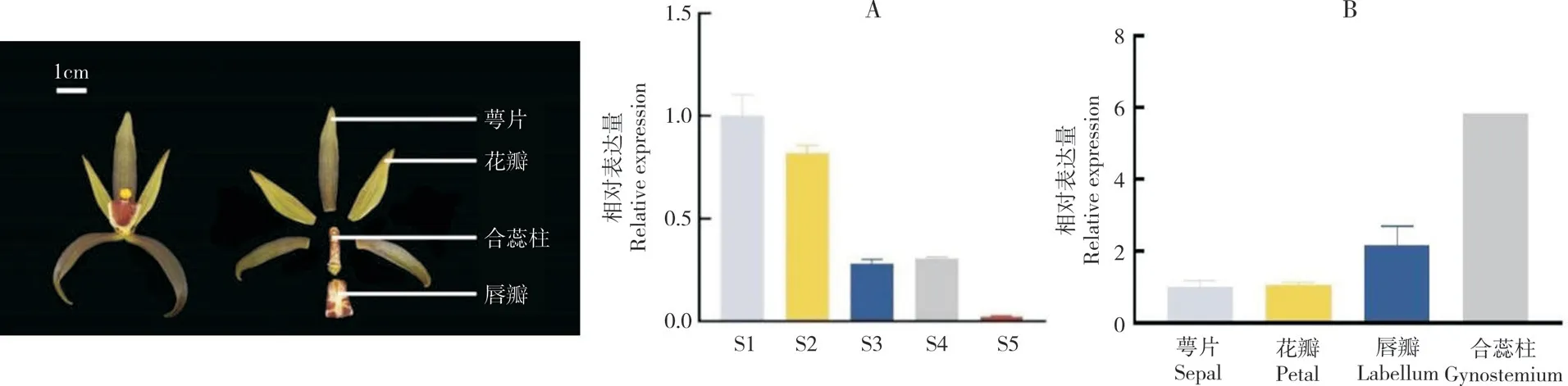
圖8 CsAP1-A 在墨蘭花中的相對表達量Fig.8 Relative expression of CsAP1-A in flower of Cymbidium sinense
2.4 不同花型墨蘭CsAP1-A 表達模式分析
分析墨蘭不同花型轉錄組數據集,用熱圖形式顯示不同花型CsAP1-A基因的表達情況。結果(圖9)表明,CsAP1-A在WT、MPV、LAPV 和NLV 4 種花型[18]中均以合蕊柱中的表達量最高、萼片中的表達量最低;在合蕊柱異常發育的MPV中,CsAP1-A在合蕊柱的表達量顯著提高,而在GPV 中的整體表達量最高,且表達區域從合蕊柱擴展到花瓣。
2.5 CsAP1-A 蛋白互作預測分析
使用String 在線網站預測CsAP1-A 蛋白與其他蛋白之間的互作可能性,并以水稻作為參考,結果(圖10)表明,CsAP1-A 可能與MADS1(A類基因)、MADS6(D 類基因)、MADS47(B 類基因)、MADS8(C 類基因)等10 個蛋白存在互作關系。MADS1 與MADS47 的綜合得分較高,推測CsAP1-A 與它們互作的可能性更大。而在水稻中,OsMADS1 和OsMADS6 蛋白均屬于MADSbox 轉錄因子家族成員,且這些蛋白在花器官發育和花期調控方面發揮重要作用[19]。因此,推測CsAP1-A 蛋白參與墨蘭花的發育。
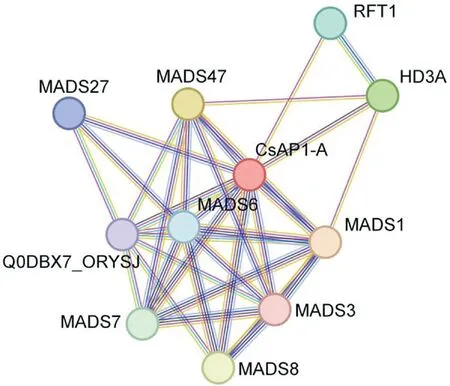
圖10 CsAP1-A 蛋白互作關系預測Fig.10 Prediction of CsAP1-A protein interactions
3 討論
MADS-box 轉錄因子在調控植物花的形態和發育過程方面扮演著重要角色,而AP1基因是MADS-box 轉錄因子家族的重要成員,被歸類為擬南芥ABCDE 開花模型中的A 類基因,在許多植物中均被證實能夠發揮調控植物開花、調控花器官發育的作用[20]。五峰玉蘭MawuAP1基因可以使擬南芥提前開花,并在花序頂端產生頂生花[21]。過表達麻風樹JcAP1基因能夠顯著縮短擬南芥的生長周期并導致提前開花[22]。過表達月季RcAP1也可以使擬南芥提前開花[23]。
本研究從墨蘭花芽中克隆出CsAP1-A基因,該基因編碼區為744 bp,編碼248 個氨基酸,含有MADS-box 和K-box 結構域,表明CsAP1-A是典型MADS-box 轉錄因子。通過與其相似度較高的蛋白進行多序列比對,發現它們的保守結構域相同。系統進化樹分析表明,CsAP1-A 蛋白與春蘭AP1/FUL3 蛋白聚為一支,說明二者親緣關系最近。
大多數A 類基因在生殖器官的表達量均高于營養器官[24]。本研究比較了CsAP1-A基因在墨蘭根、莖、葉、花、果5 個器官的相對表達量,發現CsAP1-A在花中的表達量最高,在其他器官的表達量較低,說明CsAP1-A與花的發育和形成有關。在墨蘭花芽分化的5 個時期中,CsAP1-A在花芽分化初期(S1)的表達量最高,然后逐漸下降,在開花期(S5)的表達量降到最低,這與大部分的A 類基因在不同花發育時期表達規律相似,如山茶的CjAPL1[25]、麻風樹的JcAP1[22]和洋桔梗的EgAP1[26],因為S1 期是花器官形成的起始階段和關鍵階段,AP1作為一個轉錄因子,可以在此階段啟動信號來激活其他基因促進花的形成,而隨著花芽的不斷發育,其他相互作用的調控因子和信號通路逐漸介入,AP1的表達量就會逐漸降低。在墨蘭花朵的不同組織部位中,CsAP1-A在合蕊柱的表達量最高,其次是唇瓣,推測CsAP1-A主要參與墨蘭合蕊柱的發育。對墨蘭不同花型進行轉錄組測序分析,發現CsAP1-A在WT、MPV、LaPV 和NLV 4 種花型的合蕊柱中表達量均最高,這與RT-qPCR 檢測WT 花型中CsAP1-A的表達結果相一致。AP1作為A 類基因主要在第一、二輪花器官中表達,誘導花瓣和萼片的發育[7];但在蘭科植物中,AP1不在第一、二輪花器官表達,而是在內輪的合蕊柱中表達[27]。
根據蛋白互作關系的預測結果可知,CsAP1-A 大多與MADS-box 蛋白家族存在相互作用,而這些蛋白均與花器官發育和花期相關,因此預測CsAP1-A 蛋白在墨蘭花器官發育過程中起重要作用,這為進一步研究墨蘭花器官發育分子調控機制提供理論依據。
4 結論
本研究在墨蘭花芽中克隆得到CsAP1-A基因,該基因編碼248 個氨基酸,含有MADS-box轉錄因子家族的MADS-box 和K-box 結構域。墨蘭CsAP1-A與春蘭和蕙蘭的親緣關系最近,且可能與MADS1、MADS6、MADS47、MADS8 等蛋白具有相互作用。CsAP1-A在墨蘭花中的相對表達量顯著高于其他器官,且在花芽分化初期的表達量最高,并隨著花朵發育而逐漸下降;在墨蘭花朵不同組織部位中,CsAP1-A在合蕊柱的表達量最高,其次是唇瓣、花瓣和萼片。通過對不同花型的墨蘭品種轉錄組測序發現,CsAP1-A在WT、MPV、LaPV 和NLV 4 種花型的合蕊柱中表達量均最高。綜上所述,CsAP1-A可能在墨蘭花發育和合蕊柱形成中具有重要作用。
