3-羥基丁酸對雨生紅球藻PSⅡ光化學活性與色素合成的影響
2023-11-20 07:34:08黃瑩張娟竇勇原雪峰翟勝利曲木周文禮
食品研究與開發 2023年22期
黃瑩,張娟,竇勇*,原雪峰,翟勝利,曲木,周文禮
(1.天津農學院水產學院,天津市水產生態及養殖重點試驗室,天津 300384;2.天津現代天驕農業科技股份有限公司,天津市綠色生態飼料重點實驗室,天津 301801;3.天津現代晨輝科技集團,天津 301802)
雨生紅球藻(Haematococcus pluvialis)是一種淡水單細胞藻類,隸屬于綠藻門、綠藻綱、團藻目、紅球藻科、紅球藻屬,普遍分布于自然界的淡水生境中[1]。在外界環境條件適宜時,微藻以游動狀態存在,此時細胞呈卵圓形或橢圓形,依靠兩條頂生、等長的鞭毛運動,這一階段的細胞大多呈綠色,營養生長旺盛,生物量積累迅速。在受到環境脅迫時,雨生紅球藻細胞以紅色厚壁孢子形式存在,細胞鞭毛脫落,運動能力喪失,微藻細胞逐漸合成并積累蝦青素,此階段藻細胞內積累大量蝦青素,雨生紅球藻細胞內的蝦青素含量可以占到細胞干重的1.5%~4.0%[2-3],而且全部是高生物活性的3S、3S'形態,因此,雨生紅球藻被公認為自然界中生產天然蝦青素的最佳“生物反應器”。蝦青素是一種脂溶性的次級類胡蘿卜素,屬于菇烯類的不飽和化合物,蝦青素是目前為止在自然界有機體中發現的抗氧化能力最強的物質[4],其清除自由基和單線態氧淬滅的能力遠遠高于維生素E,比玉米黃質、番茄紅素及β-胡蘿卜素等常見抗氧化物質的抗氧化能力也要高10 倍以上。因其具有清除自由基、提高機體免疫力的作用,目前多被用作保健品輔料和食品添加劑[5-7]。蝦青素的功能不僅如此,其還有良好的著色效果,可以進入生物體并貯存在組織中,例如農業農村部2003 年318 號公告中指定天然蝦青素為水產動物唯一著色劑[8-9]。雨生紅球藻不動孢子內積累大量蝦青素,且均為具有高生物活性的3S、3S'異構體,目前被公認為自然界中生產天然蝦青素的最佳生物[10]。
然而自然條件下雨生紅球藻合成蝦青素的效率和對底物、能源的利用率都很低,基本無法支撐蝦青素的工業化生產,因此使用外源化學物質進行人工誘導成為提高雨生紅球藻生產蝦青素能力的常用方法[11-17]。3-羥基丁酸(3-hydroxybutyric acid,3-HB)通常被認為是生物體內關鍵的能量代謝中間產物之一[18],它是分子量較小的高分子有機化合物,具有光學活性,純態時可結晶,羥基基團能締合,易溶于水、乙醇、乙醚、干蒸易分解等特點。目前,已有研究證實3-羥基丁酸具有一系列重要的生理活性,例如可以促進神經膠質細胞和成骨細胞生長,增強顆粒細胞和半顆粒細胞的體外吞噬作用,提高血細胞內溶菌(lysozyme,LZM)和血清酸性磷酸酶(acid phosphatase,ACP)的活性[19],除此外在臨床醫療領域,3-羥基丁酸及其聚合物已被用來治療出血性休克等多種疾病[20]。雖然3-羥基丁酸具備許多生理功能且具有重要的臨床醫療價值,但是在誘導微藻積累次級代謝產物方面尚未得到應用,因此利用3-羥基丁酸研究雨生紅球藻是一個創新性的試驗,通過研究它對雨生紅球藻生長和蝦青素積累的影響可以更進一步地尋找雨生紅球藻更優的培養條件。本研究主要探索3-羥基丁酸對雨生紅球藻光系統Ⅱ(pthotosystem Ⅱ,PSⅡ) 光化學活性與色素合成的影響,以期為藻源蝦青素的生產提供一定依據。
1 材料與方法
1.1 材料與試劑
雨生紅球藻(H.pluvialis):天津市水產生態及養殖重點試驗室;3-羥基丁酸標準品(3CAS 300-85-6,純度≥98%):美國Sigma-Aldrich 公司;BBM 培養基配方試劑(均為分析純):天津市風船化學試劑科技有限公司。
1.2 儀器與設備
調制脈沖熒光儀(IMAGING-PAM):德國WALZ有限公司;紫外-可見分光光度計(TU-1810):北京普析通用儀器有限責任公司;超聲波細胞破碎儀(JY92-Ⅱ):寧波新芝生物科技股份有限公司。
1.3 方法
1.3.1 試驗設計
以BBM 培養基對H.pluvialis 進行培養(配方見表1),設置光照強度為55 μmol/(m2·s),光暗比為12 h:12 h,培養溫度為(22±1)℃。每天搖動培養瓶6 次,防止細胞附壁或下沉。

表1 BBM 培養基配方Table 1 BBM medium formula
將培養至對數生長期的H.pluvialis 接種到250 mL三角瓶中,各試驗組微藻初始密度均為5×105cells/mL,培養體系體積控制在150 mL。試驗設置的3-羥基丁酸濃度分別為0、0.01、0.02、0.05、0.10 μg/L,每個試驗組均設置3 次平行。試驗周期為30 d,每3 d 取樣分析1 次。
1.3.2 H.pluvialis PSⅡ光化學活性測定
使用調制脈沖熒光儀測定PSⅡ光化學活性。向比色杯中依次加入3 mL 雙蒸水和15 μL 藻液,混勻,將樣品暗適應15 min,讀取最大光能轉化效率(Fv/Fm)、有效光能轉化效率(Yield)和表觀電子傳遞速率(electron transfer rate,ETR)的數值。
1.3.3 H.pluvialis 葉綠素a 含量測定
吸取2 mL 藻液置于離心管中,4 ℃下以12 000 r/min離心5 min,用移液槍吸去上清液,加入80%丙酮對藻泥進行再懸浮,然后用錫箔完全包裹離心管,在暗處置于55 ℃水浴中30 min,再于4 ℃下以12 000 r/min 離心5 min,將上清液轉移至10 mL 離心管中,并用80%丙酮定容至5 mL,使用紫外-可見分光光度計測定663 nm 處吸光值(A663),然后根據下列公式計算葉綠素a 含量(Y,mg/L)[21]。
1.3.4 H.pluvialis 蝦青素含量測定
取5 mL 藻液進行離心,去上清液,向沉淀加入5 mL雙蒸水,重復此過程2 次。加入5% KOH+30%甲醇混合液去除葉綠素。再次離心后加入5 滴乙酸降低pH值,收集藻泥后加入2 mL 丙酮,用細胞破碎儀處理10 min,然后用二甲基亞砜抽提至藻團呈白色。用紫外可見分光光度計測定抽提液在490 nm 下的吸光度A490[22]。蝦青素含量(X,mg/L)參照如下公式計算。
X=4.5×A490×VA×VB
式中:VA為二甲基亞砜體積,mL;VB為藻液體積,mL。
1.4 數據統計與分析
各試驗組數據均以平均值±標準差表示。使用SPSS 17.0 軟件進行單因素方差分析,并且采用Duncan 方法進行多重比較,顯著性水平P=0.05。
不對稱條件下的儲能虛擬同步發電機低電壓穿越控制技術//何安然,侯凱,王小紅,蔣應偉,劉建平,盧方舟//(10):122
2 結果與分析
2.1 3-羥基丁酸對H.pluvialis PSⅡ光化學活性的影響
2.1.1 3-羥基丁酸對H.pluvialis Fv/Fm的影響
3-羥基丁酸對H.pluvialis 最大光能轉化效率的影響如圖1 所示。
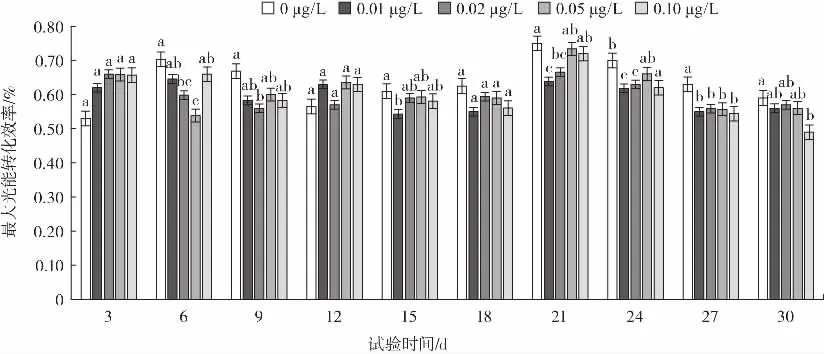
圖1 3-羥基丁酸對H.pluvialis Fv/Fm 的影響Fig.1 Effect of 3-hydroxybutyric acid on Fv/Fm in H.pluvialis
Fv/Fm為最大光能轉化效率,反映了植物PS II 的潛在最大光合能力,環境條件改變時微藻Fv/Fm會發生顯著變化。整體來看,3-羥基丁酸處理組的雨生紅球藻Fv/Fm從試驗開始至第18 天有緩慢下降趨勢,此后有一定程度上升,說明雨生紅球藻的PS II 反應中心經過一段時間后對3-羥基丁酸產生了一定的抗性,然而隨著時間的延長藻受到的損害更加嚴重,從第21 天開始Fv/Fm值又呈現逐漸下降趨勢。試驗期間,濃度分別為0.01、0.02、0.05、0.10 μg/L 的雨生紅球藻Fv/Fm較初始狀態分別降低了9.7%、13.6%、13.6%、25.7%,在大多數時間,3-羥基丁酸處理組的微藻Fv/Fm均顯著低于對照組的微藻Fv/Fm(P<0.05)。結果表明在缺氮條件下3-羥基丁酸處理組對雨生紅球藻加強了脅迫作用,并且在0.01 μg/L 的處理組時Fv/Fm值下降幅度最大。
2.1.2 3-羥基丁酸對H.pluvialis 實際光化學量子產量的影響
3-羥基丁酸對H.pluvialis 實際光能轉化效率的影響如圖2 所示。

圖2 3-羥基丁酸對H.pluvialis 實際光化學量子產量的影響Fig.2 Effect of 3-hydroxybutyric acid on Yield in H.pluvialis
Yeild 為實際光能轉化效率,反映植物PS II 在部分關閉情況下的實際原初光能捕獲效率。由圖2 可知,3-羥基丁酸處理組的Yield 參數在3 d 時,除0.02 μg/L組外,其他各3-羥基丁酸處理組的Yield 參數顯著低于對照組的參數(P<0.05);在6、9、21、24 d 時,0.10 μg/L 組的Yield 參數均低于對照組(P<0.05);在18 d 時,各處理組的Yield 參數均顯著低于對照組(P<0.05)。至試驗第27 天時,0.02 μg/L 和0.10 μg/L 濃度組的雨生紅球藻Yield 參數下降幅度最明顯,相比較初始階段分別降低了25%和16%,而其他濃度組的微藻Yield 數值下降幅度均未超過10%。說明3-羥基丁酸降低了雨生紅球藻PSⅡ的實際光合能力。
2.1.3 3-羥基丁酸對H.pluvialis 光合電子傳遞速率的影響

圖3 3-羥基丁酸對H.pluvialis ETR 的影響Fig.3 The effect of 3-hydroxybutyric acid on ETR in H.pluvialis
表觀光合電子傳遞速率(electrontransportrate,ETR)是反映植物PS II 反應中心活性與光合電子傳遞效率強弱的一個重要指標,其數值影響因素較多,主要與光強、植物吸收入射光和能量分布比例以及光子通量密度有關。由圖3 可知,3-羥基丁酸處理組的雨生紅球藻ETR 參數在第3 天時,除0.02 μg/L 組外,其余各處理組的雨生紅球藻ETR 均顯著低于對照組(P<0.05);在6、9 d 時,0.01μg/L 組和0.10 μg/L 組雨生紅球藻ETR 低于對照組;在24、27 d 時,0.1 μg/L 組的雨生紅球藻ETR顯著低于對照組;在第30天各3-羥基丁酸處理組的雨生紅球藻ETR 顯著低于對照組的ETR 參數值(P<0.05),說明3-羥基丁酸降低了雨生紅球藻PSⅡ的實際光和能力。直至試驗結束時,0.02 μg/L 組和0.10 μg/L 組的雨生紅球藻ETR 參數下降幅度最大,分別較初始階段降低了12.5%和32.9%,說明0.02 μg/L 和0.10 μg/L 濃度的3-羥基丁酸對于降低光合電子傳遞效率更加明顯。
2.2 3-羥基丁酸對H.pluvialis 色素合成的影響
3-羥基丁酸處理條件下,微藻資源在光合作用與抗逆合成之間權衡分配的情況,通過構建雨生紅球藻細胞內蝦青素與葉綠素a 含量比值這一指標來反映,如圖4 所示。

圖4 3-羥基丁酸對H.pluvialis 蝦青素與葉綠素a 含量比值的影響Fig.4 Effect of 3-hydroxybutyric acid on the content ratio of astaxanthin to chlorophyll a in H.pluvialis
試驗期間,雨生紅球藻的蝦青素與葉綠素a 含量的比值呈現波動變化,其中在第6、12、27 天出現極大值,在第9 天和第21 天出現極小值,從第27 天后又開始呈逐漸降低的趨勢。3-羥基丁酸處理對雨生紅球藻蝦青素與葉綠素a 含量比值的影響明顯高于對照組的含量比值,其中6、12、15、18、24、27、30 d 時,0.01 μg/L 組的數值顯著高于其他濃度組(P<0.05),這與Ding 等[23]使用叔丁基羥基茴香醚作為誘導物的結果相類似。試驗結果說明3-羥基丁酸影響了雨生紅球藻自身資源在光合作用與抗逆合成之間的權衡分配,而低濃度的影響作用更強。
2.2.1 3-羥基丁酸對H.pluvialis 葉綠素a 合成的影響
隨著試驗逐步進行,不同濃度的3-羥基丁酸對雨生紅球藻進行脅迫處理,雨生紅球藻細胞內的葉綠素a 含量呈波動變化趨勢如圖5 所示。

圖5 3-羥基丁酸對H.pluvialis 葉綠素a 合成的影響Fig.5 Effect of 3-hydroxybutyric acid on chlorophyll a synthesis in H.pluvialis
從圖5 中可以看出,隨著試驗時間的不斷延長,雨生紅球藻合成葉綠素a 含量變化呈不規律情況。微藻細胞內的葉綠素a 含量在試驗3、9、21 d 出現極大值,在6 d 和12 d 出現極小值,從21 d 開始又逐漸降低。試驗期間,雨生紅球藻中的葉綠素a 含量最高超過0.6 mg/L(21 d)。
2.2.2 3-羥基丁酸對H.pluvialis 蝦青素合成的影響
3-羥基丁酸對H.pluvialis 蝦青素合成的影響如圖6 所示。

圖6 3-羥基丁酸對H.pluvialis 蝦青素合成的影響Fig.6 Effect of 3-hydroxybutyric acid on astaxanthin synthesis in H.pluvialis
從圖6 中可以看出隨著試驗時間的延長,雨生紅球累積蝦青素含量逐漸緩慢呈現上升趨勢,從試驗開始至第24 天各試驗組的蝦青素水平呈緩慢上升趨勢,從第24 天開始藻細胞蝦青素含量積累出現大幅快速增長,且在第27 天蝦青素含量的積累達到了最高峰,此時0.01 μg/L 組的蝦青素含量最高達到7.57 mg/L,是對照組的1.67 倍,由此可知,3-羥基丁酸對微藻的生長雖然沒有促進作用。此后各試驗組的蝦青素水平出現迅速回落。
3 討論
3.1 3-羥基丁酸對H.pluvialis PSⅡ光化學活性的影響
葉綠素a 是微藻光合作用時需要的主要色素,是綠色葉片進行光合作用時捕獲光能的重要物質,其含量的高低在一定程度上反映了微藻光合能力的強弱[24]。當微藻受到環境脅迫時,光合作用同時受到抑制,導致光合效率降低,藻細胞吸收的部分光能通過熱量和熒光形式散發出來,因此通過測定葉綠素熒光參數可以了解微藻光合機構受脅迫而損傷的程度[25]。并且有研究證實,環境脅迫會使藻類的PSⅡ反應中心的受損,降低PSⅡ反應中心活性、電子傳遞效率和光能的轉化效率。在本研究中,通過用3-羥基丁酸處理,結果造成雨生紅球藻細胞PSⅡ光化學參數Fv/Fm、Yield 和ETR 均出現不同程度降低,說明3-羥基丁酸是雨生紅球藻生長的脅迫因子,而試驗中Yield 和ETR 之間的變化趨勢更為接近,發現它們之間具有較強相關的聯系。
3.2 3-羥基丁酸對H.pluvialis 色素合成的影響
本研究主要考察了雨生紅球藻細胞內的葉綠素a 和蝦青素的含量變化,葉綠素a 含量反映微藻的光合能力,蝦青素含量反映微藻的抗逆水平。在本試驗中雨生紅球藻細胞內的葉綠素a 含量隨時間延長呈波動變化趨勢,而3-羥基丁酸加劇了這種變化波動,這可能是微藻的光合機構在脅迫抑制和損傷修復共同作用下的結果。在試驗后期(27~30 d)雨生紅球藻的葉綠素a 含量出現極小值,說明3-羥基丁酸對微藻的脅迫超出了其修復作用的限度,這也得到了微藻3 種PSⅡ光化學參數均出現不同程度降低現象的同步印證。本研究結果表明,雖然3-羥基丁酸對雨生紅球藻的光合能力造成抑制,卻能夠提高微藻合成與積累蝦青素的能力,這與岳陳陳等[26]使用褪黑素作為誘導物的研究結果類似。此外,本試驗還發現較低濃度(0.01 μg/L) 的3-羥基丁酸有利于雨生紅球藻合成蝦青素,但當濃度較高時蝦青素的積累反而受到抑制,此現象說明0.01 μg/L 適合作為3-羥基丁酸誘導雨生紅球藻積累蝦青素的使用濃度,而高濃度3-羥基丁酸對雨生紅球藻施加的脅迫可能已經超出了其抗逆能力的限度。3-羥基丁酸對雨生紅球藻具有雙重作用,不僅具有抑制葉綠素a 合成的作用,還具有促進蝦青素積累的作用。因此,雨生紅球藻的生長和蝦青素的積累與其所處的環境脅迫有著密切關系,由于雨生紅球藻在逆境條件下積累蝦青素的自我保護機制,所以適宜脅迫條件下會增加雨生紅球藻積累蝦青素的含量。
4 結論
3-羥基丁酸對雨生紅球藻PSⅡ光反應中心造成了脅迫損傷,在脅迫條件下雨生紅球藻的光合結構和功能都受到了一定的傷害。微藻細胞PSⅡ光化學參數Fv/Fm、Yield 和ETR 均出現不同程度降低,其中0.02、0.10 μg/L 組的作用最明顯,降低的數值顯著高于其他組,并且通過蝦青素與葉綠素比值的分析同樣證明0.01 μg/L 濃度對雨生紅球藻積累蝦青素為最好的濃度。3-羥基丁酸造成雨生紅球藻葉綠素a 含量波動變化并最終下降至較低水平;3-羥基丁酸能夠促進雨生紅球藻合成并積累蝦青素,其中3-羥基丁酸的濃度為0.01 μg/L 時,雨生紅球藻積累蝦青素效果優于其他組,在到達積累峰值時最高積累量為7.57 mg/L,是對照組的1.67 倍。3-羥基丁酸會對雨生紅球藻的光合作用產生影響,同時會干涉雨生紅球藻自身資源在光合作用與抗逆合成之間的權衡分配。
