離水停食脅迫對中華鱉(Trionyx sinensis)翻身能力、血清生化、裙邊質(zhì)構(gòu)及臟器相關(guān)功能酶活力的影響*
2023-12-05 12:26:52鄭嘉豪申屠琰徐英杰李有志朱衛(wèi)東王志錚
海洋與湖沼 2023年6期
鄭嘉豪 申屠琰, 徐英杰 李有志 朱衛(wèi)東 王志錚
離水停食脅迫對中華鱉()翻身能力、血清生化、裙邊質(zhì)構(gòu)及臟器相關(guān)功能酶活力的影響*
鄭嘉豪1申屠琰1, 2徐英杰1李有志1朱衛(wèi)東2王志錚1①
(1. 浙江海洋大學(xué)水產(chǎn)學(xué)院 浙江舟山 316022; 2. 余姚市水產(chǎn)技術(shù)推廣中心 浙江余姚 315400)
中華鱉()歷來有活品消費的習(xí)慣, 離水停食是其批量銷售環(huán)節(jié)必須承受的脅迫。因此, 探究離水停食對其商品價值的影響過程與機制, 對于明確其活體銷售貨架期具有重要指導(dǎo)價值和現(xiàn)實意義。以2020年5月20日同池起捕后離水停食處理0 d (S0實驗組)、4 d (S4實驗組)、8 d (S8實驗組)、12 d (S12實驗組)、16 d (S16實驗組)的雄性商品鱉[平均體質(zhì)量(546.86±94.70) g]為研究對象, 在常溫條件下較系統(tǒng)開展了離水停食對其翻身能力、血清生化、裙邊質(zhì)構(gòu)及臟器酶活力的影響研究。結(jié)果表明: (1) 實驗鱉連續(xù)翻身次數(shù)呈S4≈S8>S0≈S12≈S16, 且均主要集中于1 min內(nèi); (2) 血清TP、ALB、TC、LDL-C均無組間差異(>0.05), GLU-O呈S0≈S4≈S8≈S12>S16, TG和HDL-C除S4、S8的TG含量小于S0(<0.05), S12的HDL-C含量大于S0(<0.05)外, 其余實驗組間均無差異(>0.05); (3) 所測7項裙邊質(zhì)構(gòu)指標(biāo)中, 僅硬度與連續(xù)翻身次數(shù)的排序完全重合, 均呈S4≈S8>S12≈S16; (4) 肝脂肪酶和胃蛋白酶活力分別呈S4>S0≈S8≈S12≈S16和S0≈S16>S4≈S8≈S12, 腸淀粉酶活力除S16實驗組顯著大于S0、S4(<0.05)外, 其余均無組間差異(>0.05); (5) 肝臟ATP酶和抗氧化酶活力隨離水停食時間的延長總體上均呈階梯式上升趨勢, 心臟抗氧化酶活力總體上在離水停食0~4 d、4~12 d、12~16 d分別呈下降、穩(wěn)定和上升趨勢, 肺抗氧化酶活力總體上在0~4 d、4~8 d、8~12 d和12~16 d依次呈上升、下降、穩(wěn)定和再次上升的趨勢, S4、S8和S12間在各臟器抗氧化酶活力值排序上明顯表露錯位的僅為脾臟GSH-PX, 呈S4 中華鱉; 離水停食; 翻身; 裙邊質(zhì)構(gòu); 血清生化; 消化酶; ATP酶; 抗氧化酶 中華鱉()隸屬于爬行綱(Reptilia), 龜鱉目(Chelonia), 鱉科(Trionychidae), 鱉屬(), 作為我國居民喜食的傳統(tǒng)名貴食材和高檔滋補品, 歷來有活品消費的習(xí)慣。浙江省是我國中華鱉的養(yǎng)殖主產(chǎn)區(qū), 最近20余年間產(chǎn)量和產(chǎn)值均一直居全國首位。據(jù)《2022中國漁業(yè)統(tǒng)計年鑒》報道, 浙江省2021年中華鱉養(yǎng)殖產(chǎn)量為99 764 t, 占全國鱉養(yǎng)殖總產(chǎn)量的27.3% (農(nóng)業(yè)農(nóng)村部漁業(yè)漁政管理局等, 2022), 中華鱉養(yǎng)殖產(chǎn)業(yè)現(xiàn)已是助力浙江沿江沿湖地區(qū)鄉(xiāng)村振興和共同富裕的重要抓手之一。 在水產(chǎn)品供應(yīng)鏈環(huán)節(jié), 活品的市場迎合度與銷售價格較鮮品和凍品均具明顯優(yōu)勢。最近10年間, 隨著“農(nóng)超對接”和“互聯(lián)網(wǎng)直銷”等新型商業(yè)銷售模式的興起, 以及水產(chǎn)品保活技術(shù)的日臻成熟, 我國水產(chǎn)品活銷產(chǎn)業(yè)獲得了井噴式發(fā)展。貨架期是指符合標(biāo)簽規(guī)定條件下保持食品質(zhì)量(品質(zhì))的期限(國家技術(shù)監(jiān)督局, 1994), 其時長主要取決于配方、加工工藝、包裝和貯藏條件。水產(chǎn)動物的活力與其形態(tài)、生理健康度和食用口感均密切相關(guān)。作為初級水產(chǎn)品的重要組成部分, 活力既是活品有別于鮮品和凍品貨架期研究的主要內(nèi)容, 也是衡量活品品質(zhì)優(yōu)劣程度和判定打折或下架與否的關(guān)鍵因素, 但迄今國內(nèi)外尚未見有關(guān)水產(chǎn)活品活力與貨架期間相互關(guān)系的系統(tǒng)報道, 關(guān)于中華鱉的貨架期研究也僅見其加工產(chǎn)品(張師師, 2006)。中華鱉以肺呼吸為主, 離水停食是其活體批量運輸和銷售環(huán)節(jié)中必須承受的脅迫。研究表明, 饑餓脅迫不僅會引起水生動物形體特征、消化生理、抗氧化生理、血清生化和組織質(zhì)構(gòu)等的一系列變化(謝小軍等, 1998; 王志錚等, 2006; 李有志等, 2022), 也勢必會導(dǎo)致水生動物機體抗逆性能、活力和健康度的顯著下降。因此, 探究進而揭示離水停食脅迫對水產(chǎn)養(yǎng)殖動物的活力、食用口感和機體相關(guān)生理的影響過程與機制, 對于指導(dǎo)其活體銷售具有重要指導(dǎo)價值和現(xiàn)實意義。農(nóng)超活銷實踐也發(fā)現(xiàn), 中華鱉的食用口感與其離水停食時長有著極為密切的關(guān)系。因此, 確定中華鱉合理的貨架時長, 對于切實保障消費者權(quán)益和維護企業(yè)商譽具重要現(xiàn)實意義。鑒于此, 本研究團隊于2020年5~6月間在常溫條件下較系統(tǒng)開展了離水停食脅迫對中華鱉翻身能力、血清生化、裙邊質(zhì)構(gòu)及臟器相關(guān)功能酶活力的影響研究, 旨為合理規(guī)范中華鱉的貨架期提供科學(xué)依據(jù), 并為中華鱉的生理生態(tài)研究提供基礎(chǔ)資料。 實驗用中華鱉()購自余姚市冷江鱉業(yè)有限公司, 為2020年5月20日同塘起捕養(yǎng)殖4年的商品鱉。起捕后, 隨機選取其中肢體完整、反應(yīng)敏捷、無病無傷、規(guī)格相近的雄性個體作為研究對象[共計165 ind., 平均體質(zhì)量(546.86±94.70) g], 并即刻移入該公司加工車間內(nèi)在常溫條件下進行離水停食處理, 并于每日上午8:00用自來水清洗鱉體1次。 于2020年5月20日(離水停食處理0 d, 記為S0)、5月24日(離水停食處理4 d, 記為S4)、5月28日(離水停食處理8 d, 記為S8)、6月1日(離水停食處理12 d, 記為S12)和6月5日(離水停食處理16 d, 記為S16)分別任取35只供試鱉作為實驗用鱉。其中, 30只用于翻身能力測試, 剩余的5只用于血清生化、裙邊物性及臟器酶活力的測定。 1.2.1 翻身能力的測試 (1) 連續(xù)翻身次數(shù)的測定: 任取其中15只實驗鱉, 借助人工將鱉體處于背部朝下、腹部朝上狀態(tài)后讓其自主翻身復(fù)原, 當(dāng)實驗鱉翻身復(fù)原后, 借助人工迅速再次將鱉體恢復(fù)至背部朝下、腹部朝上的狀態(tài)讓其自主翻身復(fù)原, 如此循環(huán)往復(fù), 當(dāng)實驗鱉在30 s內(nèi)不能完成自主翻身復(fù)原狀態(tài)時, 立即終止測試并記錄其連續(xù)翻身總次數(shù)。 (2) 1 min內(nèi)翻身次數(shù)的測定: 取剩余的15只實驗鱉, 以1 min為測試總時長測定并記錄其翻身總次數(shù)。 1.2.2 血清生化指標(biāo)的測定 75%酒精消毒實驗鱉頸部后, 用刀片割斷其頸動脈并立即采血制成抗凝血樣。血樣于3 000 r/min離心10 min后, 將分離獲得的血清即刻置于4 °C冰箱中保存?zhèn)錅y。借助日立7020型全自動生化分析儀測定血清中的總蛋白(TP)、白蛋白(ALB)、高密度脂蛋白膽固醇(HDL-C)、低密度脂蛋白膽固醇(LDL-C)、總膽固醇(TC)、甘油三脂(TG)和血糖(GLU-O), 所用測定試劑均購自寧波美康生物工程有限公司。 1.2.3 臟器酶活力的測定 于冰盤上逐只解剖采血后的實驗鱉, 分別摘取心、肝、脾、肺、胃、腸, 并去除其內(nèi)容物和附于臟器表面的脂肪與結(jié)締組織。已摘取的所有臟器經(jīng)4 °C雙蒸水沖凈、濾紙吸干表面水分并借助分析天平(Sartorius BP211D型, 精度0.1 mg) 逐一稱重后, 立即分別放入已做好標(biāo)記的樣品袋內(nèi)并置于–80 °C冰箱中保存?zhèn)錅y。測定儀器為島津UV-1240型紫外可見分光光度計, 各類消化酶和抗氧化酶的酶活力測試盒均購自南京建成生物工程研究所, 測試步驟及計算方法均按所附說明書。 1.2.4 裙邊質(zhì)構(gòu)的測定 取臟器后, 按李有志等(2022)的方法逐只取實驗鱉尾根部上側(cè)的裙邊組織, 制成10 mm×7 mm×4 mm規(guī)格的鮮樣后, 立即采用TA.XTplus型質(zhì)構(gòu)分析儀逐只測定硬度、黏性、彈性、內(nèi)聚性、膠黏性、耐咀性和回復(fù)性。質(zhì)構(gòu)儀采用TPA測量模式, 測量探頭為P/36R, 測前、測中和測后速度分別為1、0.5和1 mm/s, 測距2 mm, 測試時間間隔5 s。 整理所得各項實驗數(shù)據(jù), 借助SPSS17.0軟件分別統(tǒng)計各實驗組的翻身次數(shù)、血清生化指標(biāo)、裙邊質(zhì)構(gòu)指標(biāo)、臟器酶活力的均值與標(biāo)準(zhǔn)差, 并分別采用檢驗和LSD多重比較法檢驗組內(nèi)、組間差異顯著性(視<0.05為顯著水平)。 觀察發(fā)現(xiàn), 實驗鱉具有通過伸展頭頸部并用頭頂住身體接觸面實現(xiàn)快速翻身復(fù)原的行為學(xué)特性。由圖1可見, 離水停食對實驗鱉翻身能力的影響主要表現(xiàn)為: (1) 連續(xù)翻身次數(shù)和1 min內(nèi)翻身次數(shù)均呈以S4、S8實驗組為最多(<0.05), 其余實驗組間均無差異(>0.05)的先升后降趨勢; (2) 連續(xù)翻身次數(shù)主要集中于1 min內(nèi), 其與1 min內(nèi)翻身總次數(shù)存在差異(<0.05)的實驗組僅為S4和S8。綜上表明, 實驗鱉具很強的翻身欲望, 過長的離水停食時間將顯著降低其連續(xù)翻身能力和1 min內(nèi)的翻身總次數(shù)。 由表1可見, 離水停食對血清生化指標(biāo)的影響主要表露為: (1) 從蛋白質(zhì)組成看, TP、ALB含量均無組間差異(>0.05); (2) 從血脂組成看, TC、LDL-C含量均無組間差異(>0.05), 除S4、S8的TG含量小于S0(<0.05), S12的HDL-C含量大于S0(<0.05)外, 其余實驗組在TG和HDL-C含量上均無組間差異(>0.05); (3) 從血糖水平看, GLU-O含量以S16實驗組為最低(<0.05), 呈S0≈S4≈S8≈S12>S16。 圖1 離水停食時長對實驗鱉翻身次數(shù)的影響 注: S0、S4、S8、S12、S16分別為離水停食處理0 d、4 d、8 d、12 d、16 d實驗組; 圖內(nèi)標(biāo)不同字母表示組間存在顯著差異(<0.05)。下同 由表2可見, 離水停食對裙邊質(zhì)構(gòu)指標(biāo)的影響主要表露為: (1) 硬度、彈性和膠黏性分別呈S0≈S4≈S8> S12≈S16、S0≈S4≈S8≈S12≈S16(>0.05)和S4≈S8≈S16>S0≈S12; (2) 黏性和內(nèi)聚性, 除S4的黏性顯著小于S8(<0.05), S4和S8的內(nèi)聚性顯著大于S0(<0.05)外, 其余均無組間差異(>0.05); (3) 耐咀性和回復(fù)性, 除S4的耐咀性和S12、S16的回復(fù)性與其他實驗組均無差異(>0.05)外, 其余則分別呈S8≈S16>S0≈S12和S4≈S8>S0。由此可知, 上述裙邊質(zhì)構(gòu)指標(biāo)中僅硬度與其連續(xù)翻身次數(shù)(圖1)的組間排序完全重合, 均呈S4≈S8>S12≈S16。 表1 離水停食對實驗鱉血清生化指標(biāo)的影響 Tab.1 Effects of air exposure and starvation on serum biochemical indicators of experimental turtles 注: TP: 總蛋白; ALB: 白蛋白; HDL-C: 高密度脂蛋白膽固醇; LDL-C: 低密度脂蛋白膽固醇; TC: 總膽固醇; TG: 甘油三脂; GLU-O: 血糖 表2 離水停食對實驗鱉裙邊質(zhì)構(gòu)的影響(=5) Tab.2 Effects of air exposure and starvation on the soft tissue skirt quality of experimental turtles (n=5) 由圖2可見, 肝脂肪酶和胃蛋白酶活力分別呈S4>S0≈S8≈S12≈S16和S0≈S16>S4≈S8≈S12, 腸淀粉酶活力除S16實驗組大于S0、S4(<0.05)外, 其余均無組間差異(>0.05)。從離水停食時序上看, S4實驗組肝脂肪酶和S16實驗組胃蛋白酶活力的顯著高企意味著兩者均為消化生理發(fā)生顯著改變的關(guān)鍵時間節(jié)點。 圖2 離水停食時長對實驗鱉消化酶活力的影響 由圖3可見, 離水停食對肝臟ATP酶和抗氧化酶活力的影響主要表露為: (1) Na+/K+-ATPase、Ca2+/ Mg2+-ATPase酶活力分別呈S0 由圖4可見, 離水停食對心臟抗氧化酶活力的影響主要表露為: (1) SOD、POD酶活力分別呈S0≈S4≈ S8≈S12≈S16(>0.05)和S0≈S16>S4≈S8≈S12; (2) CAT和GSH-PX酶活力, 除S8的CAT和S4的GSH-PX酶活力均小于S0(<0.05)外, 其余均無組間差異(>0.05)。無疑, 心臟POD酶活力的顯著高企是導(dǎo)致S0實驗組個體連續(xù)翻身能力偏弱的重要內(nèi)因。 由圖5可見, 離水停食對脾臟抗氧化酶活力的影響主要表現(xiàn)為: (1) SOD酶活力除S8、S12與其他實驗組均無差異(>0.05)外, 其余呈S0≈S4 由圖6可見, 離水停食對肺抗氧化酶活力的影響主要表露為: (1) SOD和CAT酶活力分別呈S4>S0≈ S8≈S12≈S16和S16>S0≈S4≈S8≈S12; (2) POD酶活力除S0低于S4和S16(<0.05)外, 其余均無組間差異(>0.05); (3) GSH-PX酶活力除S12與其他實驗組均無差異(>0.05)外, 其余呈S4≈S16>S0≈S8。綜上, 從離水停食時序上看, 離水停食8~12 d為SOD酶活力顯著回落后, 以及CAT酶活力顯著上升前的穩(wěn)定期。 Arnold(1983)指出, 動物的運動能力和行為表現(xiàn)與其適合度密切相關(guān)。鑒于龜鱉類動物在自然生境中因翻身遲緩更易受干旱失水和被捕食等的威脅, 故常將翻身能力視為與其生存直接相關(guān)的特性之一(Finkler, 1999; Delmas, 2007)。無疑, 本研究中實驗鱉連續(xù)翻身次數(shù)均主要集中于1 min內(nèi)的結(jié)果(圖1), 既印證了翻身作為其生存直接相關(guān)特性所表露出的強敏感性, 也映射出其為完成連續(xù)翻身所需償付的巨大能耗。徐本力(1984)指出, 運動員在承受最適負荷后往往會表露出運動積極性下降和休息時間延長的嚴重疲勞狀態(tài)。這一現(xiàn)象在動物界中也普遍存在, 按此類比, 本研究將30 s內(nèi)不能完成自主翻身復(fù)原作為終止實驗鱉連續(xù)翻身的實驗設(shè)計, 符合最適負荷下的體能測試原則。綜上, 可在生產(chǎn)和銷售實踐中將連續(xù)翻身能力作為評價中華鱉活力或體質(zhì)強健程度的重要依據(jù)。 圖3 離水停食時長對實驗鱉肝臟ATP酶和抗氧化酶活力的影響 運動負荷等級可粗分為大、中、小三級, 一般將竭盡全力只能完成1~7次、8~18次和19次以上分別評為大、中、小(徐本力, 1984) 。本研究亦屬竭力負荷測試, 按此類比, S0、S12、S16實驗組與S4、S8實驗組的連續(xù)翻身運動分別屬大負荷和中負荷運動(圖1)。與此同時, S0、S12、S16實驗組的連續(xù)翻身次數(shù)均與其1 min內(nèi)的總翻身次數(shù)無差異(>0.05), 而S4、S8實驗組的連續(xù)翻身次數(shù)則均與其1 min內(nèi)的總翻身次數(shù)存在顯著差異(<0.05)的結(jié)果, 也印證了圖1中S0、S12、S16實驗組與S4、S8實驗組間在翻身運動負荷上的差異。據(jù)報道, 余姚地區(qū)池養(yǎng)中華鱉的捕食活躍期、越冬準(zhǔn)備期、越冬休眠期和活動復(fù)蘇期依次為5月下旬至9月下旬、10月上旬至11月下旬、12月上旬至次年3月下旬、4月上旬至5月下旬(李有志等, 2022)。也就是說, 除S0實驗組個體尚處于活動復(fù)蘇期末期外, 其余實驗組個體均已進入捕食活躍期。故, 圖1所示結(jié)果無疑表明實驗鱉的連續(xù)翻身能力不僅與所處的活動階段有關(guān), 也與因離水停食時間過長導(dǎo)致的體能顯著下降有關(guān)。 抗氧化酶系統(tǒng)在保持動物體內(nèi)自由基代謝平衡和維系細胞正常代謝中具重要作用(王志錚等, 2013a)。心、肺、脾、肝的健康狀況與動物機體的免疫力、抗疲勞能力和持續(xù)運動能力具強相關(guān)性。因此, 上述這些臟器的抗氧化酶活水平對于指示受檢動物在最適運動負荷下的耐受能力具重要參考價值。以S0實驗組為參照, 本研究中肝臟抗氧化壓力以S16為最高, S0為最低(圖3), 心臟抗氧化壓力以S0、S16為最高, S4、S8為最低(圖4), 脾臟抗氧化壓力以S16為最高, S0、S4為最低(圖5), 以及肺抗氧化壓力顯著高于S0的僅為S4和S16(圖6)的情形, 并結(jié)合連續(xù)翻身次數(shù)呈S4≈S8>S0≈S12≈S16(圖1)的結(jié)果, 既反映了較高的心臟抗氧化壓力是致處于活動復(fù)蘇期末期的S0實驗組個體翻身能力偏弱的客觀事實, 也揭示了過長的離水停食時間是引發(fā)S16實驗組個體心、肝、脾、肺抗氧化壓力全面顯著高企并致其負荷耐受能力顯著下降的主因。一般而言, 生物有機體在正常生理狀況下的抗氧化酶活力往往與其抗氧化壓力呈正相關(guān), 反之亦然(李有志等, 2022)。無疑, S4、S8、S12實驗組在心、肝、脾、肺各抗氧化酶酶活力值排序上, 僅脾臟GSH-PX呈S8>S12>S4(<0.05) (圖3, 圖4, 圖5, 圖6), 表露嚴重錯位的結(jié)果, 表明S12組實驗個體連續(xù)翻身能力的顯著下降應(yīng)與其脾臟抗氧化生理已出現(xiàn)一定程度的紊亂有關(guān)。綜上可知, 由離水停食處理時間過長導(dǎo)致的免疫力和抗疲勞能力下降是引發(fā)S12和S16兩實驗組個體持續(xù)翻身能力顯著弱于S4和S8的主因。 圖4 離水停食時長對實驗鱉心臟抗氧化酶活力的影響 圖5 離水停食時長對實驗鱉脾臟抗氧化酶活力的影響 圖6 離水停食時長對實驗鱉肺抗氧化酶活力的影響 據(jù)報道, 饑餓狀態(tài)下水生動物對蛋白質(zhì)的利用往往是在脂肪被大量消耗之后(Stirling, 1976; Kutty, 1978)。無疑, S0、S4兩實驗組間血清生化指標(biāo)中僅TG含量存在顯著差異, 呈S0>S4(<0.05) (表1), 肝脂肪酶、胃蛋白酶和腸淀粉酶分別呈S0 GLU是生物有機體可直接利用的能源物質(zhì)(陳永祥等, 2009)。肝臟是糖原的主要存儲器官(美國科學(xué)院國家研究委員會, 2015)。李有志等(2022)研究指出, 保持血清中GLU含量相對穩(wěn)定對于維系非休眠期中華鱉的正常生命活動具極端重要性。無疑, S16實驗組GLU-O含量的顯著下降(<0.05)和肝臟SOD、POD、GSH-PX、Ca2+/Mg2+-ATPase酶活力的顯著高企(<0.05) (表1, 圖3)的結(jié)果, 既表明離水停食16 d后的實驗鱉已進入以肝臟細胞抗氧化和跨膜運輸壓力顯著高企為主要特征的低血糖生理紊亂期, 也揭示了低血糖是引發(fā)S16實驗組免疫力、抗疲勞能力和持續(xù)運動能力下降的重要誘因。 研究發(fā)現(xiàn), 硬度與受檢動物組織的含脂量呈負相關(guān)(Dunajski, 1980; Nielsen, 2005; 李文倩等, 2010; Lin, 2012; 王志錚等, 2013b), 系表征組織質(zhì)構(gòu)的重要評價指標(biāo)。因此, 受檢組織的硬度可作為評判其脂肪消耗程度的重要依據(jù)。裙邊雖為中華鱉的惰性組織(牟恩鏇等, 2021), 但并非儲存脂肪的主要蓄庫。無疑, 質(zhì)構(gòu)評價指標(biāo)中唯硬度與連續(xù)翻身次數(shù)均呈S4≈S8>S12≈S16的結(jié)果(圖1, 表3), 在進一步印證脂肪系實驗鱉離水停食期間流失的主要能源物質(zhì)的同時, 也反映了裙邊含脂量的顯著下降意味著實驗鱉機體能量已處于嚴重透支狀態(tài)的客觀事實。鑒于此,可將裙邊硬度作為指示中華鱉體能表露顯著下降的關(guān)鍵評價指標(biāo)。 (1) 中華鱉具快速翻身復(fù)原的行為學(xué)特性。本研究中實驗鱉連續(xù)翻身次數(shù)均主要集中于1 min內(nèi)的結(jié)果, 既印證了翻身作為其生存直接相關(guān)特性所表露出的強敏感性, 也映射出其為完成連續(xù)翻身所需償付的巨大能耗。故, 可在生產(chǎn)和銷售實踐中將連續(xù)翻身能力作為評價中華鱉活力或體質(zhì)強健程度的重要依據(jù)。 (2) 中華鱉的連續(xù)翻身能力既與所處的活動階段有關(guān), 也與離水停食時間過長導(dǎo)致的體能顯著下降有關(guān)。較高的心臟抗氧化壓力是尚處于活動復(fù)蘇期末期的S0實驗組個體翻身能力偏弱的重要原因, 免疫力和抗疲勞能力下降是引發(fā)S12和S16兩實驗組個體持續(xù)翻身能力顯著弱于S4和S8的根本原因。 (3) 在本研究實驗條件下, 離水停食0~4 d、4~8 d、8~12 d依次為實驗鱉啟動體脂大量消耗的應(yīng)激期、向低能耗代謝模式切換的體能穩(wěn)定期、以肝臟細胞跨膜運輸壓力高企為代價的體能下降期, 離水停食16 d后的實驗鱉已進入以肝臟細胞抗氧化和跨膜運輸壓力顯著高企為主要特征的低血糖生理紊亂期。 (4)裙邊雖為中華鱉的惰性組織, 但并非儲存脂肪的主要蓄庫。無疑。S12和S16實驗組個體持續(xù)翻身能力的顯著下降(<0.05)與裙邊脂質(zhì)的大幅流失相契合的結(jié)果, 表明裙邊硬度可作為指示中華鱉體能表露顯著下降的關(guān)鍵評價指標(biāo)。 王志錚, 付英杰, 楊磊, 等, 2013a. 三種養(yǎng)殖模式下日本鰻鱺()養(yǎng)成品體色和肌肉品質(zhì)的差異[J]. 海洋與湖沼, 44(4): 1042-1049. 王志錚, 趙晶, 楊磊, 等, 2013b. 三種養(yǎng)殖模式下日本鰻鱺()養(yǎng)成品血清生化指標(biāo)和臟器消化酶、抗氧化酶活力的差異[J]. 海洋與湖沼, 44(2): 403-408. 王志錚, 施建軍, 呂敢堂, 等, 2006. 受短期饑餓脅迫下麥瑞加拉鯪魚()幼魚的生長、肌體組分及其內(nèi)臟消化酶活力的變化特征[J]. 海洋與湖沼, 37(3): 218-224. 農(nóng)業(yè)農(nóng)村部漁業(yè)漁政管理局, 全國水產(chǎn)技術(shù)推廣總站, 中國水產(chǎn)學(xué)會, 2022. 2022中國漁業(yè)統(tǒng)計年鑒[M]. 北京: 中國農(nóng)業(yè)出版社: 24, 35. 牟恩鏇, 申屠琰, 熊玉鳳, 等, 2021. 池塘專養(yǎng)模式下中華鱉養(yǎng)成品形態(tài)性狀對體質(zhì)量和凈體質(zhì)量的影響效應(yīng)[J]. 浙江海洋大學(xué)學(xué)報(自然科學(xué)版), 40(1): 22-28, 79. 李文倩, 李小勤, 冷向軍, 等, 2010. 鱖魚肌肉品質(zhì)評價的初步研究[J]. 食品工業(yè)科技, 31(9): 114-117, 121. 李有志, 牟恩鏇, 申屠琰, 等, 2022. 越冬對池塘專養(yǎng)模式下中華鱉()機體形態(tài)、血清生化、臟器相關(guān)功能酶活力及肌肉和裙邊物性的影響[J]. 海洋與湖沼, 53(2): 421-429. 陳永祥, 肖玲遠, 嚴太明, 等, 2009. 野生和養(yǎng)殖裂腹魚血液學(xué)指標(biāo)的比較研究[J]. 水生生物學(xué)報, 33(5): 905-910. 陳師師, 2006. 中華鱉氣調(diào)保鮮技術(shù)研究[D]. 杭州: 浙江工商大學(xué). 國家技術(shù)監(jiān)督局, 1994. 食品標(biāo)簽通用標(biāo)準(zhǔn): GB 7718-1994[S]. 北京: 中國標(biāo)準(zhǔn)出版社. 徐本力, 1984. 第三講論運動負荷[J]. 福建體育科技(S1): 93-168. 謝小軍, 鄧利, 張波, 1998. 饑餓對魚類生理生態(tài)學(xué)影響的研究進展[J]. 水生生物學(xué)報, 22(2): 181-188. 美國科學(xué)院國家研究委員會, 2015. 魚類與甲殼類營養(yǎng)需要[M]. 麥康森, 李鵬, 趙建民, 譯. 北京: 科學(xué)出版社: 167. ARNOLD S J, 1983. Morphology, performance and fitness [J]. American Zoologist, 23(2): 347-361. DELMAS V, BAUDRY E, GIRONDOT M,, 2007. The righting response as a fitness index in freshwater turtles [J]. Biological Journal of the Linnean Society, 91(1): 99-109. DUNAJSKI E, 1980. Texture of fish muscle [J]. Journal of Texture Studies, 10(4): 301-318. FINKLER M S, 1999. Influence of water availability during incubation on hatchling size, body composition, desiccation tolerance, and terrestrial locomotor performance in the snapping turtle[J]. Physiological and Biochemical Zoology, 72(6): 714-722. KUTTY M N, 1978. Ammonia quotient in Sockeye salmon () [J]. Journal of the Fisheries Research Board of Canada, 35(7): 1003-1005. LIN W L, ZENG Q X, ZHU Z W,, 2012. Relation between protein characteristics and TPA texture characteristics of crisp grass carp (C. ET V) and grass carp () [J]. Journal of Texture Studies, 43(1): 1-11. NIELSEN D, HYLDIG G, NIELSEN J,, 2005. Liquid holding capacity and instrumental and sensory texture properties of herring (L.) related to biological and chemical parameters [J]. Journal of Texture Studies, 36(2): 119-138. STIRLING H P, 1976. Effects of experimental feeding and starvation on the proximate composition of the European bass[J]. Marine Biology, 34(1): 85-91. EFFECTS OF AIR EXPOSURE AND STARVATION STRESS ON THE FLIPPING ABILITY, SERUM BIOCHEMISTRY, SOFT TISSUE SKIRT QUALITY, AND ORGAN-RELATED ENZYME ACTIVITY IN CHINESE SOFT-SHELLED TURTLES ZHENG Jia-Hao1, SHENTU Yan1, 2, XU Ying-Jie1, LI You-Zhi1, ZHU Wei-Dong2, WANG Zhi-Zheng1 (1. College of Fisheries, Zhejiang Ocean University, Zhoushan 316022, China; 2. Yuyao Aquaculture Technology Extension Center, Yuyao 315400, China) Chinese soft-shelled turtles have long been consumed as live products, and air exposure and starvation are inevitable environmental stresses during bulk sales. Therefore, exploring the impact of air exposure and starvation on the commercial value of Chinese soft-shelled turtles and its mechanisms is of great practical significance for determining the shelf life of live sales. In this study, male commercial turtles [average mass (546.86±94.70) g] subjected to air exposure and starvation treatment for 0 d (S0group), 4 d (S4group), 8 d (S8group), 12 d (S12group), and 16 d (S16group) after being caught from the same pool on May 20, 2020, were used as research subjects. The effects of air exposure and starvation on their flipping ability, serum biochemistry, soft tissue skirt quality, and organ enzyme activity were systematically studied under normal temperature conditions. The results show that: (1) The consecutive flipping frequency of experimental turtles was S4≈S8>S0≈S12≈S16, mainly concentrated within 1 minute; (2) There were no significant differences in serum TP, ALB, TC, and LDL-C between groups (>0.05). GLU-O showed S0≈S4≈S8≈S12>S16, TG, and HDL-C, except that TG content in S4and S8was lower than that in S0(<0.05), and HDL-C content in S12was higher than that in S0(<0.05), there were no significant differences between other experimental groups (>0.05); (3) Among the seven measured soft tissue skirt quality indicators, only hardness completely coincided with the order of consecutive flipping frequency, which was S4≈S8>S12≈S16; (4) Hepatic lipase and gastric protease activities were S4>S0≈S8≈S12≈S16and S0≈S16>S4≈S8≈S12, respectively. Except for the significantly higher intestinal amylase activity in the S16group than in the S0and S4groups (<0.05), there were no significant differences between other groups (>0.05); (5) Hepatic ATPase and antioxidant enzyme activities showed a stepwise upward trend with the extension of air exposure and starvation time. Cardiac antioxidant enzyme activities showed a downward trend from 0 d to 4 d, stabilization from 4 d to 12 d, and an upward trend from 12 d to 16 d. Pulmonary antioxidant enzyme activities showed an upward trend from 0 d to 4 d, a downward trend from 4 d to 8 d, stabilization from 8 d to 12 d, and a subsequent upward trend from 12 d to 16 d. Among S4, S8, and S12, only splenic GSH-PX enzyme activity showed a clear misalignment in the order of enzyme activities, with S4 Chinese soft-shelled turtle; air exposure and starvation; flipping; soft tissue skirt quality; serum biochemistry; digestive enzyme; ATPase; antioxidant enzyme * 國家星火計劃項目, 2012GA701053號; 中央財政重大農(nóng)業(yè)技術(shù)推廣項目, YY2017003號; 寧波市科技局農(nóng)業(yè)創(chuàng)新創(chuàng)業(yè)項目, 2014C91026號。鄭嘉豪, 碩士研究生, E-mail: 1473829907@qq.com; 同等貢獻第一作者: 申屠琰, 高級工程師, 碩士生導(dǎo)師, E-mail: 122752557@qq.com 王志錚, 教授, E-mail: wzz_1225@163.com 2023-05-18, 2023-09-05 S966.5; Q956 10.11693/hyhz202305001071 材料與方法
1.1 供試中華鱉
1.2 實驗方法
1.3 數(shù)據(jù)處理
2 結(jié)果
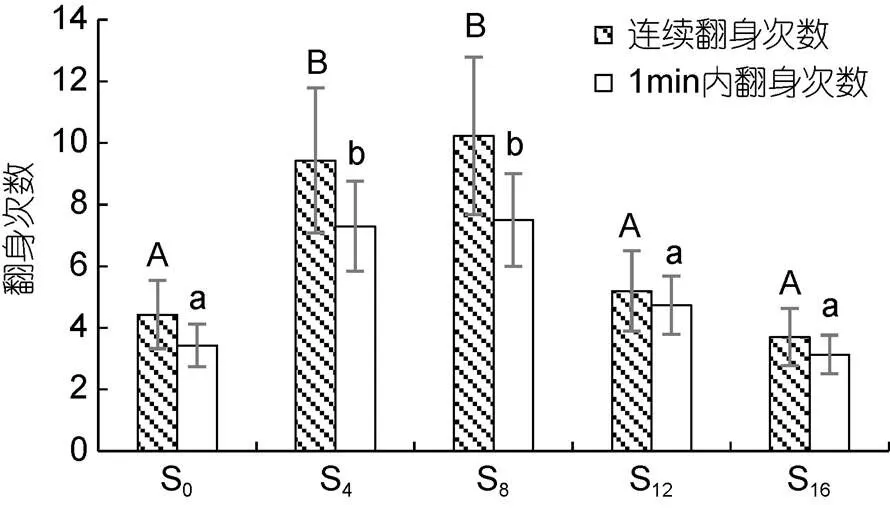
2.1 離水停食對翻身能力的影響
2.2 離水停食對血清生化指標(biāo)的影響
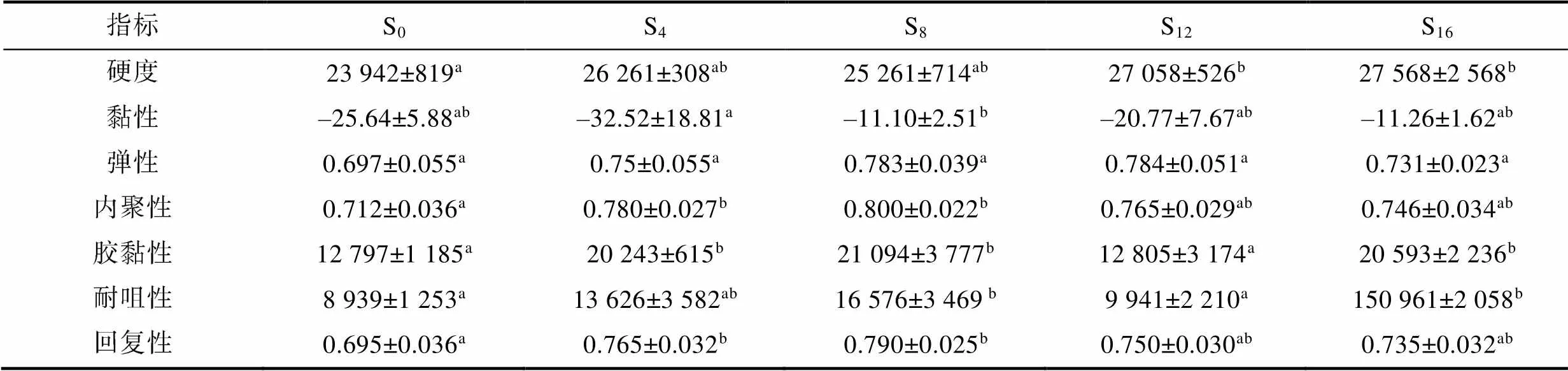
2.3 離水停食對裙邊質(zhì)構(gòu)的影響
2.4 離水停食對消化酶活力的影響

2.5 離水停食對肝臟ATP酶和抗氧化酶活力的影響
2.6 離水停食對心臟抗氧化酶活力的影響
2.7 離水停食對脾臟抗氧化酶活力的影響
2.8 離水停食對肺抗氧化酶活力的影響
3 討論
3.1 離水停食對實驗鱉翻身能力的影響特征

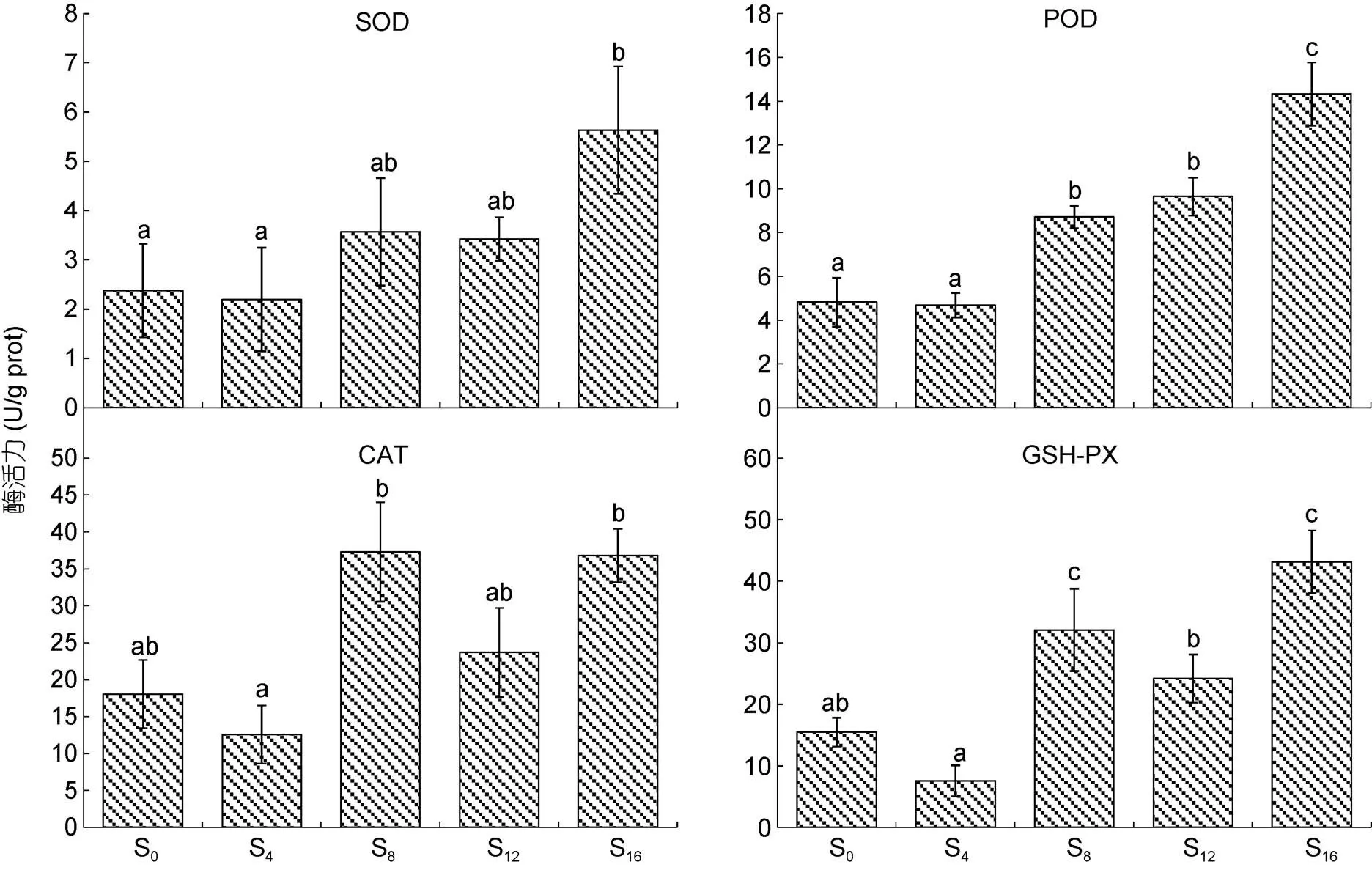
3.2 離水停食對實驗鱉臟器抗氧化生理的影響特征


3.3 離水停食對實驗鱉能源物質(zhì)利用對策和裙邊質(zhì)構(gòu)的影響特征
4 結(jié)論
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42音樂探索(2022年2期)2022-05-30 21:01:37中老年保健(2021年3期)2021-08-22 06:50:04天津醫(yī)科大學(xué)學(xué)報(2021年2期)2021-03-29 05:31:08現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52太空探索(2016年5期)2016-07-12 15:17:55
