藍斑NE能神經元在束縛應激致小鼠咬肌緊張度升高中的作用
2023-11-27 12:00:52劉楊李強陳永進趙雅娟
實用口腔醫學雜志 2023年6期
劉楊 李強 陳永進 趙雅娟
口頜肌肌肉痛是顳下頜關節紊亂病(temporomandibular disorders,TMD)最常見的臨床分類,在TMD患者中的發生率約為80%,嚴重影響患者的正常生活[1-2]。據報道,存在口頜肌肌肉痛的TMD患者往往比正常人群有更高的焦慮或抑郁水平[3-4];受到慢性心理應激的人群更易出現口頜肌的緊張與疲勞,引發或加重肌肉與顳下頜關節的疼痛[5-6]。因此,應激導致的口頜肌緊張度升高與肌疲勞被認為是TMD口頜肌肌肉痛表現的重要誘發因素。
肌電活動變化是評價口頜肌功能狀態、評估TMD的發展與治療效果中重要檢測指標[7-8]。臨床研究顯示,TMD患者咬肌肌電水平明顯高于正常人群[9-10];動物實驗研究亦證實,心理應激狀態下的實驗動物會出現焦慮樣行為,并且其咬肌會出現緊張度升高以及能量代謝異常[11-12]。
位于腦干的藍斑(locus coeruleus,LC) 與機體的應激反應密切相關[13]。LC內神經元類型主要為去甲腎上腺素(noradrenaline,NE)能神經元,其合成并分泌的神經遞質NE在應激相關的行為變化、情緒、以及認知等方面發揮重要調控作用[14-15]。在解剖結構上,LC與調節口頜肌運動的重要核團三叉神經中腦核(Mesencephalic trigeminal nucleus,Vme)不僅位置相臨近,同時LC還發出神經投射至Vme,調控Vme神經元的興奮性[16]。研究證實,應激可導致Vme神經元興奮性升高,并與應激后咬肌緊張度升高密切相關[17],然而,肌緊張度升高是否是由于LC神經元被激活,進而通過神經通路投射至Vme所致,仍需進一步探究。
本研究擬采用經典的束縛應激建立動物模型,觀察小鼠應激狀態下的行為學改變與清醒狀態下咬肌肌電水平的變化,同時檢測LC神經元的激活情況,采用化學遺傳學技術特異性地抑制LC內NE能神經元的激活,探究LC在心理應激誘發口頜肌緊張度升高中的調控作用,為深入研究“應激-中樞神經-外周神經-口頜肌的全傳遞調控通路”在TMD發病中的作用和機理提供實驗及理論依據。
1 材料與方法
1.1 實驗動物
本研究共使用SPF級雄性C57BL/6小鼠40 只(6~8 周齡),由空軍軍醫大學實驗動物中心提供[許可證號:SCXK(陜)2019-001]。飼養環境溫度(25±1) ℃,濕度(50±5)%,12 h光/暗循環,隨意進食飲水。所有動物實驗經過空軍軍醫大學第三附屬醫院倫理委員會批準(批準號:K9-2022-020)。
1.2 主要實驗設備與試劑
自制束縛器;高架十字迷宮系統(RD1208-EP,上海移數公司);定制肌電采集電極(蘇州科斗腦機科技有限公司);MP46型生理記錄儀(BIOPAC,美國);立體定位注射儀(68025,深圳瑞沃德),微量進樣器(平頭1 μL,上海滬鴿);冰凍切片機(Leica CM1800,Leica,德國);FV1000激光共聚焦顯微鏡(Olympus,日本);病毒載體rAAV-TH-hM4D(Gi)-mCherry-WPRE-hGH pA(武漢樞密);疊氮平-n-氧化物(clozapine-n-oxide,CNO)(Sigma,美國);小鼠抗Fos(Abcam,英國);兔抗-TH(Millipore,美國);A594 驢抗小鼠IgG、A647 驢抗兔IgG(Thermo Fisher Scientific,美國)。
1.3 小鼠束縛應激動物模型的建立
本研究采用的束縛應激動物模型參照相關文獻[17],將應激組小鼠置于管壁帶有數個通氣孔的50 mL離心管中;每日早上8:00開始連續束縛4 h,1 次/d,共束縛14 d。在給予束縛應激的過程中,小鼠禁食禁水。對照組小鼠正常飼養。
本研究共分兩部分實驗。第一部分:將16 只小鼠隨機分為對照組與應激組(n=8),研究束縛應激是否能夠導致小鼠出現行為學改變、咬肌緊張度升高,同時觀察LC內神經元的激活情況。第二部分:將24 只小鼠隨機分為對照+溶劑組、應激+溶劑組和應激+CNO組(n=8),采用化學遺傳學技術抑制LC內NE能神經元的激活,再次觀察束縛應激后小鼠的行為學變化與咬肌的緊張度。
1.4 行為學測試
采用高架十字迷宮(elevated plus maze text,EPM)實驗檢測動物的焦慮水平[18]。EPM距離地面50 cm,由2 個相對的開放臂、2 個相對的閉合臂以及中央活動區域組成。實驗開始時,將小鼠放置在中央區域,任其自由活動5 min,通過攝像頭記錄其活動情況。通過自動分析系統,得出各組小鼠進入開臂次數百分比和開臂滯留時間百分比,作為評估焦慮水平的參數。
1.5 咬肌肌電水平檢測
采用定制電極采集小鼠咬肌肌電位(electromyography,EMG)水平。使用絕緣軟導線(長2.5 cm,直徑0.5 mm),一端焊接一根2 mm長的不銹鋼金屬微針(直徑0.15 mm),另一端焊接于2×3的圓孔排母接口,排母上焊接一根銀絲做地線。實驗開始前,參照文獻[19]將電極提前置于小鼠左側咬肌。腹腔注射戊巴比妥鈉麻醉后(35 mg/kg),在小鼠左側面頰及顱頂部備皮、消毒。切開左側下頜與顱頂皮膚,將電極微針插入左側咬肌肌腹并縫合固定,縫合面頰皮膚。將絕緣導線經耳后及顱頂皮下走行,于顱頂切口處穿出,將排母接口端用牙科自凝塑料固定于顱頂骨上,再次對術區進行消毒。
術后恢復1 周后開始進行實驗。束縛應激結束后,采集各組小鼠在清醒狀態下的咬肌肌電活動水平。采集方法:采用連接線將小鼠顱頂排母接口與MP46型2導生理記錄儀相連,置于飼養籠內適應30 min,使用AcqKnowledge軟件(Biopac公司,美國)采集小鼠清醒狀態下咬肌肌電信號(每只小鼠采集30 min),并對數據進行統計分析,得到每只小鼠咬肌的累積肌電(integral EMG,iEMG)及振幅均方根(root mean square,RMS)。
1.6 免疫組織熒光染色
采用NE能神經元的特異性標記物——酪氨酸羥化酶(tyrosine hydroxylase,TH)來標記LC神經元;采用即刻早期基因fos的表達產物——Fos蛋白標記被激活的神經元;通過免疫熒光雙重染色觀察LC內Fos陽性神經元的數量,評估應激后LC神經元激活情況。
戊巴比妥鈉腹腔注射深度麻醉后(60 mg/kg),小鼠經心臟灌流100 mL 0.01 mol/L磷酸鹽緩沖液(PBS,pH 7.4),以及200 mL 4%多聚甲醛進行灌注固定,取出腦組織進行后固定及蔗糖脫水。根據小鼠腦立體定位圖譜[20],修整腦組織塊至LC(Bregma尾側5.34~5.68 mm)部位,冰凍切片(厚30 μm),0.01 mol/L PBS漂洗,10%驢血清室溫孵育30 min,加入一抗(小鼠抗Fos,1∶500;兔抗-TH,1∶500)孵育過夜,0.01 mol/L PBS漂洗,加入二抗(A594 驢抗小鼠IgG,1∶500;A647 驢抗兔IgG,1∶500),避光孵育4 h,0.01 mol/L PBS漂洗,裱片,室溫晾干,熒光封片。激光共聚焦顯微鏡下觀察拍照,采用Image-Pro Plus 圖像分析系統計算各組LC核團內Fos陽性神經元的數量。
1.7 化學遺傳學實驗
小鼠腹腔注射戊巴比妥鈉(35 mg/kg)麻醉,頭部備皮并采用碘伏消毒后,固定于立體定位儀上,沿頭部正中矢狀位剪開皮膚1~2 cm,充分暴露顱骨。根據小鼠腦圖譜[20]定位LC(前囟尾端5.50 mm,中線外側0.90 mm,腦表面向下3.90 mm),按上述坐標在顱骨鉆孔,將攜帶有TH啟動子以及抑制性G蛋白受體基因hM4D(Gi)的腺病毒載體rAAV-TH-hM4D(Gi)-mCherry-WPRE-hGH pA(2.0E+12 vg/mL)置于微量注射器內,定位注射于LC區域(注射量0.05 μL)。術后1 周,將肌電電極埋置于左側咬肌,再恢復1 周后進行分組與實驗。待應激結束后,各組小鼠腹腔注射藥物CNO或溶劑,給藥30~40 min之后進行行為學與咬肌肌電水平的檢測。之后對小鼠進行灌注取材,切片驗證病毒轉染效果。
1.8 統計學分析
2 結 果
2.1 束縛應激可導致小鼠焦慮樣行為與咬肌肌電活動水平增強
小鼠行為學結果如圖1。應激組小鼠在高架十字迷宮的活動中,其進入開臂次數百分比、滯留開臂時間的百分比顯著低于對照組(P=0.0083,P=0.0010),提示束縛應激可導致小鼠焦慮樣行為。
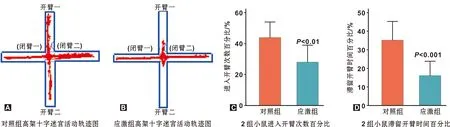
圖1 小鼠慢性束縛應激后行為學檢測結果
小鼠咬肌肌電水平結果如圖2。在受到14 d束縛應激后,應激組小鼠咬肌肌電的iEMG與RMS水平均顯著高于對照組(P=0.0002,P=0.0002),提示束縛應激可導致小鼠咬肌緊張度升高。
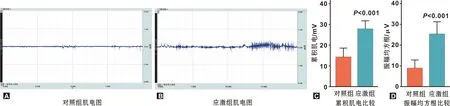
圖2 小鼠慢性束縛應激后咬肌肌電水平檢測結果
2.2 束縛應激后藍斑神經元顯著激活
兩組小鼠Fos表達結果如圖3。藍色熒光顯示為TH陽性神經元,作為LC的位置標記。紅色熒光顯示為Fos陽性神經元細胞核。在LC核團范圍內進行Fos陽性神經元計數結果顯示,應激組LC部位的Fos陽性神經元數量顯著高于對照組(P=0.0003),提示束縛應激可致LC神經元顯著激活。
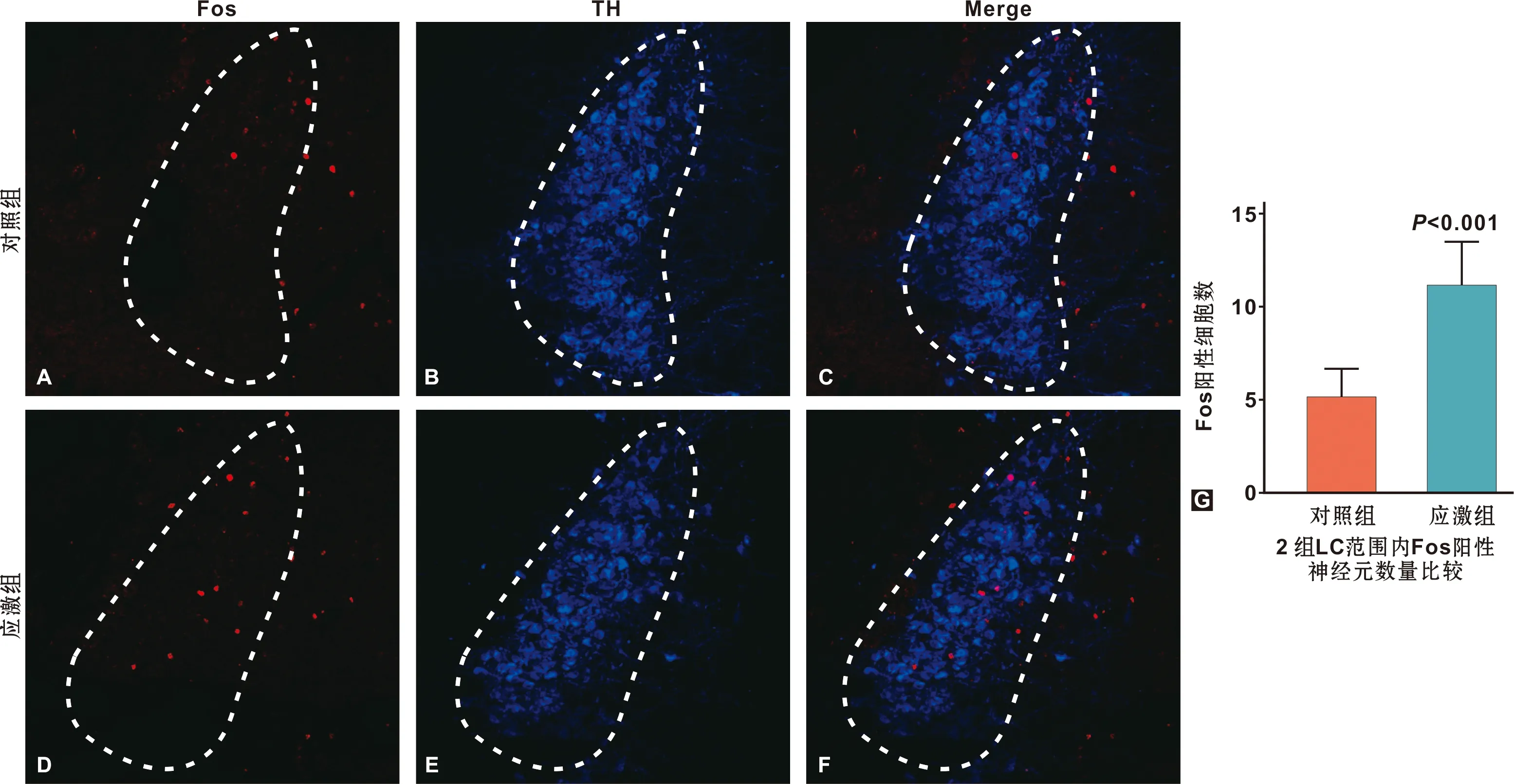
圖3 慢性束縛應激后小鼠LC內Fos染色結果
2.3 特異性抑制藍斑內NE能神經元活動可緩解束縛應激導致的焦慮行為與咬肌肌電水平
為了進一步驗證LC在束縛應激致咬肌肌電水平增高過程中的作用,采用化學遺傳學技術特異性抑制LC內NE能神經元的激活(圖4)。實驗結束后組織切片結果顯示,帶有紅色熒光的病毒載體成功轉染至LC神經元胞體內(圖4B~C),確保了給予CNO后能夠有效抑制LC內NE能神經元的激活。
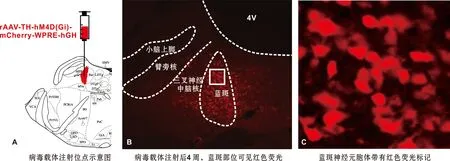
圖4 用于化學遺傳學的病毒載體轉染結果
行為學結果顯示(圖5),應激+溶劑組小鼠在高架十字迷宮中進入開臂次數百分比、滯留開臂時間的百分比低于對照+溶劑組(P=0.0007,P=0.0008),應激+CNO組小鼠高架十字迷宮中進入開臂次數百分比、滯留開臂時間的百分比高于應激+溶劑組(P=0.0218,P=0.0131)。肌電結果顯示(圖6),應激+溶劑組小鼠咬肌肌電的iEMG與RMS水平高于對照+溶劑組(P=0.0003,P=0.0002),應激+CNO組小鼠咬肌肌電的iEMG與RMS水平低于應激+溶劑組(P=0.0078,P=0.0248)。表明抑制藍斑內NE能神經元的激活可緩解束縛應激導致的焦慮行為與咬肌緊張度升高。
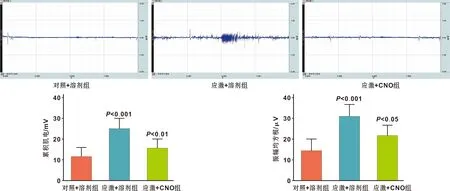
圖6 化學遺傳學給藥后各組小鼠咬肌肌電水平檢測結果
3 討 論
越來越多的研究認為,心理應激是TMD發生與發展的重要危險因素[3-6,21]。中樞神經系統是機體應激反應的重要部分,探究心理應激如何通過中樞神經通路的調控,影響TMD的發生發展,對深入理解二者密切相關的中樞層面原因、并尋求更有針對性的治療具有重要的意義。通過動物應激模型是研究心理因素與TMD的關系,探討其發生發展機制的常用方法。本研究采用小鼠建立心理應激動物模型,實驗中采用的束縛應激方式因其操作方便、重復性好等優點,在有關心理應激相關的研究中被廣泛采用[17,22]。連續14 d、4 h/d的慢性束縛應激構建心理應激動物模型,觀察到小鼠在高架十字迷宮中進入開放臂的次數與時間顯著降低,表明慢性束縛應激可成功誘導小鼠出現焦慮樣情緒,與以往的研究結果相一致[18,22]。
臨床研究表明,焦慮情緒與口頜肌的緊張度的關系可能是加重TMD癥狀的重要原因[5-6]。本研究進一步檢測了小鼠受到應激后咬肌的肌電活動水平。以往研究中,實驗動物由于難以配合,多采用麻醉狀態下記錄肌電活動,限制了實驗結果的參考價值。本課題組根據小鼠口頜部解剖特點,參照相關文獻[17],定制了植入式針式電極,將其植入小鼠的咬肌內,并在顱骨部位進行固定,可在小鼠清醒并自由活動的狀態下,進行長時間咬肌肌電的采集。結果發現,慢性束縛應激可引起咬肌肌電水平的顯著升高。咬肌是頜面部最重要的一對口頜肌,其運動由中樞內三叉神經運動核(trigeminal motor nucleus,Vmo)支配,Vme調控咬肌的本體覺傳入,并向Vmo發出谷氨酸能投射。本課題組前期的研究證實,慢性束縛應激可導致Vme神經元興奮性升高,Vme-Vmo谷氨酸能投射增強,并與應激導致的肌緊張度升高有關[17]。然而,應激后Vme神經元的興奮性增加是否是由于受到了中樞內應激相關核團的投射影響所致,仍需進一步探究。
緊鄰Vme內側的LC作為大腦的應激相關中樞之一,被認為與注意、喚醒、記憶等生理功能密切相關,并介導了壓力性應激反應[13-15]。LC中的NE能神經元是大腦內合成NE的主要部位,與應激反應密切相關,許多精神疾病諸如焦慮癥、抑郁癥都存在LC內NE能系統的失調[23-24]。本研究為證實束縛應激是否能夠引起LC神經元的直接激活,檢測了LC內即刻早期基因c-fos的表達產物Fos蛋白的表達水平[25]。TH是細胞合成NE過程中的關鍵酶,用TH來標記藍斑神經元[26],結果發現,小鼠受到14 d應激后,可觀察到應激組在TH陽性神經元的范圍內,Fos陽性細胞的數量明顯多于對照組,說明束縛應激可導致LC神經元的顯著激活。由于LC緊鄰Vme,且已有文獻證實,LC直接向Vme發出神經投射,調控Vme神經元的興奮性變化[16]。因此推測,應激后LC神經元激活,并通過向Vme的神經投射使得Vme神經元興奮性升高,從而導致咬肌的緊張度升高。為驗證以上假說,進一步采用了化學遺傳學的方法,探究應激后LC神經元的激活是否與咬肌緊張度的升高有關。
化學遺傳學的核心技術是使用可被特定藥物來激活的特定人工設計的受體(designer receptors exclusively activated by designer drugs,DREADDs),從而對神經細胞群或神經環路的活性進行特異性的控制[27]。在神經科學研究中,最常用的兩種DREADDs包括可激活神經元放電的hM3Dq和可抑制神經元放電的hM4D(Gi)。DREADDs可被生物惰性配體CNO高選擇性地激活,且對其余中樞靶點缺乏親和力[28]。以DREADDs加上CNO定向激活的化學遺傳學方法為最小侵入性、特異性調控神經元活性的研究提供了有力的技術支撐。本研究由于應激后LC神經元出現激活,故選用了抑制性DREADDs hM4D(Gi)。為了使其能夠特異性的抑制LC神經元,又添加了TH啟動子,并使用腺病毒載體輔助將構建的DREADDs定位注射入LC。在應激結束后,給予每組動物腹腔注射CNO或溶劑,結果顯示,應激+溶劑組小鼠相比對照+溶劑組出現了顯著的焦慮樣改變,而給予CNO后,其焦慮水平又顯著降低;另一方面,應激+溶劑組小鼠的咬肌肌電水平顯著高于對照+溶劑組,而給予CNO后,咬肌肌電水平又顯著下降。這進一步證明,LC內NE能神經元的激活是束縛應激導致的焦慮行為以及咬肌緊張度升高的重要原因。有文獻報道,Vme神經元細胞膜含有α2A-腎上腺素能受體,并可被來自LC的NE能神經投射所激活,繼而導致其神經元電生理特性發生改變[29]。因此,通過LC-Vme神經通路的作用,LC將神經元激活信息傳遞至Vme,繼而影響口頜肌的運動狀態,很可能是LC內NE能神經元參與調控應激導致咬肌緊張度升高的中樞機制之一。
綜上所述,慢性束縛應激能夠造成小鼠的焦慮樣情緒改變,且咬肌的肌緊張度升高;而應激后LC內NE能神經元的激活可能是應激導致焦慮樣行為與咬肌緊張度升高的中樞調控機制之一。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
人大建設(2019年12期)2019-05-21 02:55:32
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
