海金沙植物的繁殖特性及植株生長習性研究進展
2023-11-28 06:16:30童家赟黃滿霖魏汝芯詹若挺
廣州中醫藥大學學報 2023年12期
關鍵詞:研究
童家赟, 黃滿霖, 魏汝芯, 詹若挺,2
[1.廣州中醫藥大學中藥學院,廣東廣州 510006;2.嶺南中藥資源教育部重點實驗室(廣州中醫藥大學),廣東廣州 510006]
中藥海金沙來源于蕨類植物海金沙科海金沙屬草本植物[,Lygodium japonicum(Thunb.)Sw.]的干燥成熟孢子,具有清熱利濕、通淋止痛的功效[1]。現代研究表明,海金沙提取物能夠促進輸尿管的蠕動,增加輸尿管上段壓力,從而有利于結石排出;還能降低腎臟中草酸和鈣離子含量,增加排尿量,抑制草酸鈣晶體從亞穩態轉化為穩態晶體,從而降低結石形成風險[2-6]。海金沙藥材為臨床常用中成藥尿石通丸、復方石淋通片、腎石通顆粒、三金排石湯、排石顆粒等的重要原料。其中尿石通丸、復方石淋通片是治療尿路結石的常用藥,為國家二級中藥保護品種,已列入基本藥物目錄[7]。據了解,僅東莞市亞洲制藥科技有限公司對海金沙孢子粉的年需求量就超過50 噸。然而,海金沙原植物常散生于自然環境中,一直以來只能通過人工采收,藥材采收效率極低。隨著經濟的發展和人工成本的提高,藥農采收野生海金沙藥材的積極性急劇降低,供不應求導致近年來該藥材的價格較快上漲,相關中成藥的生產和可持續發展受到嚴重制約。此外,同屬植物曲軸海 金 沙(L. flexuosum)和 小 葉 海 金 沙(L.microphyllum)與海金沙形態非常相似,地理分布亦有重疊,常有誤采和混用現象[8]。因此,海金沙這種較大宗的野生來源藥材的生產,急需開展集約化和規模化種植來保證產量和臨床用藥的質量。以下對海金沙繁育和植物生長條件相關的研究概況進行綜述,以期在前人研究的基礎上優化其繁育方法和條件,為最終解決其藥材生產問題提供參考。
1 海金沙的繁殖特性及繁殖機制研究概況
1. 1 海金沙世代生活史中的組織形態的發生發育相較于種子植物從種子萌發到幼苗形成的速度,蕨類植物的幼苗形成則較緩慢和復雜。蕨類植物具有明顯的有性世代的配子體(單倍體)和無性世代的孢子體(二倍體)交替現象。早在1991年,國內學者對海金沙(Lygodium japonicum)從孢子萌發到長成幼孢子體苗的全過程進行了較為仔細的觀察,研究表明整個過程總共約需半年時間[9]。海金沙孢子經無菌播種6 d 后萌發,形成絲狀體,接著長成心形的配子體,播種后約30 ~35 d 生成精子器,接著生成頸卵器;成熟的配子體授精后約半個月長成幼孢子體,再經過4個月可以長成具3片葉的幼苗[9]。之后,日本的高橋尚子(Takahashi N)等[10]采用生物顯微鏡和金相顯微鏡觀察了海金沙孢子經萌發后形成頂端細胞分生組織(apical-cellbased meristem)和多細胞分生組織(multicellular meristem)兩種分生組織,以及細胞經過一系列平周和垂周分裂后而形成心形配子體的全過程。劉朝輝等[11]采用透射電鏡觀察了海金沙的精子細胞發育過程,發現其結構發育與薄囊蕨亞綱的其他植物相似。郭嚴冬、Cao J G 等[12-13]采用樹脂包埋切片、透射電鏡和細胞化學技術觀察了海金沙配子體發育生成頸卵器和卵細胞的過程。邵文等[14]采用石蠟切片觀察了海金沙頸卵器中卵子受精后產生合子、合子進行細胞分裂形成四分體、最后形成基足細胞和營養器官(第一和第二葉)的整個過程。由以上研究可知,目前海金沙從孢子萌發到長成幼孢子體的組織形態發生全過程已經比較清楚,其示意圖見圖1。
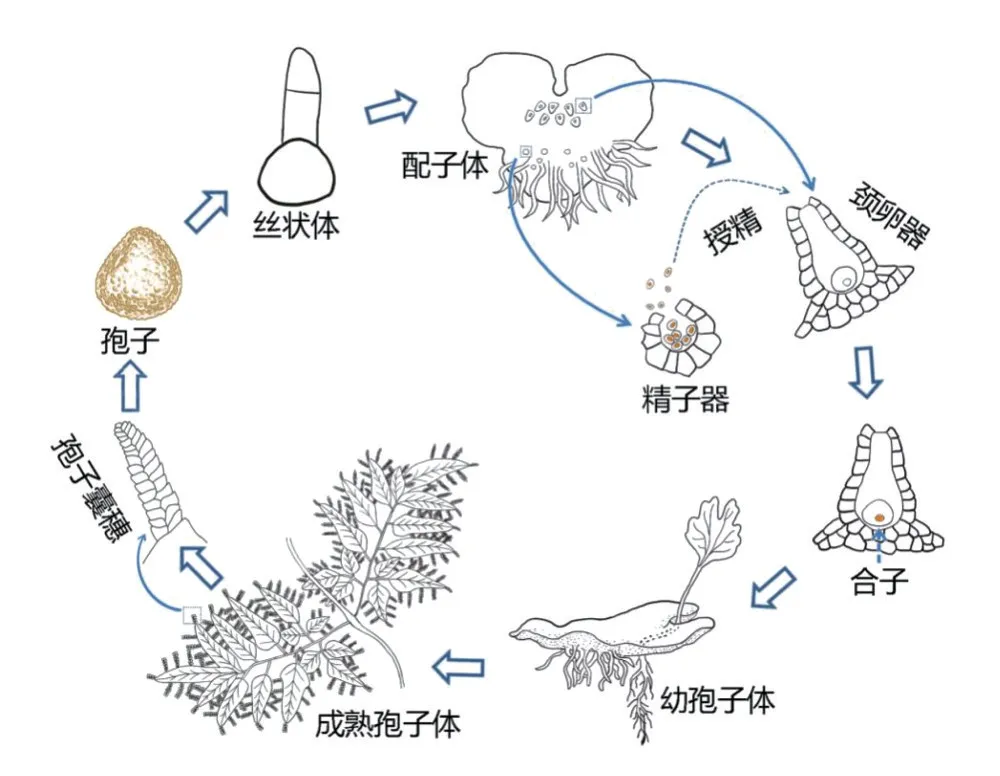
圖1 海金沙生活史示意圖Figure 1 Schematic diagram of the life history of, Lygodium japonicum
1.2 海金沙配子體性別決定機制
1. 2. 1 遺傳因素和內在激素調控雌雄配子體的生成 早在1960 年,Ulrich N[15]就證實了在無菌培養過程中,海金沙的成熟配子體提取物添加入培養基后能誘導其他年幼配子體提前產生精子器。1975 年,竹野清俊(Takeno K)等[16-17]發現外源的赤霉素類(gibberellins,GAs)可誘導海金沙配子體的精子器形成;赤霉素類的活性強度由大到小排列為GA7>GA4>GA9>GA3>GA5>GA1>GA8;隨 后發現外源的赤霉素類會抑制頸卵器的生成,活性強 度 由 大 到 小 排 列 為GA4= GA9>GA7>GA3>GA1=GA5=GA8。1979 年,竹野清俊等[18-19]驗證了海金沙配子體提取物也會抑制頸卵器的分化;隨后在原葉體培養基中加入外源的成精子囊素(antheridiogen)如GA9等,發現GA9的抑制效應與配子體提取物相當,認為其機制可能與由GA9轉化生成的GA9-甲酯發揮的效應有關;但是其控制配子體在不同時間點產生性別分化的機制尚不明確。
竹野清俊等[20]基于以上研究之后,通過實驗觀察提出了海金沙配子體性別分化時間序列模型:當居群內的原葉體(配子體)處于不同的發育階段時,最先達到1.5 mm 寬的原葉體形成頸卵器,這時只發育形成單性的雌性配子體。然后,原葉體開始生成控制生殖器官形成的天然物質;但這些物質并不影響那些較大的正在分化頸卵器的單性的雌性原葉體。而居群內發育遲緩的原葉體對這些大的原葉體分泌的物質非常敏感,使得頸卵器的形成受到抑制,從而產生精子器,也就是使這些更小的原葉體發育形成單性的雄性配子體。最終,雌性原葉體可以在不受外源成精子囊素的影響下產生精子器,從而產生雙性配子體。
2014 年,日本的田中君木(Tanaka J)等[21]克隆了海金沙赤霉素合成途徑中相關基因并作表達模式分析,并通過蛋白雙雜交和免疫印跡等一系列試驗,闡明了蕨類植物赤霉素合成途徑及其信號傳導的時空分離是決定配子體性別的重要分子機制。田中君木等認為,首先,早熟的配子體生成GA9,然后被甲酯化成GA9-甲酯, GA9-甲酯被分泌到環境中,從而被鄰近的晚熟配子體(絲狀體階段)攝取;GA9-甲酯被攝取之后,在晚熟的配子體細胞內被脫甲酯重新生成GA9,之后GA9被GA3氧化酶(GA3ox)催化生成GA4,接著具有生物活性的GA4通過GA4→GID→DELLA 通路觸發了配子體上精子器的生成,但抑制了頸卵器的發生。
1.2.2 環境因素影響雌雄配子體的生成 蕨類植物的性別分化除了受到自身遺傳因素和內在激素調控外,也受到培養密度、營養條件、光照等環境因素的影響[22]。高曉雯等[23]的研究表明,水分對配子體形態發育的影響也較大。隨著水分的增加,海金沙孢子的萌發速率加快,絲狀體的長度增加。在水分不足條件下,配子體易發育成雄性配子體,而在水分充足條件下,配子體易發育成兩性配子體或雌性配子體。次生假根的長度也可隨著水分的增加而增加。但是該研究未對培養過程的基質加水量進行準確計量。后續研究需要參照該報道的方法進一步確定有利于分化生成兩性配子體或適當比例的單性配子體及有利于授精產生合子的最適合水分量。
2 海金沙的育苗影響因素研究概況
2.1 影響孢子萌發因素的研究蕨類植物孢子萌發除了受到自身遺傳因素和內在激素調控外,也受到培養密度、營養條件、溫度、光照等環境因素的影響[24-25]。如紫萁(Osmunda japonica)孢子播種密度在100 ~3 000 個/cm2的區間時,隨著密度的升高,孢子的萌發率越高,密度為3 000 個/cm2時的萌發率可達95.4%;而當密度達到5 000 個/cm2時,其萌發率反而降低[26]。目前,海金沙播種密度試驗未見相關報道。無機鹽方面,郭治友等[27]的研究認為,低鹽的Knop’s培養基和全量(高鹽)的MS培養基對海金沙孢子萌發影響不顯著。但也有研究顯示,Knop’s 培養基、1/8 的MS 培養基和1/4 的MS培養基上的孢子萌發率較MS培養基更高[28]。兩篇報道互相矛盾,有待進一步驗證。
有研究認為,冷凍保存可顯著降低小葉海金沙(L.microphyllum)孢子萌發率,但是對海金沙孢子萌發無顯著影響[29]。海金沙孢子在低溫條件下保存多長時間后而不會顯著降低萌發率,這也是值得探討的問題。
2.2 雌雄配子體交配和幼孢子體生成的研究竹野清俊(Takeno K)等[30]開展的分離雙性配子體并與單性配子體進行交配的試驗顯示,只有約30%的雌性配子體生成幼孢子體,而單個的雙性配子體未能形成幼孢子體;并認為該結果可能是由配子體內自交產生的純合子不能發育導致的。但是,Lott M S 等[31]通過研究認為,海金沙孢子萌發后,成熟配子體主要是雌雄同體型,同一配子體內自交高于90%。以上二者的研究結論相去甚遠,需要進一步證實。
Sakaushi S 等[32]在高速視頻顯微鏡、透射電子顯微鏡(TEM)和掃描電子顯微鏡(SEM)下觀察發現,海金沙精子的鞭毛朝向同一方向,并且只能朝前方游動,即使遇到障礙物也不會向后游動;并認為這個特點可能會影響雌雄配子體的授精。到底有多少個孢子形成配子體后能成功授精成為合子并生成幼孢子體,其最適合的環境因素有哪些,目前未見相關的報道。
2.3 幼苗繁育(植株再生)研究海金沙可采用分株繁殖的方式,操作上比較簡單[33],但分株繁殖方式的繁殖系數較低,滿足不了規模化種植對大量種苗的需求,而通過孢子萌發有可能實現種苗的大規模生產。由于蕨類植物生活史(世代交替史)的復雜性,要通過孢子萌發途徑進行海金沙的繁殖和植株再生,相較一般的種子植物步驟更繁瑣,要解決的問題也更多。目前僅見Maeda M等[34-35]對海金沙幼孢子體葉片提取的原生質體進行了植株再生的初步研究;郭治友等[27]采用Knop’s 培養基、MS 培養基等進行了孢子無菌萌發和植株再生的初步研究;另有將海金沙孢子播于泥炭土上進行萌發和育苗的專利報道[36]。有關海金沙孢子萌發育苗方面的研究報道非常少,迄今也未見采用海金沙幼芽或嫩葉進行愈傷組織培養擴繁的研究。
3 海金沙植株生長習性研究現狀
目前,對海金沙進行規模化和集約化種植的研究尚未見報道,以下只是針對野生或少量盆栽植株生長狀態的觀察。
3. 1 光照對海金沙植株生長的影響一般認為,蕨類植物大多是陰生植物,海金沙野生植株也常生長在樹林下有遮蔭的環境中。張澤宏等[37]觀察了海金沙等5 種蕨類植物葉片解剖結構,根據葉片薄、沒有海綿組織和柵欄組織的分化、氣室大等特征,認為這些是蕨類適應陰生環境的特征。但目前海金沙植株生長最適合的光照強度或遮蔽度并未見報道。
3.2 溫度與濕度對海金沙生長發育的影響溫度與濕度對海金沙植株的生長發育會產生一定的影響,冬季低溫和干旱條件下可出現植株枯萎或枯葉[38];對普通溫室內培養的11 種蕨類植物生長發育進行為期一年半的觀察,只有海金沙和鹿角蕨(Platycerium wallichii)兩種植物未生成孢子,推測可能是由于溫度與濕度不適合所致[38]。另有學者觀察發現,低海拔氣候溫暖地區的海金沙植株能正常產生孢子,而在高海拔冷涼地區則不能[39]。有研究表明,近緣物種小葉海金沙(L.microphyllum)在較高溫度環境中的群落長勢明顯較好;室內模擬環境下溫度與其光合速率呈顯著正相關[40]。但適合海金沙植株生長的溫度和水分條件范圍至今尚不清楚。
3.3 海金沙的病蟲害值得注意的是,海金沙和小葉海金沙在美國東南部的佛羅里達州和澳大利亞等地被認為是嚴重影響生態的入侵植物,多位學者從生物防治的角度研究了真菌感染和飛蛾對海金沙生長的影響。如Rayachhetry M B 等[41-43]學者從染病植株上分離病菌后重新感染植株,發現假柄銹菌(Puccinia lygodii)或者二級霉(Bipolaris sacchari)感染可致海金沙和小葉海金沙葉片壞死。有學者通過觀察發現,炭疽病菌(Glomerella cingulata)感染可使海金沙和小葉海金沙頂梢枯死,小羽葉邊緣褐變和萎蔫[44-45]。此外,還發現鱗翅目(Lepidoptera)草螟科(Crambidae)的兩種飛蛾,Neomusotima conspurcatalis和,Austromusotima camptozonale特 異 性地產卵在海金沙和小葉海金沙的植株上,且其幼蟲啃食能有效控制宿主植物的生長[46-47]。在未來規模化種植生產過程中,以上真菌和飛蛾的防治應當引起重視。
4 小結和展望
目前,國內外對于蕨類植物的繁殖特性及繁殖技術研究主要集中于巢蕨等觀賞或瀕危蕨類植物[48-50],藥用蕨類植物相關研究則很少。海金沙從孢子萌發到生成配子體,再長成幼孢子體的全過程,其組織形態發生業已清楚;配子體性別決定機制的遺傳因素和內在激素調控也得到了闡明。但是,適合孢子萌發、配子體授精等繁殖過程中的環境因素有待進一步優化;迄今未見規模化的育苗和規范化種植技術研究。
目前,至少還有以下問題需要進一步探討:①篩選最適合海金沙孢子萌發的條件,包括培養密度、營養量(包括無機鹽、蔗糖等)、溫度及光照等。②篩選適合孢子貯藏且有利于其在一段時間內保持活力的溫度與濕度條件。③篩選孢子形成配子體及授精成為合子,并生成幼孢子的最適合環境條件,如影響配子體授精的溫度、水分、培養密度、光照等。④研究建立外殖體(幼芽或嫩葉)進行無性繁殖的離體快繁體系。⑤進行栽培技術研究:如探究適合植株生長和孢子囊生成的光照、水肥、溫度等條件;研究田間管理方式,如搭設支架等對植株生長和孢子囊生成的影響;探究適宜的采收期。建議今后應通過對上述影響海金沙育苗和栽培的關鍵技術條件進行系統的研究、探索和優化,為海金沙藥材生產提供技術支撐,以維持其藥材的可持續發展及保障有關中成藥原料的供應。
(我校中藥學院藥用植物教研室教師彭光天博士和童毅博士對本文進行了審閱并提出意見和建議,特此致謝!)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
