NF-κB 和Nrf2 信號(hào)通路在成肌細(xì)胞抗氧化應(yīng)激中的作用
2023-11-14 15:50:04王鶴潔李俊玲竇敏敏翟竹匯李振月
山西農(nóng)業(yè)科學(xué) 2023年11期
關(guān)鍵詞:氧化應(yīng)激信號(hào)
李 君 ,王鶴潔 ,安 潔 ,李俊玲 ,竇敏敏 ,翟竹匯 ,何 浪 ,李振月 ,秦 健 ,,3,杜 榮
(1.山西農(nóng)業(yè)大學(xué) 動(dòng)物醫(yī)學(xué)學(xué)院,山西 太谷 030801;2.山西農(nóng)業(yè)大學(xué) 生命科學(xué)學(xué)院,山西 太谷 030801;3.山西農(nóng)業(yè)大學(xué) 實(shí)驗(yàn)教學(xué)中心,山西 太谷 030801)
氧化應(yīng)激最初由SOHAL 教授提出,指的是在機(jī)體受到各種內(nèi)外環(huán)境的有害刺激時(shí),體內(nèi)會(huì)產(chǎn)生過(guò)多的高活性分子如活性氧自由基(Reactive oxygen species,ROS)和活性氮自由基(Reactive nitrogen species,RNS),而細(xì)胞內(nèi)的抗氧化系統(tǒng)不能及時(shí)清理,會(huì)導(dǎo)致氧化與抗氧化系統(tǒng)的失衡,使細(xì)胞膜和線粒體以及遺傳物質(zhì)受到原發(fā)或繼發(fā)性的損傷,嚴(yán)重情況下可能導(dǎo)致細(xì)胞發(fā)生癌變或者死亡[1-2]。研究發(fā)現(xiàn),核因子κB(NF-κB)和核因子E2 相關(guān)因子2(Nrf2)是被過(guò)量ROS 激活的2 個(gè)主要轉(zhuǎn)錄因子,活化后的轉(zhuǎn)錄因子進(jìn)入細(xì)胞核,引起下游各種靶基因的表達(dá)以抵抗不良影響[3]。在NF-κB 信號(hào)通路的多個(gè)靶點(diǎn)中,SOD2和FHC在抗氧化活性方面有著重要的作用,其中NF-κB 可通過(guò)FHC的上調(diào)來(lái)抑制ROS,進(jìn)而抑制腫瘤壞死因子誘導(dǎo)的細(xì)胞凋亡[4];而CHANG 等[5]研究發(fā)現(xiàn),殼寡糖能夠激活Nrf2 途徑,促進(jìn)Nrf2 介導(dǎo)的HO-1和GSH-Px基因表達(dá),從而減少ROS 水平,提高肉雞的肉質(zhì)。
骨骼肌約占動(dòng)物身體的40%,其具有維持運(yùn)動(dòng)、代謝和支持身體的重要作用[6]。骨骼肌發(fā)育的過(guò)程較為復(fù)雜,胚胎和圍產(chǎn)期是決定動(dòng)物骨骼肌生長(zhǎng)性能的關(guān)鍵時(shí)期[7]。現(xiàn)已明確地塞米松作為一種人工合成的糖皮質(zhì)激素,因其在圍產(chǎn)期有抗炎、鎮(zhèn)痛、抗毒、抗過(guò)敏等多種優(yōu)勢(shì)被廣泛使用。但由于糖皮質(zhì)激素受體幾乎存在于所有的細(xì)胞中,高劑量或長(zhǎng)期使用會(huì)產(chǎn)生一定的副作用[8]。研究表明,地塞米松能夠在體內(nèi)和體外的骨骼肌細(xì)胞、成骨細(xì)胞以及脂肪細(xì)胞等多種類型的細(xì)胞中引起氧化應(yīng)激損傷[9-10]。氧化應(yīng)激會(huì)使蛋白質(zhì)代謝加快而抑制蛋白質(zhì)的合成,最終引起肌肉發(fā)育不良或肌肉萎縮[11]。目前,利用分離培養(yǎng)的成肌細(xì)胞誘導(dǎo)分化已被廣泛應(yīng)用于研究肌肉生成機(jī)制的體外模型[12-13]。
為探明在成肌分化過(guò)程中,NF-κB 和Nrf2 信號(hào)通路在地塞米松誘導(dǎo)的氧化應(yīng)激中是否發(fā)揮作用,本試驗(yàn)以綿羊成肌細(xì)胞為研究對(duì)象,在其分化過(guò)程中,選用地塞米松來(lái)誘導(dǎo)氧化應(yīng)激,比較正常和應(yīng)激狀態(tài)下不同分化階段NF-κB 和Nrf2 信號(hào)通路相關(guān)基因的表達(dá)變化,并分析各基因的相互關(guān)系,旨在為闡明骨骼肌細(xì)胞分化過(guò)程中的抗氧化機(jī)制奠定基礎(chǔ),同時(shí)為防治胎兒骨骼肌發(fā)育過(guò)程中受到的氧化損傷提供理論依據(jù)。
1 材料和方法
1.1 試驗(yàn)材料
綿羊胎兒成肌細(xì)胞由本實(shí)驗(yàn)分離培養(yǎng)所得。主要試劑包括青霉素-鏈霉素(PS)、I 型膠原酶、0.25%胰酶(TE)、馬血清(HS)、地塞米松,均購(gòu)于索萊寶;DMEM/F12、NBCS 購(gòu)于GIBCO 公司;RNAiso Plus 購(gòu)于TaKaRa 公司;活性氧檢測(cè)試劑盒購(gòu)于碧云天公司。
1.2 試驗(yàn)方法
1.2.1 綿羊胎兒成肌細(xì)胞的分離與培養(yǎng) 樣品來(lái)自于太谷屠宰場(chǎng)屠宰時(shí)意外發(fā)現(xiàn)的早期懷孕母羊。將胎兒從子宮中取出,用加了雙抗的生理鹽水沖洗表面,無(wú)菌手術(shù)刀切成大小為1 mm3的組織碎片,以1 500 r/min 的轉(zhuǎn)速離心5 min。然后,用0.2%的I 型膠原酶消化1 h,中間每10 min 混勻一次,消化結(jié)束后,再次離心。接著用0.25%胰酶消化20 min,加等量培養(yǎng)基(89% DMEM/F12、10% NBCS、1% PS)終止消化,用0.074 mm 細(xì)胞篩進(jìn)行過(guò)濾,離心,懸浮培養(yǎng)在100 mm 細(xì)胞培養(yǎng)皿中,隨后通過(guò)差速貼壁的方法純化成肌細(xì)胞。
1.2.2 綿羊胎兒成肌細(xì)胞的分化與處理 綿羊胎兒成肌細(xì)胞長(zhǎng)到80%左右時(shí),對(duì)照組用含有2%HS和1% PS的DMEM/F12正常誘導(dǎo)分化48、72 h。根據(jù)山西農(nóng)業(yè)大學(xué)分子細(xì)胞調(diào)控課題組前期經(jīng)驗(yàn),在低濃度時(shí),地塞米松對(duì)成肌分化有促進(jìn)作用,中高濃度時(shí)對(duì)成肌分化有抑制作用。因此,本試驗(yàn)中試驗(yàn)組的處理方式為:在正常分化的細(xì)胞中添加中濃度(0.375 mg/mL)的地塞米松,分別處理細(xì)胞48、72 h。
1.2.3 ROS 檢測(cè) 采用活性氧檢測(cè)試劑盒檢測(cè)各組成肌細(xì)胞中的ROS 水平。每孔加入4%多聚甲醛溶液固定2 min,PBS 洗滌3 次。然后加入ROS檢測(cè)工作液,置培養(yǎng)箱20 min,洗滌細(xì)胞3 次,熒光顯微鏡觀察并分析ROS 水平。1.2.4 RNA-seq 分析 分別將不加地塞米松分化48、72 h 和加地塞米松分化48、72 h 的細(xì)胞樣品收集放置于RNAiso Plus 裂解液中,送至百邁客公司進(jìn)行轉(zhuǎn)錄組測(cè)序分析。根據(jù)測(cè)序結(jié)果,分析NF-κB和Nrf2 信號(hào)通路相關(guān)基因的mRNA 相對(duì)表達(dá)情況。
1.3 生物信息學(xué)及統(tǒng)計(jì)學(xué)分析
采用皮爾森相關(guān)系數(shù)法分析相關(guān)性,采用SPSS 22.0 單因素(ANOVA)方差分析差異顯著性;同時(shí)利用STRING 數(shù)據(jù)庫(kù)分析并建立蛋白互作網(wǎng)絡(luò)。每個(gè)處理3 個(gè)重復(fù)。
2 結(jié)果與分析
2.1 綿羊胎兒成肌細(xì)胞分離、分化及地塞米松處理后的形態(tài)和ROS 變化
分離培養(yǎng)的綿羊成肌細(xì)胞在2~3 d 內(nèi)開始從組織塊爬出,并在第3 天時(shí)開始大量增殖,如圖1 所示,細(xì)胞形態(tài)正常,呈長(zhǎng)梭形或者梭形,可用作后續(xù)試驗(yàn)。誘導(dǎo)分化后,單核成肌細(xì)胞開始變長(zhǎng)并逐漸融合為多核肌管。
圖1 綿羊胎兒成肌細(xì)胞在培養(yǎng)基中的生長(zhǎng)狀態(tài)Fig.1 Growth status of sheep fetal myoblasts in the culture medium
地塞米松處理前后的變化如圖2 所示,隨著分化時(shí)間延長(zhǎng)肌管數(shù)量增加,地塞米松處理抑制了成肌細(xì)胞的分化,導(dǎo)致形成的肌管變短變細(xì)、數(shù)量減少。相應(yīng)地,與N 48 h 組相比,Dex 48 h 組中ROS含量增加了130.96%,差異極顯著(P<0.01);與N 72 h 組相比,Dex 72 h 組中ROS 含量增加了151.57%,差異極顯著(P<0.01)。

圖2 綿羊胎兒成肌細(xì)胞在加或不加Dex 的分化培養(yǎng)基中分化的狀態(tài)(A)和ROS 水平(B)Fig.2 Differentiation status(A),ROS levels(B)of sheep fetal myoblasts in differentiation medium with or without Dex
2.2 NF-κB 和Nrf2 通路相關(guān)基因的mRNA 表達(dá)結(jié)果
由測(cè)序結(jié)果分析可知(圖3),NF-κB 信號(hào)通路中,與N 48 h 組相比,Dex 48 h 組中P65、IKK2、IκBα、IκBβ、SOD2和FHC的mRNA 相對(duì)表達(dá)量分別上升了53.68%、36.38%、219.36%、55.64%、415.32% 和388.12%,差異極顯著(P<0.01);與N 72 h組相比,Dex 72 h組中P65、IKK2、IκBα、SOD2和FHC的mRNA 相對(duì)表達(dá)量分別上升了45.39%、66.85%、226.32%、436.08%和339.08%,差異極顯著(P<0.01),IκBβ的mRNA 相對(duì)表達(dá)量上升了47.58%,差異達(dá)顯著水平(P<0.05)。
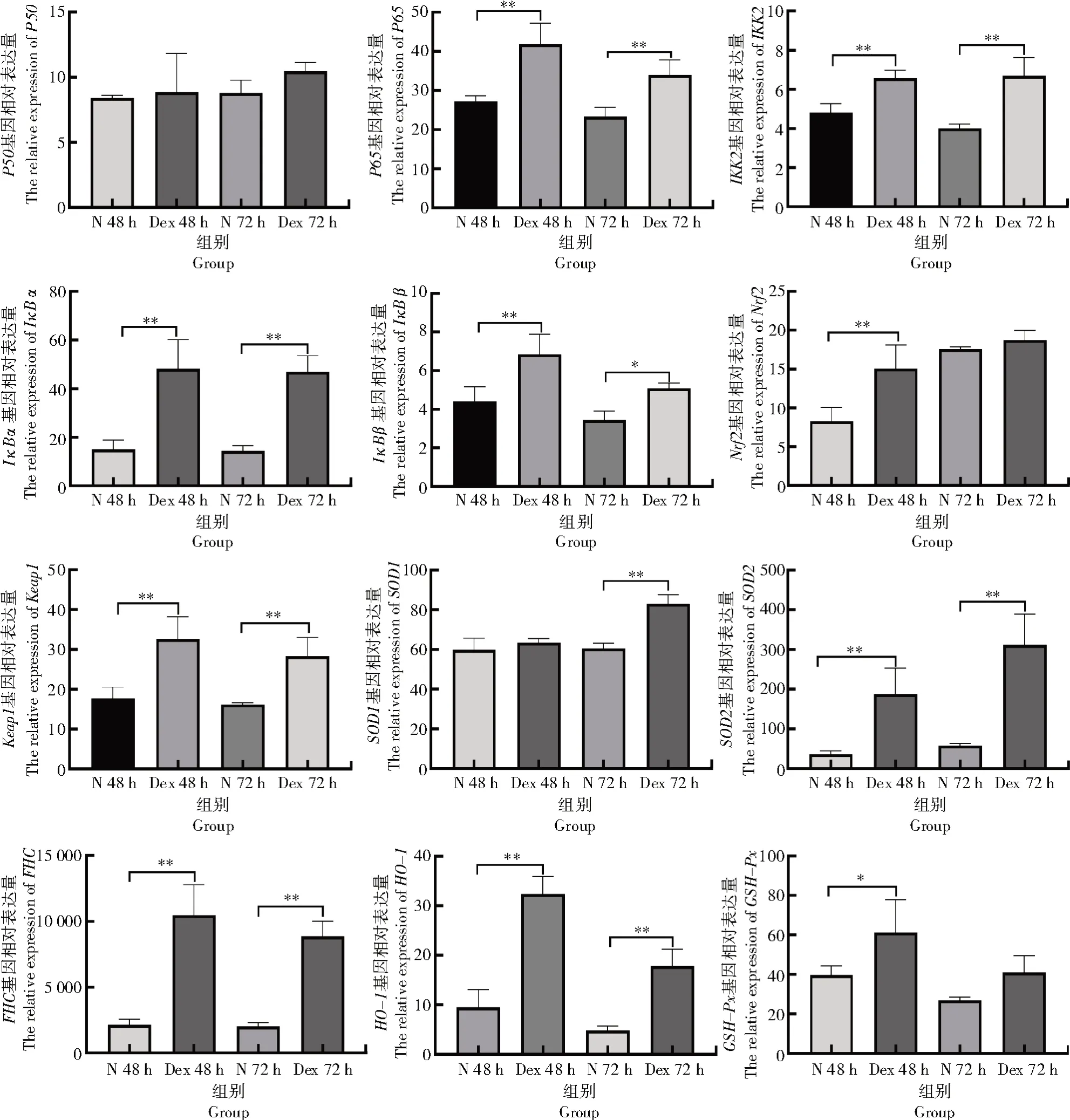
圖3 NF-κB 和Nrf2 信號(hào)通路相關(guān)基因的相對(duì)表達(dá)量Fig.3 Relative expression level of genes related to NF-κB 和Nrf2 signaling pathways
Nrf2 信號(hào)通路中,與N 48 h 組相比,Dex 48 h組中Keap1、Nrf2和HO-1的mRNA 相對(duì)表達(dá)量分別上升了83.98%、81.46%和242.02%,差異極顯著(P<0.01),GSH-Px的mRNA 相對(duì)表達(dá)量上升了54.45%,差異顯著(P<0.05)。與N 72 h 組相比,Dex 72 h 組中Keap1、SOD1和HO-1的mRNA 相對(duì)表達(dá)量分別上升了74.78%、37.20%和270.51%,差異達(dá)極顯著水平(P<0.01)。
2.3 NF-κB 和Nrf2 通路相關(guān)基因mRNA 表達(dá)的相關(guān)性
由表1 可知,分化48 h 后,NF-κB 信號(hào)通路中,P65與IKK2、IκBα、IκBβ和FHC呈極顯著正相關(guān)(P<0.01);IKK2與IκBα呈極顯著正相關(guān)(P<0.01),與IκBβ、SOD2和FHC呈顯著正相關(guān)(P<0.05);IκBα與IκBβ和FHC呈極顯著正相關(guān)(P<0.01);IκBβ與FHC呈顯著正相關(guān)(P<0.05)。Nrf2信號(hào)通路中,Nrf2與SOD2呈極顯著正相關(guān)(P<0.01);HO-1與Keap1呈極顯著正相關(guān)(P<0.01)。NF-κB 信號(hào)通路和Nrf2 信號(hào)通路相關(guān)性分析結(jié)果顯示,Keap1與P65、IKK2、IκBα、IκBβ和FHC呈極顯著正相關(guān)(P<0.01);HO-1與P65、IKK2、IκBα和FHC呈極顯著正相關(guān)(P<0.01),與IκBβ呈顯著正相關(guān)(P<0.05);GSH-Px與IκBβ和FHC呈顯著正相關(guān)(P<0.05)。
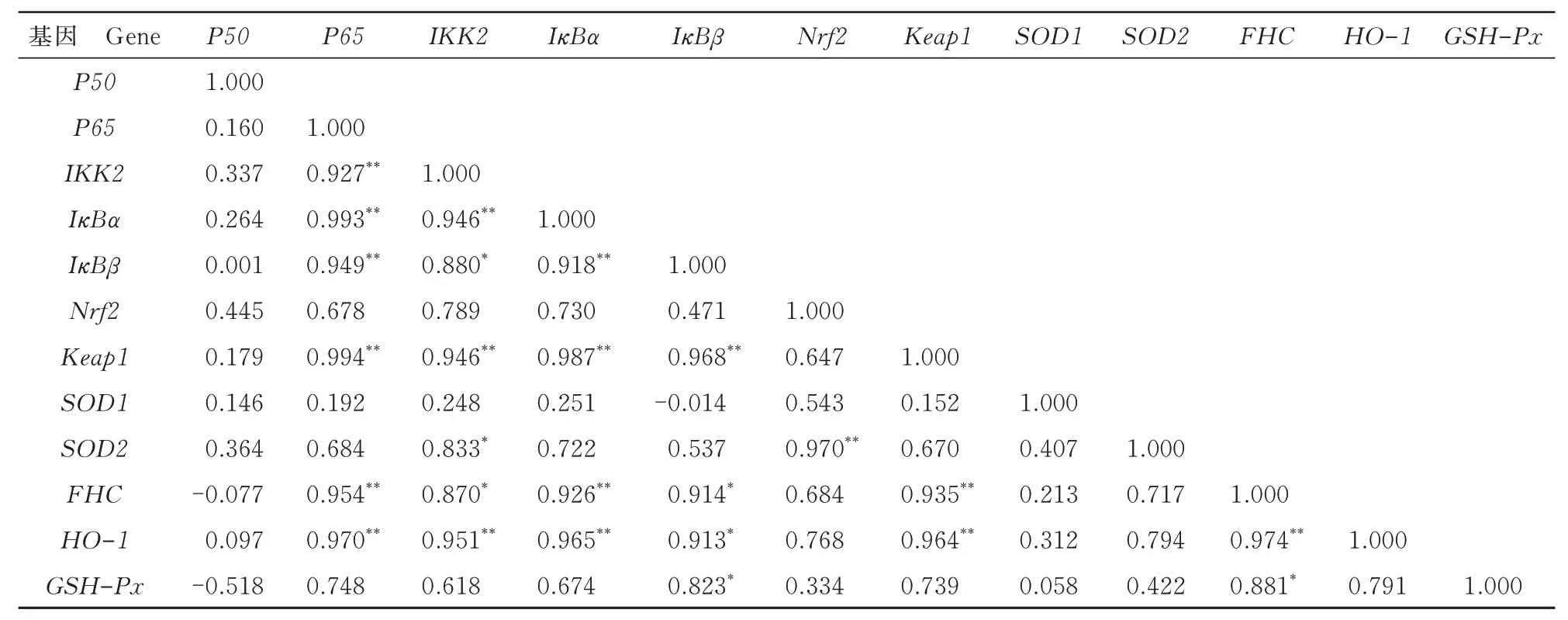
表1 分化48 h 后各組相關(guān)基因mRNA 表達(dá)的相關(guān)性Tab.1 Correlation of mRNA expression of the related genes in each group after 48 h of differentiation
由表2 可知,分化72 h 后,NF-κB 信號(hào)通路中,P50與IκBβ呈顯著正相關(guān)(P<0.05);P65與IKK2、IκBα、IκBβ和FHC呈極顯著正相關(guān)(P<0.01);IKK2與IκBα和FHC呈極顯著正相關(guān)(P<0.01),與IκBβ呈顯著正相關(guān)(P<0.05);IκBα與IκBβ和FHC呈極顯著正相關(guān)(P<0.01),與SOD2呈顯著正相關(guān)(P<0.05);IκBβ與FHC呈極顯著正相關(guān)(P<0.01);SOD2與FHC呈顯著正相關(guān)(P<0.05)。Nrf2 信號(hào)通路中,Keap1與HO-1和GSH-Px呈極顯著正相關(guān)(P<0.01),與SOD1呈顯著正相關(guān)(P<0.05);SOD1與HO-1呈極顯著正相關(guān)(P<0.01),與SOD2呈顯著正相關(guān)(P<0.05);SOD2與HO-1呈顯著正相關(guān)(P<0.05);HO-1與GSH-Px呈極顯著正相關(guān)(P<0.01)。Nrf2 信號(hào)通路和NF-κB 信號(hào)通路相關(guān)性分析結(jié)果顯示,Keap1與P65、IKK2、IκBα、IκBβ和FHC呈極顯著正相關(guān)(P<0.01);SOD1與P50呈極顯著正相關(guān)(P<0.01),與IKK2、IκBα、IκBβ和FHC呈顯著正相關(guān)(P<0.05);HO-1與P65、IKK2、IκBα、IκBβ和FHC呈極顯著正相關(guān)(P<0.01),與P50呈顯著正相關(guān)(P<0.05);GSH-Px與P65、IKK2和IκBα呈極顯著正相關(guān)(P<0.01),與IκBβ和FHC呈顯著正相關(guān)(P<0.05)。
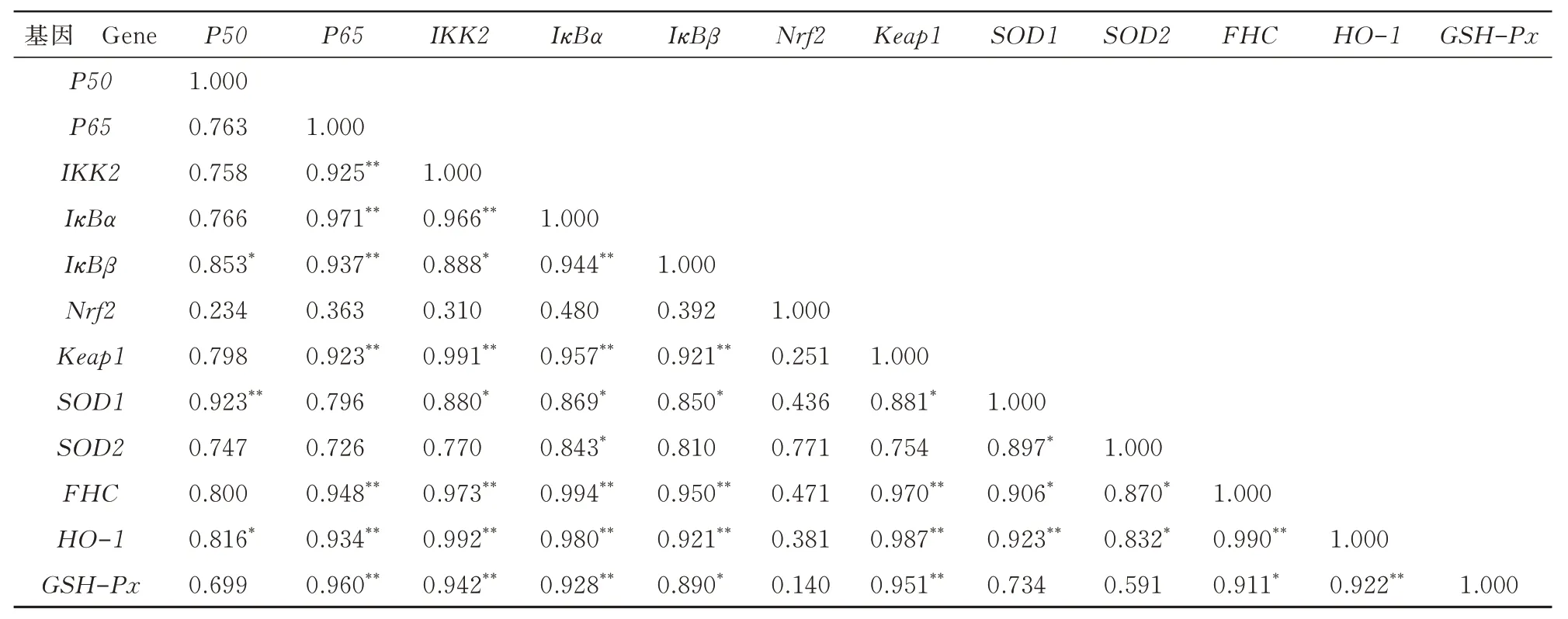
表2 分化72 h 后各組相關(guān)基因mRNA 表達(dá)的相關(guān)性Tab.2 Correlation of mRNA expression of the related genes in each group after 72 h of differentiation
2.4 NF-κB 和Nrf2 通路相關(guān)基因的蛋白互作分析結(jié)果
為了進(jìn)一步了解NF-κB 和Nrf2 這2 條通路中氧化應(yīng)激相關(guān)基因之間的關(guān)系,利用STRING 數(shù)據(jù)庫(kù)分析并建立了蛋白互作網(wǎng)絡(luò),結(jié)果顯示(圖4),NF-κB 和Nrf2 這2 條通路的多數(shù)基因之間存在互作關(guān)系,而且2 條通路之間具有交叉對(duì)話作用,其中,Keap1是聯(lián)系NF-κB 和Nrf2 這2 條通路的關(guān)鍵樞紐,Nrf2(NFE2L2)和IκBα(NFκBIA)之間的互作關(guān)系也是2 條通路溝通的方式之一,而SOD2是2 條通路共同調(diào)控的下游基因。

圖4 NF-κB 與Nrf2 通路相關(guān)基因的蛋白互作網(wǎng)絡(luò)Fig.4 Protein-protein interaction(PPI)network of genes related to NF-κB and Nrf2 pathways
3 結(jié)論與討論
細(xì)胞的功能會(huì)受到細(xì)胞內(nèi)在損傷和外界刺激的影響,包括氧化應(yīng)激、DNA 損傷、蛋白質(zhì)損傷和新陳代謝改變等[13]。在骨骼肌發(fā)育、再生以及體外研究成肌分化的過(guò)程中,不可避免地會(huì)受到氧化應(yīng)激影響,適當(dāng)?shù)腞OS 能夠參與肌衛(wèi)星細(xì)胞的激活以及成肌細(xì)胞的增殖分化,但過(guò)量增加的ROS 會(huì)導(dǎo)致細(xì)胞功能障礙,影響骨骼肌細(xì)胞的發(fā)育和再生[14]。研究表明,地塞米松可以引起小鼠成肌細(xì)胞氧化應(yīng)激[15]。本研究中,通過(guò)形態(tài)學(xué)觀察與ROS檢測(cè)結(jié)果發(fā)現(xiàn),與正常對(duì)照組相比,地塞米松處理導(dǎo)致分化48 h 和72 h 的綿羊成肌細(xì)胞內(nèi)ROS 含量顯著上升,成肌細(xì)胞出現(xiàn)受損,分化和融合成肌管的數(shù)目減少,肌管的直徑變小,表明成肌分化過(guò)程中的氧化應(yīng)激模型成功構(gòu)建。其原因可能是由于地塞米松引起的氧化應(yīng)激會(huì)抑制成肌細(xì)胞內(nèi)蛋白質(zhì)的合成,促進(jìn)肌管內(nèi)蛋白質(zhì)的降解[16],從而影響成肌細(xì)胞的分化及融合過(guò)程。
NF-κB 蛋白家族作為一種多效性的轉(zhuǎn)錄因子,在調(diào)節(jié)細(xì)胞凋亡、炎癥反應(yīng)、應(yīng)激反應(yīng)、免疫應(yīng)答中發(fā)揮著重要的生物學(xué)功能[17-18]。該家族中P65 和P50 構(gòu)成的異二聚體屬于典型的誘導(dǎo)型[19],正常情況下,其與抑制蛋白IκB 形成三聚體存在于細(xì)胞質(zhì)中。當(dāng)受到刺激后,會(huì)激活I(lǐng)κB 激酶的亞型IKK2,使IκB 發(fā)生降解,P65 和P50 得以進(jìn)入核內(nèi)啟動(dòng)SOD2和FHC等下游基因的表達(dá)[20]。作為一種超氧化物清除的關(guān)鍵酶,SOD2 的主要作用是清除氧化磷酸化產(chǎn)物超氧陰離子自由基[21]。FHC編碼的鐵蛋白可以將細(xì)胞內(nèi)的鐵儲(chǔ)存起來(lái)以限制芬頓反應(yīng)的發(fā)生,使細(xì)胞中的ROS 含量減少[22]。本試驗(yàn)中,Dex 48 h 組和Dex 72 h 組分別與對(duì)照相比發(fā)現(xiàn),NF-κB 通路中IKK2、P65、IκBα、IκBβ、SOD2、FHC的mRNA 表達(dá)水平均呈極顯著升高,表明ROS 激活了NF-κB 通路,繼而促進(jìn)了下游靶基因SOD2和FHC的表達(dá),其中部分原因是由IKK2、P65基因表達(dá)量的升高所致。同時(shí),抑制因子IκBmRNA 的表達(dá)也出現(xiàn)了顯著上升,其原因可能是IκB也屬于NF-κB 的下游靶基因,該轉(zhuǎn)錄因子的活化或表達(dá)增加會(huì)直接調(diào)控IκB 的表達(dá),使其表達(dá)上調(diào)從而反饋性抑制NF-κB 的過(guò)度活化[23-24]。通過(guò)對(duì)各基因之間mRNA 表達(dá)的相關(guān)性分析結(jié)果可知,分化48、72 h 時(shí)NF-κB 通路相關(guān)基因隨地塞米松處理表現(xiàn)出的表達(dá)相關(guān)性存在一致性也存在差異性。2 個(gè)階段中P65與IKK2、IκBα、IκBβ和FHC均存在顯著相關(guān)性,而IKK2、IκBα、IκBβ和FHC相互之間多數(shù)存在顯著相關(guān),但2 個(gè)階段有所差異,此外,2 個(gè)階段中與SOD2呈現(xiàn)顯著相關(guān)的基因有所不同。
Nrf2 通路是抗氧化應(yīng)激的主要機(jī)制之一。YANG 等[25]研究發(fā)現(xiàn),香葉素能夠通過(guò)激活Nrf2/HO-1 通路來(lái)減輕氧化應(yīng)激,從而預(yù)防腦缺血/再灌注損傷。李孟心等[26]證明了Nrf2/HO-1 信號(hào)通路在H2O2誘導(dǎo)的hiPSCs 氧化應(yīng)激中發(fā)揮了明顯的抗氧化作用。正常情況下,細(xì)胞內(nèi)的Nrf2 與Keap1 結(jié)合,以非活性的形式存在。當(dāng)發(fā)生氧化應(yīng)激后Nrf2與Keap1 發(fā)生解離并轉(zhuǎn)移至細(xì)胞核內(nèi),與Maf 蛋白形成異二聚體后,通過(guò)與250 多個(gè)基因啟動(dòng)子區(qū)域中存在的抗氧化反應(yīng)元件(ARE)序列結(jié)合,來(lái)啟動(dòng)一系列抗氧化基因的轉(zhuǎn)錄,其中包括HO-1、SOD和GSH-Px等[27-28]。本試驗(yàn)研究表明,與對(duì)照相比,處理組HO-1、SOD1、SOD2和GSH-PxmRNA的表達(dá)量多數(shù)達(dá)到顯著上升,至少部分是由于Nrf2表達(dá)上調(diào)介導(dǎo)的。HO-1、SOD、GSH-Px表達(dá)增加后,分別主要通過(guò)阻止血紅素基團(tuán)促氧化[29]、清除氧化磷酸化產(chǎn)物超氧陰離子自由基[21]、分解有毒的過(guò)氧化物[30-31]等作用而減緩ROS 對(duì)綿羊成肌細(xì)胞分化的不利影響。同時(shí),抑制因子Keap1mRNA的表達(dá)也出現(xiàn)了顯著上升,這與上述NF-κB 通路中抑制因子IκBmRNA 表達(dá)上升的機(jī)制可能是類似的,是機(jī)體的一種負(fù)反饋調(diào)節(jié)。通過(guò)對(duì)各基因之間mRNA 表達(dá)的相關(guān)性分析結(jié)果可知,分化48、72 h時(shí)Nrf2 通路相關(guān)基因隨地塞米松處理表現(xiàn)出的表達(dá)相關(guān)性同樣存在異同。其中,Keap1、SOD1、SOD2、HO-1、GSH-Px之間,多數(shù)基因在48 h 或(和)72 h 表現(xiàn)出相關(guān)性,而Nrf2的表達(dá)與SOD2存在相關(guān)性。當(dāng)細(xì)胞受到氧化應(yīng)激時(shí),多種信號(hào)通路被激活參與抗氧化應(yīng)激,這些通路之間也存在相互作用。
本試驗(yàn)進(jìn)一步對(duì)NF-κB 和Nrf2 這2 條通路基因的表達(dá)相關(guān)性以及蛋白互作關(guān)系進(jìn)行分析可知,盡管有些基因之間的表達(dá)相關(guān)性不顯著,但這2 條通路各基因之間存在調(diào)控關(guān)系。因?yàn)榛虻膍RNA 轉(zhuǎn)錄往往同時(shí)受到多個(gè)相關(guān)轉(zhuǎn)錄因子的調(diào)控以及其他因素的影響。李慧[32]研究發(fā)現(xiàn),NF-κB的抑制蛋白IκB 可以抑制NF-κB 通路并活化Nrf2通路,表明細(xì)胞內(nèi)這2 條通路之間存在相互作用,這與本研究結(jié)果基本一致。
SOD2是2 條通路共同調(diào)控的下游基因。本試驗(yàn)中,SOD2基因可能同時(shí)受到轉(zhuǎn)錄因子Nrf2和P65的調(diào)控,而且受到其他相關(guān)基因的影響,因此它的表達(dá)水平是受到各因子調(diào)控或影響的綜合結(jié)果。Nrf2 通路中的關(guān)鍵抑制因子Keap1 是一種上游蛋白,參與不同信號(hào)通路的協(xié)調(diào),在Nrf2 與NFκB 信號(hào)通路之間發(fā)揮著重要的作用[33]。
綜上可見,無(wú)論是分化48 h 還是72 h 時(shí),Keap1與NF-κB 通路相關(guān)基因P65、IKK2、IκBα、IκBβ和FHC的相關(guān)性均為顯著。結(jié)合蛋白互作關(guān)系進(jìn)一步表明,Keap1基因是聯(lián)系NF-κB 和Nrf2 這2 條通路的關(guān)鍵樞紐。說(shuō)明在綿羊成肌細(xì)胞分化過(guò)程中,NF-κB 和Nrf2 信號(hào)通路被激活,其上游基因P65、IKK2、IκBα、IκBβ、Nrf2、Keap1以及相應(yīng)下游靶基因SOD1、SOD2、FHC、HO-1、GSH-Px的mRNA相對(duì)表達(dá)量均顯著升高,2 條通路以Keap1因子為重要樞紐而交互聯(lián)系,共同在地塞米松誘導(dǎo)的氧化應(yīng)激中發(fā)揮抗氧化作用,從而降低細(xì)胞的氧化應(yīng)激損傷,減緩氧化應(yīng)激對(duì)成肌分化造成的影響。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
世界科學(xué)技術(shù)-中醫(yī)藥現(xiàn)代化(2020年2期)2020-07-25 02:05:56
中國(guó)生殖健康(2019年3期)2019-02-01 06:12:26
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
西南軍醫(yī)(2016年6期)2016-01-23 02:21:19
新疆醫(yī)科大學(xué)學(xué)報(bào)(2015年10期)2015-12-26 12:33:30
海軍航空大學(xué)學(xué)報(bào)(2015年3期)2015-11-11 17:20:00
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
西南軍醫(yī)(2015年2期)2015-01-22 09:09:37
